
Abstract
The WNT/β-catenin signaling pathway plays a central role in the biology of the periodontium, yet the function of specific extracellular WNT ligands remains poorly understood. By using a Wnt1-inducible transgenic mouse model targeting Col1a1-expressing alveolar osteoblasts, odontoblasts, and cementoblasts, we demonstrate that the WNT ligand WNT1 is a strong promoter of cementum and alveolar bone formation in vivo. We induced Wnt1 expression for 1, 3, or 9 wk in Wnt1Tg mice and analyzed them at the age of 6 wk and 12 wk. Micro–computed tomography (CT) analyses of the mandibles revealed a 1.8-fold increased bone volume after 1 and 3 wk of Wnt1 expression and a 3-fold increased bone volume after 9 wk of Wnt1 expression compared to controls. In addition, the alveolar ridges were higher in Wnt1Tg mice as compared to controls. Nondecalcified histology demonstrated increased acellular cementum thickness and cellular cementum volume after 3 and 9 wk of Wnt1 expression. However, 9 wk of Wnt1 expression was also associated with periodontal breakdown and ectopic mineralization of the pulp. The composition of this ectopic matrix was comparable to those of cellular cementum as demonstrated by quantitative backscattered electron imaging and immunohistochemistry for noncollagenous proteins. Our analyses of 52-wk-old mice after 9 wk of Wnt1 expression revealed that Wnt1 expression affects mandibular bone and growing incisors but not molar teeth, indicating that Wnt1 influences only growing tissues. To further investigate the effect of Wnt1 on cementoblasts, we stably transfected the cementoblast cell line (OCCM-30) with a vector expressing Wnt1-HA and performed proliferation as well as differentiation experiments. These experiments demonstrated that Wnt1 promotes proliferation but not differentiation of cementoblasts. Taken together, our findings identify, for the first time, Wnt1 as a critical regulator of alveolar bone and cementum formation, as well as provide important insights for harnessing the WNT signal pathway in regenerative dentistry.
Keywords
Introduction
The WNT β-catenin signaling pathway is central for differentiation and proliferation of dental stem cells. The canonical WNT signaling is mediated via the binding of a WNT ligand to the 7-pass transmembrane receptor Frizzled (FZD) and the single-pass low-density lipoprotein receptor–related protein 5 or 6 (LRP5/6), leading to dephosphorylation-mediated stabilization and nuclear translocation of β-catenin (MacDonald and He 2012). Transgenic mouse studies have demonstrated that tissue-specific deletion or overexpression of β-catenin affects all stages of dental development, including root formation (Jarvinen et al. 2006; Liu et al. 2008; Chen et al. 2009; Kim et al. 2013; Zhang et al. 2013). However, until now, it has not been possible to target β-catenin pharmacologically (Nusse and Clevers 2017). A more promising strategy involves targeting extracellular components of the WNT β-catenin pathway. Here, an antibody targeting SOST, a negative regulator of WNT β-catenin, has only recently been approved for the treatment of women with severe postmenopausal osteoporosis (Cheng et al. 2020).
So far, mutations in WNT10A and WNT10B have been shown to cause syndromic tooth agenesis (in combination with malformation of other epithelial-derived organs such as hair and glands) and a nonsyndromic type of tooth agenesis in humans (Adaimy et al. 2007; Yu et al. 2016). In this study, we introduce the WNT ligand (WNT1) to be important for alveolar bone and cementum formation. Previously, we and others have shown that Wnt1 is a central regulator of bone formation (Laine et al. 2013; Gori et al. 2016; Joeng et al. 2017; Luther et al. 2018). Mutations in WNT1 are associated with osteogenesis imperfecta and with early onset osteoporosis (Fahiminiya et al. 2013; Keupp et al. 2013; Joeng et al. 2014), which is phenocopied in mice carrying the human WNT1 mutation or with a cell-specific deletion of Wnt1 in the osteoblast lineage. Furthermore, using a Wnt1-transgenic mouse line that expresses Wnt1 under the control of a Col1a1-responsive promoter, it has been shown that Wnt1 has a strong bone-anabolic function (Luther et al. 2018). Here, however, it remains to be determined whether Wnt1 has the same bone-anabolic function in the alveolar bone. Moreover, since Wnt1 is a key regulator of neural crest cells, which account for the majority of dental mesenchymal stem cells (Echelard et al. 1994), it is important to understand the role of Wnt1 during postnatal tooth formation. We, therefore, asked how Wnt1 affects postnatal dental development using Wnt1-transgenic mice with inducible expression of Wnt1 in Col1a1-expressing cells such as alveolar osteoblasts, cementoblasts, and odontoblasts.
Materials and Methods
Mice
To generate Wnt1Tg mice, we crossed Col1a1-rtTA mice (Peng et al. 2008) with ptet-Wnt1 mice (Gunther et al. 2003) on a mixed background. For genotyping, tail biopsies were used to identify double transgenic mice (Wnt1Tg) and control mice (mice lacking either Col1a1-rtTA or ptet-Wnt1). All other genotype litters were excluded from the study. We analyzed male mice at the age of 6 and 12 wk and female mice at the age of 52 wk. For each experiment including statistical analysis, a minimum of biological triplicates was used. Mice were allocated randomly to DOX-ON, DOX-OFF groups (coin flip), and experiments (micro–computed tomography [CT] and staining) were performed in a blinded fashion.
All mice were maintained in the animal facility of the University Medical Center Hamburg-Eppendorf in agreement with our animal ethics committee and comply with the ARRIVE Checklist (Percie du Sert et al. 2020) (see more details in the Appendix).
Micro-CT and Stereomicroscopy
Skulls were analyzed by contact radiography with a Faxitron X-ray cabinet (Faxitron X-ray Corp.), followed by micro-CT scanning (µCT-40; Scanco Medical) with a voxel size of 15 μm, 55 kV, 145 µA, and 200 ms as previously described (Koehne et al. 2013) and analyzed (see the Appendix). For morphological analysis, incisors and mandibular molars were imaged with a high-resolution inverted microscope (Olympus DSX500i) and a 10× objective lens (DSX-10×; Leica Microsystems).
Histology
Histology was performed on undecalcified specimens for histomorphometry and decalcified material for hematoxylin and eosin staining, as described in the Appendix.
Immunohistochemistry
Immunohistochemistry was performed on 5-µm paraffin sections, as described in the Appendix.
Fluorescence In Situ Hybridization (HCR 3.0)
Staining was performed, using the Molecular Instruments HCR v.3.0 protocol, for “generic sample on slide.” Fluorescence in situ hybridization probes were designed and purchased from Molecular Instruments.
Quantitative Backscattered Electron Imaging
The degree of tissue mineralization was measured by means of the backscattered electron intensity, as previously described (Koehne et al. 2013) (see the Appendix).
Cell Culture
The cementoblast cell line (OCCM-30; D’Errico et al. 2000) was cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (v/v; HyClone; Logan). Proliferation and differentiation assays were performed as described in the Appendix.
Results
Wnt1 Overexpression Affects Mandibular and Alveolar Bone Formation
To analyze short- and long-term effects of Wnt1 expression on periodontal homeostasis, we used an inducible Wnt1 transgenic mouse model (hereafter called Wnt1Tg), which targets alveolar osteoblasts and odontoblasts. In this model, doxycycline (DOX)–dependent Wnt1 transgene expression is governed by the Col1a1 promoter-driven tetracycline-controlled transcriptional activator (Col1a1-tTA), resulting in conditional Wnt1 expression in cells expressing Col1a1 (under the 2.3-kb fragment promoter) (Rossert et al. 1995; Peng et al. 2008) upon DOX withdrawal. We induced Wnt1 expression for 1, 3, or 9 wk in Wnt1Tg mice and analyzed them at the age of 6 or 12 wk (Fig. 1A). To confirm transgene induction, we first compared the expression of Wnt1 and Col1a1 messenger RNA (mRNA) in molars and incisors of 6-wk-old Wnt1Tg mice and controls 1 and 3 wk after DOX withdrawal using in situ hybridization (Fig. 1B and Appendix Fig. 1B, C). Whereas the expression of Wnt1 was not detectable in teeth sections of control mice (Fig. 1B and Appendix Fig. 1B), Wnt1 mRNA expression was increased upon DOX withdrawal in osteoblasts, osteocytes, odontoblasts, cementoblasts, and cementocytes of Wnt1Tg mice (Fig. 1B and Appendix Fig. 1C).
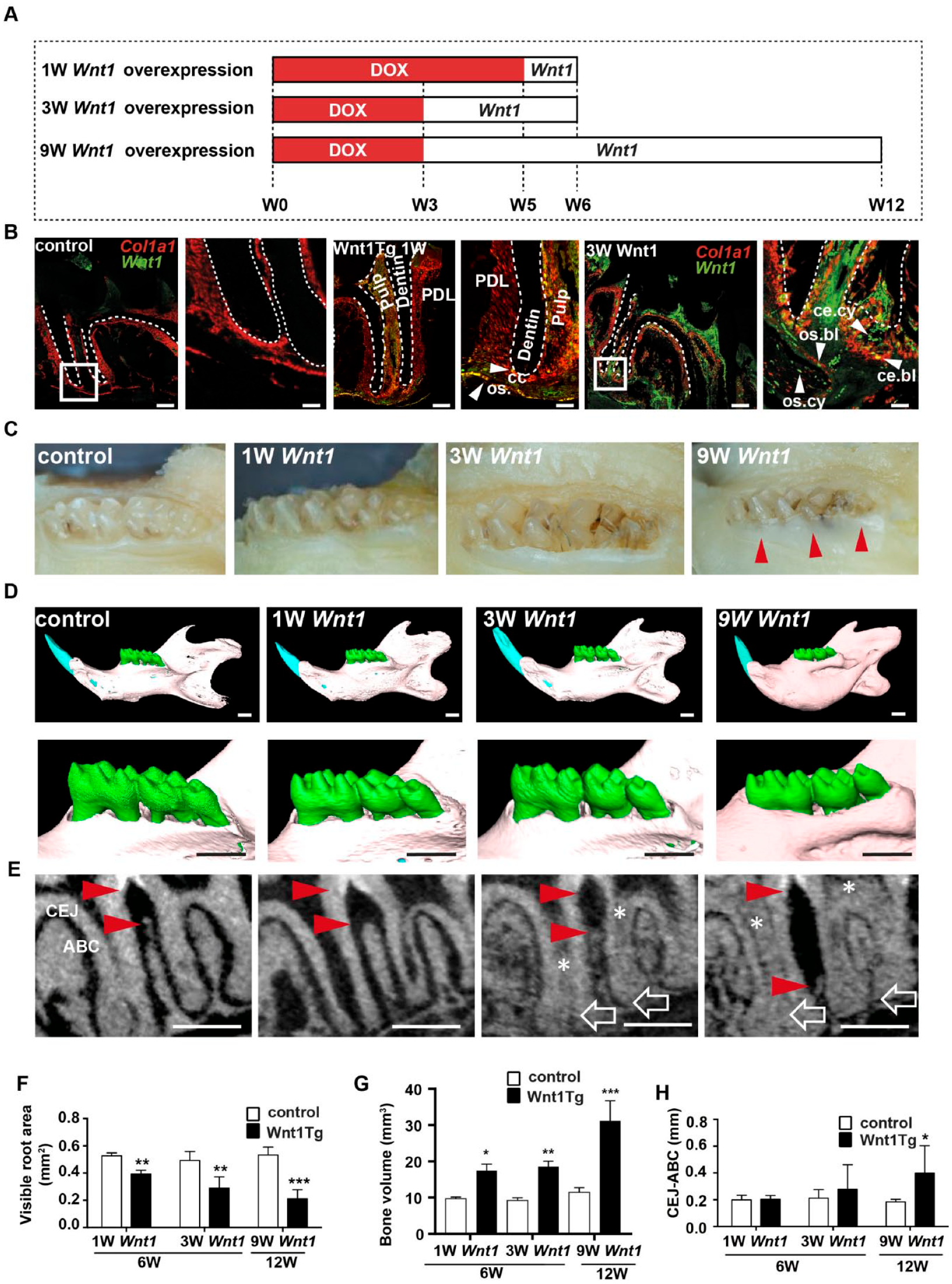
Conditional and tissue-specific expression of Wnt1 affects alveolar bone formation. (
Next, our initial macroscopic analysis of the teeth showed clear differences between Wnt1Tg and control mice (Fig. 1C and Appendix Fig. 1A). Here, the incisors of Wnt1Tg mice had a chalky white appearance after 3 and 9 wk of Wnt1 expression (Appendix Fig. 1A), whereas the crowns of the mandibular molars were normally formed. However, the visible crown height appeared shorter in Wnt1Tg mice after 9 wk of Wnt1 expression as compared to those of controls (Fig. 1C).
We next took advantage of micro-CT to analyze the mandibular molars (M1–M3) of Wnt1Tg mice, in which doxycycline has been removed for 1, 3, or 9 wk. The 3-dimensional (3D) segmentation images revealed a clear increase of mandibular bone mass in Wnt1Tg mice after 1, 3, and 9 wk of Wnt1 expression (Fig. 1D, upper panels). In addition, the alveolar ridges were higher in Wnt1Tg mice as compared to controls (Fig. 1D, lower panels). Quantification of the visible root area as well as mandibular bone volume revealed a significant bone growth already after 1 wk of Wnt1 induction, which is in line with previous findings showing that Wnt1 has a rapid bone-anabolic effect (Luther et al. 2018) (Fig. 1F, G). To our surprise, however, and contrary to these findings, we observed bone loss between the first and second molars in Wnt1Tg mice (Fig. 1E, red arrows). Although the variation of the observed alveolar bone loss was high, quantification of the distance between the cementum-enamel junction and the alveolar bone crest revealed significant differences between Wnt1Tg and control mice after 9 wk of Wnt1 expression (Fig. 1H).
Wnt1 Overexpression Leads to Ectopic Calcification in the Pulp
We next investigated the mandibular teeth in more detail using micro-CT imaging as well as undecalcified histology (Fig. 2). We observed that tooth length was not different between Wnt1Tg mice and wild-type (WT) mice at all ages analyzed (Appendix Fig. 2A, B). However, wall thickness analysis of the pulp together with von Kossa/van Gieson staining of nondecalcified sections revealed a decrease in pulp volume and the presence of mineralized matrix within the pulp cavities of Wnt1Tg mice after 3 and 9 wk of Wnt1 expression (Fig. 2A–D). Interestingly, our cellular analysis showed that the odontoblast layer was still present in incisors of Wnt1Tg mice after 1 wk of Wnt1 expression (Appendix Fig. 3A, B). Interestingly, we observed in the pulp a significant increase in vascularization upon Wnt1 expression (Appendix Fig. 3C). Importantly, after 3 wk and 9 wk of Wnt1 expression, the incisors were filled with ectopic calcifying matrix. A high proportion of this matrix consisted of nonmineralized organic tissue (45% after 3 wk of Wnt1 expression and 40% after 9 wk of Wnt1 expression) (Appendix Fig. 3D).
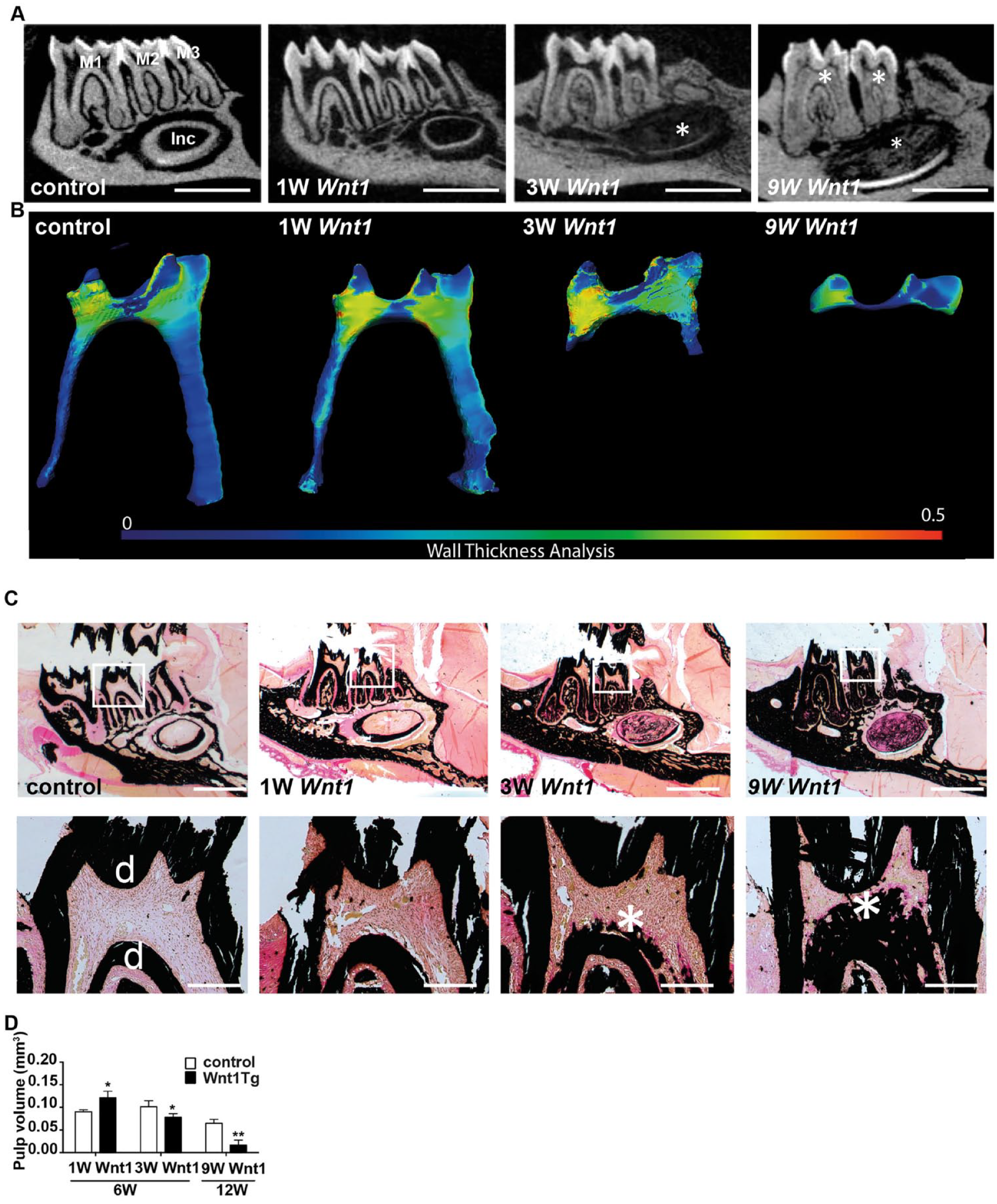
Wnt1 causes ectopic calcification in the pulp. (
To further analyze the observed increase in calcified tissue in the pulp and the effect on dentin formation, we performed fluorescent imaging of calcein dye, which had been injected 10 and 3 d before sacrifice (Appendix Fig. 4). The calcein double layers indicated that dentin formation was still evident after 1 and 3 wk of Wnt1 expression (Appendix Fig. 4B). This observation suggests that Wnt1 expression in odontoblasts does not directly affect its function. In addition, the pattern of calcein labels in the pulp indicates that the ectopic formation of the calcified tissue begins at the root apices and moves coronally toward the pulp horns.
Wnt1 Overexpression Increases Cementum Formation
We next studied the root cementum in Wnt1Tg mice (Fig. 3). Micro-CT analyses together with histological examinations showed that 3 and 9 wk of Wnt1 expression resulted in an increase of acellular and cellular cementum in Wnt1Tg mice (Fig. 3A, B). The acellular and cellular cementum layers were thicker after 3 and 9 wk of Wnt1 expression as compared to that of mice without Wnt1 expression (Fig. 3B). In fact, quantification revealed a 2-fold increase of acellular cementum thickness and cellular cementum area in Wnt1Tg mice after 3 and 9 wk of Wnt1 expression as compared to control mice (Fig. 3C, D). In addition, the amount of nonmineralized cellular cementum (i.e., cementoid) was significantly higher in Wnt1Tg mice as compared to those of control mice (Appendix Fig. 5A, B). Interestingly, this increase of nonmineralized matrix was not evident in bone (Appendix Fig. 5C). Quantification of fluorescent calcein imaging further demonstrated high cementum formation after 3 wk of Wnt1 expression, whereas no double calcein labels could be detected after 9 wk of Wnt1 expression (Appendix Fig. 4C, D). However, an important histological finding was also the severe periodontal breakdown between the molar roots after 9 wk of Wnt1 expression (Fig. 3B), confirming our micro-CT observations. In fact, von Kossa/van Gieson staining and polarized light imaging revealed detachment of PDL fibers in Wnt1Tg mice after 9 wk of Wnt1 expression (Appendix Fig. 6). Taken together, these results suggest that Wnt1 is a strong promoter of cementum formation. However, long-term exposure of Wnt1 may induce periodontal destruction and ectopic calcification.
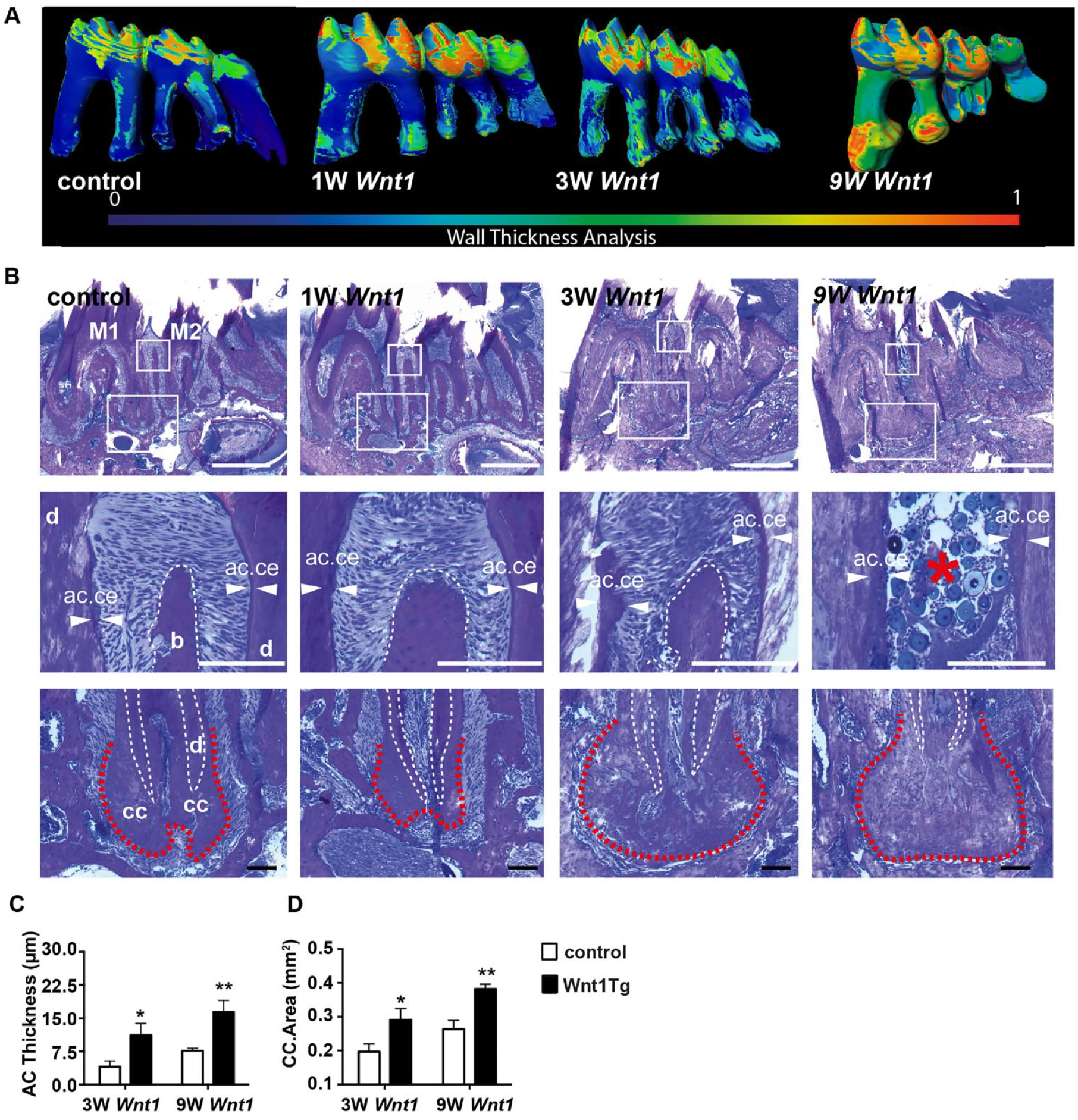
Wnt1 increases acellular and cellular cementum formation. (
The Ectopic Mineralization in the Pulp Has Cementum-Like Characteristics
To further characterize the ectopic calcification, we used immunostaining for noncollagenous matrix proteins of Wnt1Tg and control mice (Fig. 4). Osteopontin (OPN) was expressed in bone and acellular cementum of Wnt1Tg and WT mice (Fig. 4A). The expression of OPN in these tissues was clearly higher in Wnt1Tg as compared to those of WT mice. Cellular cementum was only slightly stained for OPN, whereas no expression of OPN could be detected in the dentin layers. To further distinguish dentin from cementum, we next stained for the dentin-matrix protein dentin sialophosphoprotein (DSP). DSP was detected in the dentin layers of Wnt1Tg and WT mice (Fig. 4B). The calcified pulp tissue of Wnt1Tg mice showed only a slight expression of DSP. Almost no expression of DSP could be detected in bone and cementum in both mouse lines.
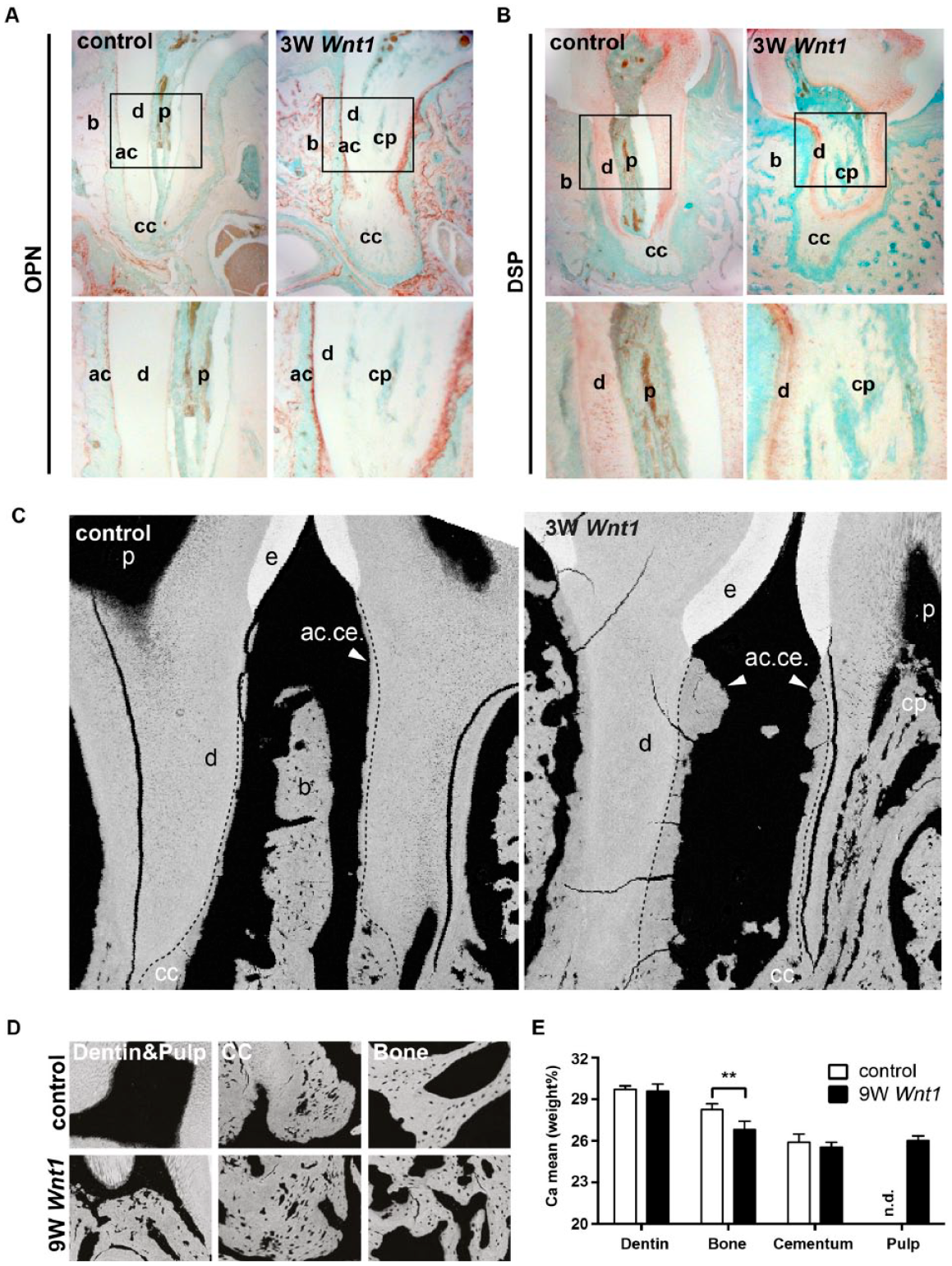
Characterization of ectopic pulp matrix in Wnt1Tg mice (
We next analyzed the calcium content using quantitative backscattered electron imaging and compared it with those of bone, dentin, and cementum (Fig. 4C–E). The backscattered images confirmed the periodontal bone loss and the increased cementum formation in Wnt1Tg (Fig. 4C). Quantification of the mean calcium content of dentin and cementum revealed no differences between Wnt1Tg and control mice (Fig. 4D, E). The mean calcium content of the alveolar bone was significantly lower in Wnt1Tg as compared to that of WT mice. This finding, however, is not unexpected given the fact that high bone formation can lower the mean calcium content as newly formed bone packages are less mineralized (Koehne et al. 2014). Importantly, the mean calcium content of the mineralized pulp matrix in Wnt1Tg was comparable to that of cellular cementum (Fig. 4E). These results altogether suggest that the calcium content and the distribution of noncollagenous proteins within the ectopic matrix share similarities with cellular cementum.
Wnt1 Overexpression Affects Cementoblast Proliferation Only in Growing Teeth
We finally asked how Wnt1 affects the periodontium of aged mice. To investigate this, we analyzed 52-wk-old mice after 9 wk of Wnt1 expression (Fig. 5A). This experimental setup allowed us to study both developing teeth (incisor) as well as tooth maintenance (molar). Micro-CT imaging and decalcified histology revealed that the molars of aged Wnt1Tg mice were less affected by Wnt1 induction (Fig. 5B). In fact, there was no evidence of pulp calcification or cementum increase. Furthermore, the periodontal bone between the first and second molars was not reduced in aged Wnt1Tg mice (Fig. 5C). The alveolar ridges, however, were higher in aged Wnt1Tg mice as compared to those of WT mice. Although this bone-anabolic effect was less clear as compared to those of younger Wnt1Tg mice, quantification of the visible root area revealed a significant reduction in Wnt1Tg mice as compared to WT (Fig. 5D). We also observed significantly increased mandibular bone volume in Wnt1Tg mice as compared to WT (Fig. 5E). This indicates that Wnt1 promotes alveolar bone formation in aged mice without causing periodontal destruction as observed in younger mice. That Wnt1 expression primarily affects growing teeth was further documented by the strong effect on the incisor, where the pulp appeared fully calcified (Fig. 5B).
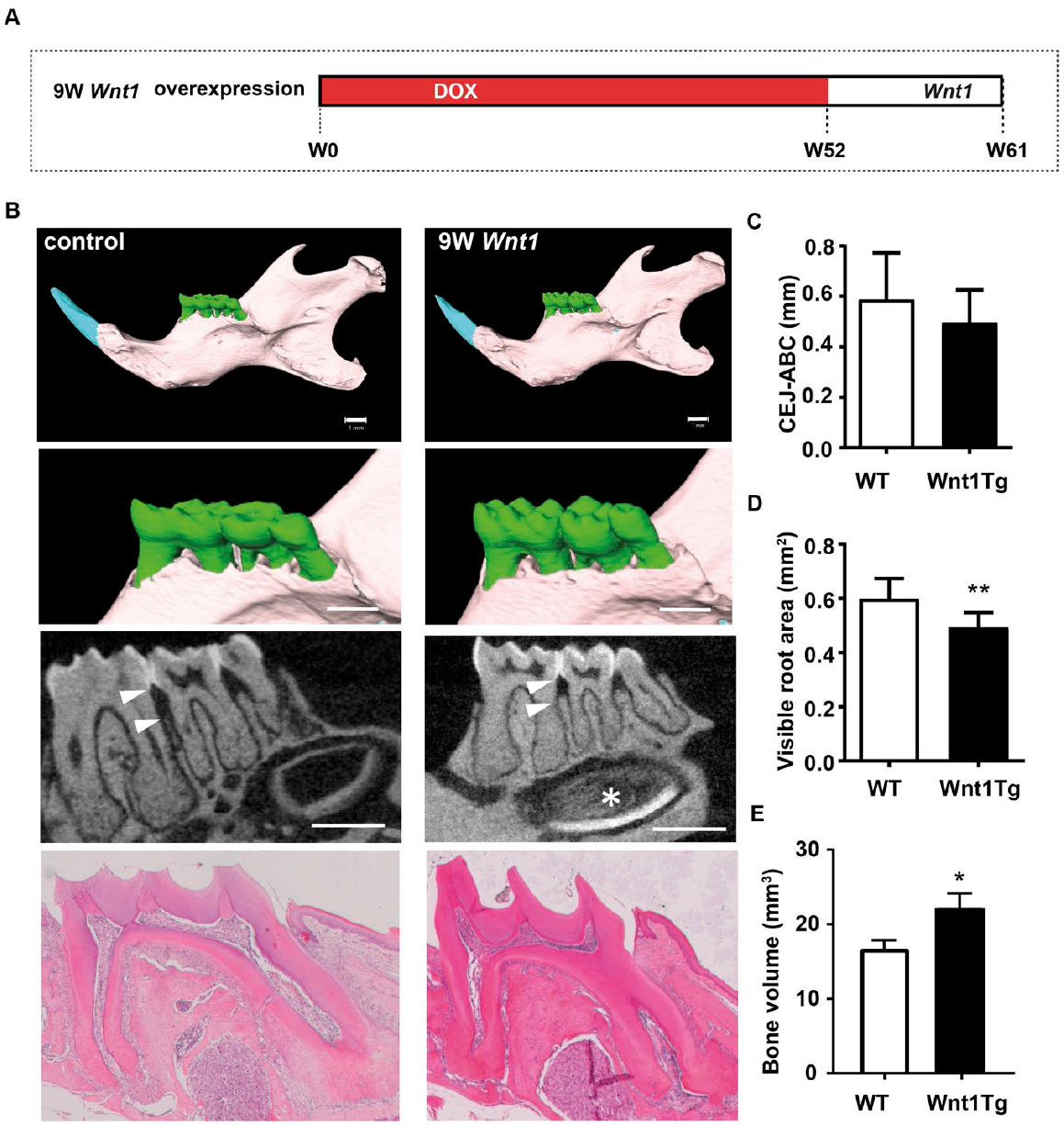
Wnt1 expression affects only growing teeth. (
Given these results, we finally determined the effect of Wnt1 on cell proliferation. For this, we used the cementoblast cell line OCCM-30 stable expressing Wnt1-HA (Appendix Fig. 7A, B). Here we observed that Wnt1 stimulates proliferation (Appendix Fig. 7C) but not differentiation of OCCM-30 cells (Appendix Fig. 7D, E).
Taken together, our results demonstrate that Wnt1 promotes cementum and alveolar bone growth. However, the exact molecular mechanisms of Wnt1 on cementoblasts remain subject to further research.
Discussion
Even though the role of WNT signaling in the regulation of tooth growth and maintenance is widely accepted (Tamura and Nemoto 2016), none of the 19 WNT ligands (except for WNT10A and WNT10B, which appeared to cause tooth agenesis) (Yu et al. 2016) have been identified to regulate tooth formation in vivo.
Here we present, for the first time in vivo, a novel role of Wnt1 for cementum formation and periodontal homeostasis. More specifically, our histological analyses together with micro-CT imaging revealed that Wnt1 is a strong promoter of alveolar bone and cementum formation. This is in line with a previous study showing that constitutive activation of β-catenin in the dental mesenchyme leads to differentiation of cementoblasts and induces excessive cementum formation in mice (Kim et al. 2011). However, by using an inducible and tissue-specific activation of WNT signaling, we provide several new insights in the role of WNT signaling for periodontal homeostasis. For example, one peculiar dental finding of this study was that long-term induction of Wnt1 resulted in ectopic formation of calcified tissue within the pulp chambers. Backscattered imaging of the calcium content and immunostaining for noncollagenous protein revealed that the composition of this ectopic matrix features characteristics very similar to cellular cementum. This is in line with our in situ stainings showing expression of Wnt1 in cementoblasts and cementocytes of cellular cementum. Our in vitro experiments using the OCCM-30 cementoblast cell lines further suggest that Wnt1 affects cementoblasts in a cell-autonomous manner. However, it is important to mention that our in vitro experiments do not entirely explain our findings in Wnt1tg mice, as we could not detect in OCCM-30 cells an increase in mineralization upon Wnt1 overexpression. The precise mechanism of how Wnt1 affects cementum formation, therefore, requires further investigation. Furthermore, we did not detect Wnt1 expression in acellular cementum cementoblasts despite a clear increase of acellular cementum formation (Appendix Fig. 8). It, therefore, remains to be determined whether the pOBCol2.3 promoter used in this study to regulate Wnt1 (Braut et al. 2002; Boban et al. 2006) is also expressed in acellular cementoblasts.
Furthermore, we show that Wnt1 not only serves as a stimulant for cementoblasts but also regulates alveolar and mandibular bone formation. This confirms our previously published data on the bone-anabolic effect of Wnt1 (Luther et al. 2018). Indeed, alveolar bone growth was evident in young as well as aged Wnt1tg mice. However, an unexpected finding in this regard was that long-term expression of Wnt1 in young mice resulted in periodontal bone loss. We interpret this as a sign of overstimulation as cementum formation needs to be orchestrated with the formation of periodontal fibers to achieve proper periodontal anchorage. It, therefore, seems likely that long-term Wnt1 expression causes an increase of dysfunctional cementum, in which PDL fibers cannot properly anchor anymore (Kim et al. 2011). This is further supported by our analyses of PDL organization in Wnt1tg mice. However, it is important to note that this periodontal breakdown was not observed in aged mice with long-term Wnt1 induction. Although we did not investigate the expression pattern of Wnt1 in aged mice, our analysis suggests that the detrimental effects of Wnt1 might therefore be more relevant for growing teeth. In this regard, it might also be interesting to study vascularization processes during bone formation, which have not been analyzed in this mouse model so far, although WNT signaling is crucial for angiogenesis (Olsen et al. 2017). We observed increased vascularization in the dental pulp after 1 wk of Wnt1 induction. Therefore, it seems that Wnt1-expressing odontoblasts induce angiogenesis by attracting endothelial cells (Masckauchan et al. 2005). This, however, has not been addressed in this study and deserves further experimental analysis.
Altogether, our findings open up a new paradigm in dental regeneration as β-catenin activating drugs such as BC21 (Kahn 2014) can have a huge effect on tooth development when given at a critical step during the development. Furthermore, prevention and management of alveolar bone loss are dependent upon regular periodontal tissue maintenance. Here we show that short-term induction of Wnt1 can increase bone formation and therefore prevent alveolar bone loss while having a supporting effect on cementum formation. Nevertheless, long-term exposure will potentially lead to unwanted calcified pulp tissue as well as periodontal bone loss at least in growing teeth. This seems also important to consider when osteoanabolic medication harnessing the WNT pathway is given to young patients.
Taken together, our results provide new insights into the role of Wnt1 for tooth development that can be employed for future regenerative strategies in dentistry.
Author Contributions
C. Nottmeier, J. Petersen, T. Koehne, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; N. Liao, contributed to conception, design, data acquisition, analysis, and interpretation, critically revised the manuscript; A. Simon, M.G. Decker, M. Kaucka, contributed to data acquisition, analysis, and interpretation, critically revised the manuscript; J. Luther, M. Schweizer, contributed to data acquisition and analysis, critically revised the manuscript; T. Yorgan, contributed to data acquisition and interpretation, critically revised the manuscript; E. Bockamp, contributed to data acquisition, critically revised the manuscript; B. Kahl-Nieke, M. Amling, T. Schinke, contributed to data interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-pdf-1-jdr-10.1177_00220345211012386 – Supplemental material for Wnt1 Promotes Cementum and Alveolar Bone Growth in a Time-Dependent Manner
Supplemental material, sj-pdf-1-jdr-10.1177_00220345211012386 for Wnt1 Promotes Cementum and Alveolar Bone Growth in a Time-Dependent Manner by C. Nottmeier, N. Liao, A. Simon, M.G. Decker, J. Luther, M. Schweizer, T. Yorgan, M. Kaucka, E. Bockamp, B. Kahl-Nieke, M. Amling, T. Schinke, J. Petersen and T. Koehne in Journal of Dental Research
Footnotes
Acknowledgements
This work is in memory of Dr. Jean-Pierre David, a brilliant bone biologist, who inspired this and many other projects (DFG grant DA1067/5-1). The Wnt1-HA Plasmid was a find gift from Jan Kitajewski. We also thank UKE Microscopy Imaging Facility (DFG Research Infrastructure Portal: RI_00489) for its technical support. We further thank Dr. Maike Jaworski and Dr. Andreas Trumpp from the Deutsches Krebsforschungszentrum (DKFZ), Heidelberg, Germany, for providing the Col1a1-rtTA mouse line.
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: supported by the DFG (Deutsche Forschungsgemeinschaft) grants KO5420/1-1 and KO5420/1-2 (T. Koehne), DFG grant YO299/1-1 (T. Yorgan), DFG grant SCHI504/6-1 (T. Schinke), DFG grant AM103/29 (M. Amling), and a Swedish Research Council Vetenskapsrådet grant (J. Petersen).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.