
Abstract
Prosocial behaviours are more commonly found among the higher educated. We examined to what extent genetic effects on prosocial behaviour captured by polygenic scores for educational attainment and cognitive performance can be detected. We apply a combination of an extended family approach using sibling fixed effects with polygenic scores to participants and their siblings in the Wisconsin Longitudinal Study. These models better capture shared environmental influences on prosocial behaviour. We find that those with a higher genetic propensity to spend more years in education were more likely to have engaged in forms of formal prosocial behaviours, that is, blood donation, charitable giving and volunteer work. We found no clear association between the polygenic score for educational attainment and informal helping. The polygenic score for educational attainment explained ∼2% of the variance in formal prosocial behaviour. The association was substantially reduced when the highest level of education respondents attained was taken into account. Shared environmental influences among siblings did not affect the association of cognitive performance with formal prosocial behaviour. Shared environmental influences were responsible for about one-quarter of the association between educational attainment and formal prosocial behaviour. While the polygenic score for educational attainment could explain as little as ∼2% of the variance in formal prosocial behaviour, including it in the model helps us to better understand the pathways of inter-generational transmission and direction of causality in the association between educational attainment and prosocial behaviour.
Keywords
Introduction
The rapid development of genome-wide association studies (GWAS) in the past few decades has enabled new ways of identifying and disentangling genetic and environmental effects in a plethora of traits (Tam et al., 2019). In the field of socio-genomics at the intersection of genetics and sociology, educational attainment is one of the most studied variables (Mills and Tropf, 2020). Educational attainment is a key variable in the stratification process, affecting income, wealth and health around the globe (Breen and Jonsson, 2005; Hout, 2012). Therefore, scientists have invested considerable efforts in the identification of genetic variants associated with educational attainment. The most comprehensive GWAS to date (Okbay et al., 2022) relies on ∼3 million observations, explaining 12% to 16% of the variance in educational attainment.
Educational attainment is a ubiquitous correlate of an important category of prosocial behaviour – activities that benefit others at a cost to oneself (e.g. Benabou and Tirole, 2006; Penner et al., 2005). Prosocial behaviour is multi-dimensional (Padilla-Walker and Carlo, 2014): depending on whether an intermediary organisation is involved, prosocial behaviour can occur in formal or informal settings (Wittek and Bekkers, 2015). Formal prosocial behaviours involve intermediary organisations seeking contributions from individuals to recipients, and include charitable giving through fundraising organisations, volunteer work for non-profit organisations and the donation of substances of human origin, for example, blood and cells, through collection agencies. Informal prosocial behaviours do not involve intermediary organisations, but directly benefit recipients. They include social and emotional support to friends and family members, and caring for parents and children. Prosocial behaviours in formal settings, such as volunteer work and blood donation, are more likely to be planned than informal prosocial behaviours, which are more likely to occur spontaneously in ongoing social relationships.
Studies of prosocial behaviour among adults have shown that prosocial behaviour is positively related to empathic concern, particularly in informal settings (Einolf, 2008; Ottoni-Wilhelm and Bekkers, 2010). In formal settings, marked differences in prosocial behaviour are observed between citizens with lower and higher educational attainment, creating an educational gradient: higher-educated citizens are more involved in organisations as volunteers, donate higher sums of money to charities and usually are more likely to donate blood (Bekkers, 2006; Lee et al., 1999; Wilson and Musick, 1997). While the educational gradient in prosocial behaviour is not as prominent in informal settings (Taniguchi, 2012; Wilson and Musick, 1997), higher-educated citizens are also more likely to participate in politics and vote (Persson, 2015). Therefore, researchers speak about the ‘civic returns on education’ (Dee, 2004) and ‘nonpecuniary benefits of schooling’ (Oreopoulos and Salvanes, 2011). These expressions suggest that educational attainment enhances civic engagement, beyond the talents and benefits that individuals have obtained from their parents and family background.
In this study, we seek to provide a better explanation of the educational gradient in prosocial behaviour than previous research by taking into account genetic factors and family background characteristics that facilitate educational attainment. Previous studies have not investigated how family background and genetic factors are responsible for the educational gradient in prosocial behaviour. Due to the omission of genetic factors and family background characteristics, the associations of educational attainment with prosocial behaviour are likely to be overestimated. For instance, in their study of the education gradient in volunteering, Son and Wilson (2017) examine how perceived control beliefs may explain the association, assuming that control beliefs are gained through education. However, the possibility that educational attainment and perceived control may also result from genetic factors as well as family background characteristics is ignored. Without taking these factors into account, associations between educational attainment and outcomes such as volunteering are likely to be biased.
We investigate prosocial behaviour, in both formal and informal settings, among participants in the Wisconsin Longitudinal Study (WLS; Hauser et al., 2020). Specifically, we explore how shared family background effects – such as for example being exposed to the same parental role models – and genetic effects, captured by polygenic scores for years of education and cognitive performance, create differences in prosocial behaviour. We estimate the associations of prosocial behaviour with genetic variants known to be associated with educational attainment using an extended family approach (Bartels, 2021), by including data on siblings from the same family. With this design, we can identify the extent to which the educational gradient in prosocial behaviour is rooted in genetic variants and shared family background characteristics, such as shared parental role models, collaboration and traditions within the family.
The design of our study creates an informative test of theoretical models explaining civic returns of education. If the educational gradient in prosocial behaviour is maintained when genetic variants and shared family background characteristics are controlled, we can infer that the gradient does not originate from them, and further research on the contribution of educational attainment to outcomes such as prosocial behaviour is warranted. If, however, the educational gradient in prosocial behaviour is reduced or eliminated when genetic variants and shared family background characteristics are controlled, further research on the aetiology of educational attainment in terms of family background and genetic factors is required to explain the educational gradient in prosocial behaviour.
Theoretical framework and previous evidence
Causative and selection models of the education gradient in prosocial behaviour
How can higher levels of prosocial behaviour among the higher educated be explained? One possibility is offered by the causative model. This model explains the educational gradient in prosocial behaviour as the result of factors that are resulting from educational attainment (Persson, 2015). Education increases cognitive skills and provides citizens with information about societal needs and makes them aware of such needs (Wolfinger and Rosenstone, 1980). Furthermore, education equips citizens with civic skills, such as communication skills, knowledge of political or health systems and the ability to think critically, that enable them to contribute to collective goods and solve social issues (Brady et al., 1995). Such skills are valuable in social life and make higher-educated citizens more desirable as donors and volunteers. Educational attainment also helps citizens acquire higher status jobs, which are not only rewarded with social status but also with a higher income and ultimately wealth (Hout, 2012). Taken together, the cognitive, social and financial resources that citizens with higher educational attainment acquire enable them to engage more frequently and intensively in prosocial behaviour (Wilson and Musick, 1997).
The selection model explains the educational gradient in prosocial behaviour as the result of pre-adult socialisation characteristics that are present in individuals before they complete educational attainment (Persson, 2015). According to this model, individuals with a higher level of education are more engaged in prosocial behaviour because of common environmental as well as genetic factors that pre-dispose them to help others and contribute to the common good. A set of pre-adult socialisation characteristics associated with both educational attainment and prosocial behaviour may explain why higher-educated citizens are spending more resources helping others. Traits that pre-dispose individuals to attain a higher level of education such as cognitive performance also enable them to acquire and process information faster and more accurately, enable them to learn and train skills that require cognitive effort, achieve success on the labour market and earn higher incomes (Judge et al., 1999). These skills and resources are also positively associated with engagement in prosocial behaviour. In this selection model, the greater prosociality among the higher educated is yet another result of the ‘genetics of success’ (Belsky et al., 2016).
The selection model and the causative model can be true at the same time and their mechanisms can reinforce each other. Citizens may be more engaged in prosocial behaviour because of traits that are also associated with higher cognitive performance and educational attainment, while at the same time their cognitive abilities and educational attainment also facilitate prosocial behaviour.
Evidence from twin studies on pre-adult socialisation factors
Twin studies provide one type of evidence on pre-adult socialisation factors by decomposing variance in outcomes, such as educational attainment and prosocial behaviour, into three broad categories: genetic effects due to genetic make-up, identical for identical twins, shared environmental effects due to growing up in the same household with the same parents and unique environmental effects due to experiences from school, peers, health and other events, specific to one twin or the other. The genes and the shared environment of the family as well as the broader social context in which citizens grow up contain pre-adult socialisation factors that have long-lasting effects, even before individuals complete their education.
The evidence from twin studies demonstrates sizable genetic and shared environmental effects on both educational attainment as well as on prosocial behaviour. The proportions of variance in educational attainment and prosocial behaviours explained by shared environmental effects are relatively high compared to many other traits that behavioural geneticists have examined (Polderman et al., 2015). For educational attainment, genetic effects explain between 35% and 45% of the variance, and shared environmental effects explain between 25% and 40% of the variance (Polderman et al., 2015).
For prosocial behaviour, that is, volunteering and charitable giving, three studies drawing upon US data (Bekkers et al., 2017; Dawes et al., 2015; Son and Wilson, 2010) show that between 7% and 36% of the variance can be explained by genetic effects. Two studies (Dawes et al., 2015; Son and Wilson, 2010) report no evidence for shared environmental effects, and a third (Bekkers et al., 2017) reports that shared environmental effects explain between 0% and 26% of the variance in prosocial behaviour. For blood donation, a study drawing upon twin data from Denmark reported genetic effects explaining 53% of the variance in the propensity to become a blood donor, and shared environmental effects explaining 28% of the variance (Pedersen et al., 2015).
The need for genetically informative designs
Previous studies on prosocial behaviour have produced evidence that giving and volunteering behaviour of children and parents are correlated: children of parents who volunteer are more likely to volunteer (Bekkers, 2007; Binder, 2020; Mustillo et al., 2004). Parents who engage in informal helping behaviour are more likely to have children who do so themselves (Ramaekers et al., 2022). Children of parents engaging in charitable giving are more likely to engage in charitable giving themselves (Wilhelm et al., 2008). There is also evidence that blood donation is correlated between parents and children (Quéniart, 2013). Without a genetically informative design, however, such correlations do not tell us much about the aetiology of this similarity between parents and children (Bartels, 2021). Surveys and registry data containing responses from multiple siblings from the same family provide an opportunity to disentangle genetic, unique and shared environmental effects (Gidengil et al., 2019). Combining survey and registry data with increasingly available genetic data from large, genotyped datasets, offers an even more promising avenue for decomposing genetic effects, shared and unique environmental effects. Such an approach can help to tackle challenges of causal inference. While different methods to strengthen causal inference in observational research have been developed over the past few decades, genetically informed methods represent especially powerful tools to account for genetic and environmental confounding (Pingault et al., 2018). It should be acknowledged that effect sizes of all these factors are typically small and add only little to the overall explained variance in complex phenotypes such as education and prosocial behaviour (Bartels, 2021). Yet, not considering genetic confounding is no longer justified, given the large availability and accessibility of genetic data, not only in medicine but also related to social science, behavioural and economics questions.
Considering genetic confounding and including genetic data in more traditional social science research designs, not only helps us to better estimate the net effects of non-genetic factors, for example, environmental influences, and determine causal inferences, it also helps us identify and prioritise promising avenues for policy interventions (Pingault et al., 2018). For instance, if the associations of educational attainment with desirable outcomes merely or primarily reflect genetic effects on cognitive performance, it could be argued that these indicate genetic inequalities at birth. Public investments in education could then be targeted to support those who were unlucky in the DNA lottery (Harden, 2021).
Evidence on unique environmental effects of education
Evidence on genetic and shared environmental effects of education comes from co-twin control designs, also known as discordant twin designs (McGue et al., 2010). Because monozygotic twins who grow up in the same family are conceived with the same genes and are subject to the same shared environmental influences, any differences between them must be due to unique environmental effects. Moreover, if educational attainment affects prosocial behaviour through unique environmental effects, differences in the educational attainment of monozygotic twins should be correlated with differences in prosocial behaviour. The higher-educated twin of a pair should give more time, money and blood than their lower-educated twin.
Studies with co-twin control designs among monozygotic twins reveal that twins from the same pair with different levels of education are virtually identical in terms of their charitable giving (Bekkers et al., 2017) and volunteering (Bekkers et al., 2017; Gibson, 2001). The lack of differences between higher- and lower-educated monozygotic twins from the same pair indicates that educational attainment is not related to prosocial behaviour because of unique environmental effects. Instead, these results give rise to the expectation that a substantial proportion of the association between educational attainment and prosocial behaviour is due to genetic and shared environmental effects. To test these pathways, we present the first study of prosocial behaviour to include a polygenic score for educational attainment, along educational attainment itself and parental educational attainment.
Polygenic scores for educational attainment
One limitation of classical twin studies is that they provide only a broad estimate of the proportion of variance explained by genetic effects. This limitation is addressed through technological advancements and strong decline in the cost of genotyping and DNA sequencing, which have enabled direct measurements of specific genetic variants. By pooling sequenced DNA data from large samples of individuals who reported the number of years spent in education, or the highest level of education they attained, behavioural geneticists have been able to detect a large number of genetic variants that are associated with educational attainment. Based on these identified genetic variants, a polygenic score can be constructed. A polygenic score is a quantitative measure of the genetic propensity (‘risk’) for a certain phenotype (e.g. educational attainment, psychological traits or health and disease), that sums up the effects of genetic variants across the entire genome (Plomin and Von Stumm, 2022). Polygenic scores capture all robustly significant associations with genetic variants in one variable.
The fourth and most recent comprehensive GWAS of educational attainment (Okbay et al., 2022) draws upon genomic data of ∼3 million observations and identifies 3952 single-nucleotide polymorphisms (SNPs) associated with educational attainment. Polygenic scores based on the data explained 12% to 16% of the variance in educational attainment. These findings improve upon the third GWAS, based on 1,047,538 observations from the samples of respondents with European ancestry, which identified 1271 SNPs associated with educational attainment (Lee et al., 2018). This polygenic score we use here because the more comprehensive score is not yet available, explained 11% (a similar proportion) of the variance in educational attainment. This is about one-quarter to one-third of the variance in educational attainment that twin studies suggest can be explained by genetic effects. Subsequent studies have shown that the polygenic score captures SNPs that are also correlated with cognitive performance, occupational status, income, health indicators and subjective well-being (Belsky et al., 2016; Davies et al., 2016; Engzell and Tropf, 2019; Harden and Koellinger, 2020; Mills and Tropf, 2020), characteristics that are also related to prosocial behaviour (Penner et al., 2005; Wilson and Musick, 1997).
The association of educational attainment with prosocial behaviour may be due partly to the influence of cognitive performance on educational attainment and on prosocial behaviour. In this scenario, genetic effects on cognitive performance will be partly responsible for the association between educational attainment and prosocial behaviour in studies without a genetically informative design. To identify shared and unique environmental effects on educational attainment and prosocial behaviour, it is thus important to include statistical controls for cognitive performance.
Survey data from representative population samples indicate that the association between educational attainment and prosocial behaviours is weaker when controlled for cognitive performance. This holds for composite scores (Haider and Von Stumm, 2022), for volunteering (Proulx et al., 2018) and charitable giving (Bekkers, 2006; James, 2011). A twin study in Nigeria (Hur, 2020) reports that correlations between cognitive performance and prosocial behaviour are due to genetic effects. Finally, a study partially relying on the same survey data that we use documented a positive association between a polygenic score for educational attainment and voting in the USA and Sweden (Dawes et al., 2021). Educational attainment and cognitive performance accounted for about half of the relationship between the genetic propensity to acquire education and voter turnout. At the same time, parental educational attainment was still associated with voting in models including polygenic scores for educational attainment and cognitive performance.
The availability of polygenic scores capturing SNPs known to be associated with educational attainment and cognitive performance allows us to estimate more precisely how the educational gradient in prosocial behaviour evolves. Figure 1 depicts the conceptual model and hypothesised associations. By applying a combination of a sibling approach with polygenic scores, we aim to shed light on the complex interplay of genetic, shared and unique environmental effects in the association between educational attainment and prosocial behaviours. We hypothesised that both parental education as well as the polygenic scores for educational attainment and cognitive performance are associated with prosocial behaviour, and that educational attainment partially mediates these associations. Supported by previous research, we formulated the following hypotheses (H1–H4):
H1: The polygenic score for educational attainment (Educational-PS) is positively related to prosocial behaviour. H2: The association between the polygenic score for educational attainment and prosocial behaviour is partly mediated by the respondents’ educational attainment. H3: When controlling for the polygenic score for educational attainment and respondents’ educational attainment, an association between the parents’ educational attainment and prosocial behaviour remains. H4: The association between the polygenic score for educational attainment and prosocial behaviour occurs in both formal and informal prosocial behaviour. The aforementioned relationships are stronger for formal than for informal prosocial behaviour.
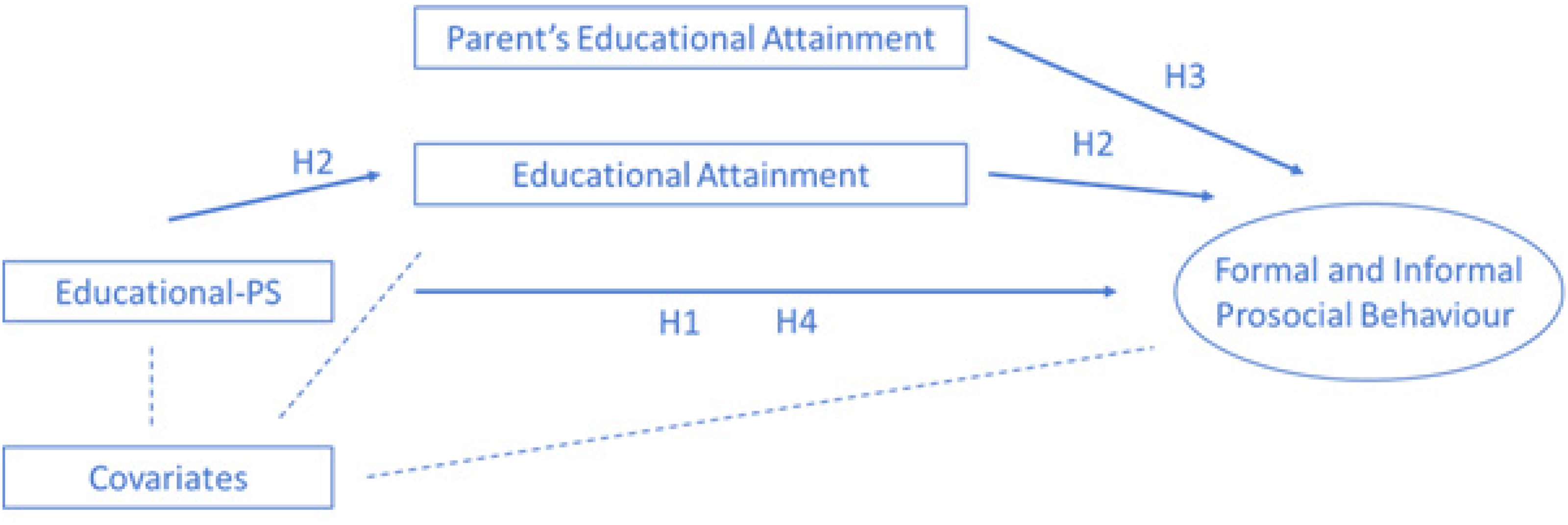
Conceptual model. An oval represents a latent variable; rectangles represent manifest indicators. Arrows indicate causal relationships; striped lines indicate potentially bidirectional associations.
Materials and methods
Data
We used data from the WLS, tracking a random sample of graduates from Wisconsin high schools in 1957 and their randomly selected siblings (Hauser et al., 2020). Follow-up surveys were conducted in 1964, 1975, 1992, 2004 and 2011, including questions on educational attainment, parental education, social and civic participation, health, home ownership and income. Between 2007 and 2011, DNA from 5967 participants was genotyped using the Illumina HumanOmniExpress-24-v1-1 BeadChip that includes 713,014 SNPs (Herd, 2016). Because polygenic scores based on the currently available GWAS have limited predictive value for persons with non-European ancestry (Duncan et al., 2019), we explored whether excluding participants who reported that the original nationality from their father's or mother's side was non-European (n = 93) or whose ancestry was unclear (n = 300) made a difference (see online Appendix Table S4 in the Supplemental materials). It did not, therefore we retained all observations. We used the data from Round 1 (1958) on IQ test performance and parental education, and from Round 5 (2004) on prosocial behaviour, as well as the polygenic scores for educational attainment and cognitive performance of n = 5015 graduates and n = 2713 siblings who lived in the same household in 1958. A pre-analysis plan for this study, the code producing the results and an Appendix with supplementary analyses are available at https://osf.io/db4wq.
Measures
Prosocial behaviour. We analysed prosocial behaviour in both formal and informal settings. Formal prosocial behaviour included lifetime engagement in blood donation, engagement in charitable giving in the past year and the number of types of organisations respondents volunteered for in the past year (0–5). The questions on volunteering referred to volunteering for religious organisations, educational organisations, political groups or labour unions, senior citizen and other national or local organisations. The question on charitable giving asked whether the respondent or spouse made charitable contributions of money or property totalling $500 or more coded as 1 = yes, 0 = no. The question on blood donation asked whether the respondent had ever given blood for the use of others coded as 1 = yes, 0 = no.
Informal prosocial behaviour included helping in four different areas: daily helping, housework helping, emotional/instrument support and childcare. For each of these areas, a sum score (0–6) was computed for the number of recipients to whom the participant had provided help during the last 12 months. Recipients included six different categories, including (1) friends, neighbours and co-workers, (2) parents, (3) children, (4) brothers and sisters, (5) grandchildren and (6) other relatives who do not live with the participant.
Educational attainment. Educational attainment is measured as the number of years of education that participants had completed at the time of Round 5 of the WLS (2004). To avoid cell counts <5 we collapsed respondents indicating less than 8 years in one category 8 years or less and respondents indicating 21+ years were included in the category 20 or more.
Polygenic scores for years of education and cognitive performance. The polygenic scores for years of education and cognitive performance that we use in this study were computed using summary statistics from the 2018 GWAS (Lee et al., 2018). The polygenic score for educational attainment (‘Educational-PS’) was calculated as a weighted sum of a WLS respondents’ genotypes at 1271 SNPs (Becker et al., 2021). The polygenic score for cognitive performance was based on 225 SNPs. In sensitivity analyses (see Tables S5 and S6 in the Supplemental materials) we report results of regression models including 10 principal components capturing population stratification (Herd and Sicinski, 2022) and multi-variate polygenic scores (Turley et al., 2018). The results of these models are similar to the results reported below.
Cognitive performance. The WLS includes measures for cognitive performance in the Henmon–Nelson Test of Mental Ability, a 30 min 90 item test which graduates completed in 1957 (Herd and Sicinski, 2022). Scores are provided by the Wisconsin Testing Service that administered the test (Retherford and Sewell, 1988).
Parental education. In the first round of the WLS, graduates reported the level of educational attainment of their father and mother. In the third round of the WLS, 2118 participants who had not previously reported on their parents’ education provided retrospective reports on the level of educational attainment of the head of the household when the respondent was 16–18 years old – we used the information available for 1837 fathers. For siblings who had missing values on parental education because they had not participated in the sibling survey, we assigned the parental educational attainment levels reported by graduates from the same families.
Family of origin characteristics. In addition to the level of education of the parents, we included measures of parental income, socio-economic status and town size. Data on parental income were obtained from tax records over the years 1957 to 1960. Family socio-economic status was a factor-weighted score produced by the WLS team based on father's education, mother's education, the socio-economic index score for father's occupation and average parental income. Town size was measured in nine categories, ranging from rural (father was a farmer), to over 150,000 (Milwaukee).
Covariates. In our analyses, we adjusted for a series of covariates that are likely to be correlated with educational attainment and prosocial behaviours: age, gender, subjective health, home ownership, household income, satisfaction with financial situation and parents’ educational attainment. Including these characteristics creates a conservative test, because they may also depend on educational attainment (see Table S8 in the Supplemental materials). The age of participants was measured in years in 2004; gender was coded as male = 0, female = 1; subjective health was measured in 2004 on a 5-point Likert scale, ranging from 1 = very poor to 5 = excellent; home ownership in 2004, coded as 1 = yes, 0 = no; household income was measured in 2004 as the sum of personal income per year by the respondent, the spouse (if any) and income from other household members measured in dollars; satisfaction with financial situation was measured in 2004 on a 5-point Likert scale, ranging from 1 = not at all to 5 = completely. Fathers’ and mothers’ educational attainment were measured in the years of education the parents of the participants had followed.
Analytic approach
First, we applied structural equation modelling (SEM) to generate latent variables for formal and informal prosocial behaviour (see Figure 1) using the following criteria for model fitness: comparative fit index (CFI) > 0.90, Tucker–Lewis index (TLI) > 0.90, the root-mean-square error of approximation (RMSEA) < 0.08 and Bayesian information criterion (BIC) (Barrett, 2007).
Next, we estimated bivariate correlations to examine whether the polygenic scores for educational attainment and cognitive performance were associated with educational attainment and prosocial behaviour in formal and informal settings. In a series of regressions, we estimated the association between educational attainment and prosocial behaviour with and without controlling for the polygenic scores for educational attainment and cognitive performance. In models without the polygenic scores, associations between prosocial behaviour and educational attainment are overestimated because genetic effects on educational attainment and prosocial behaviour are not accounted for. We partial out genetic effects by controlling for the polygenic scores capturing all known genetic variants associated with educational attainment and cognitive performance. We z-standardised all non-binary variables to facilitate comparisons of coefficients in the regressions. Table S10 in the Supplemental materials provides regressions with non-standardised variables.
In the regressions we estimated two types of models: one with and one without fixed effects for families. The models without fixed effects analyse variation in educational attainment and prosocial behaviours between respondents, without taking their genetic relatedness as siblings and similarity in family origins into account. These models do not take full advantage of the genetic information available in the data. In the models including fixed effects for families, differences between families are cancelled out, and only differences between respondents from the same family remain as a source of variance. This is a strong design for disentangling genetic and unique environmental effects. By definition, the family environment is a shared environmental factor. Because they are eliminated in the models including fixed effects for families, all differences we observe in these models must be due to genetic and unique environmental effects. Because we include polygenic scores of educational attainment and cognitive performance, a comparison of models that do and do not rule out shared environmental effects demonstrates to what extent correlates of prosocial behaviour are due to shared environmental effects.
Results
Descriptive statistics
Demographic information of the participants is presented in Table 1. Participants had a mean age of 64.2 years (SD: 4.13, range 40–86) in the 2004 survey. Of the sample, 52% were female. Participants had attained 13.9 years of education (SD 2.43, range 7–20) on average. Because the WLS consists of a sample of high school graduates, the minimum number of years of education for graduates is 12. Among siblings, 4% did not complete high school. Of the graduates, 55% and of siblings 47% did not continue in education after graduation. A bachelor's degree was obtained by 14% of graduates and 16% of siblings and 4% of graduates and 6% of siblings continued after a 2-year master's degree to obtain a professional degree, a PhD or MD.
Characteristics of study participants (n = 7728).
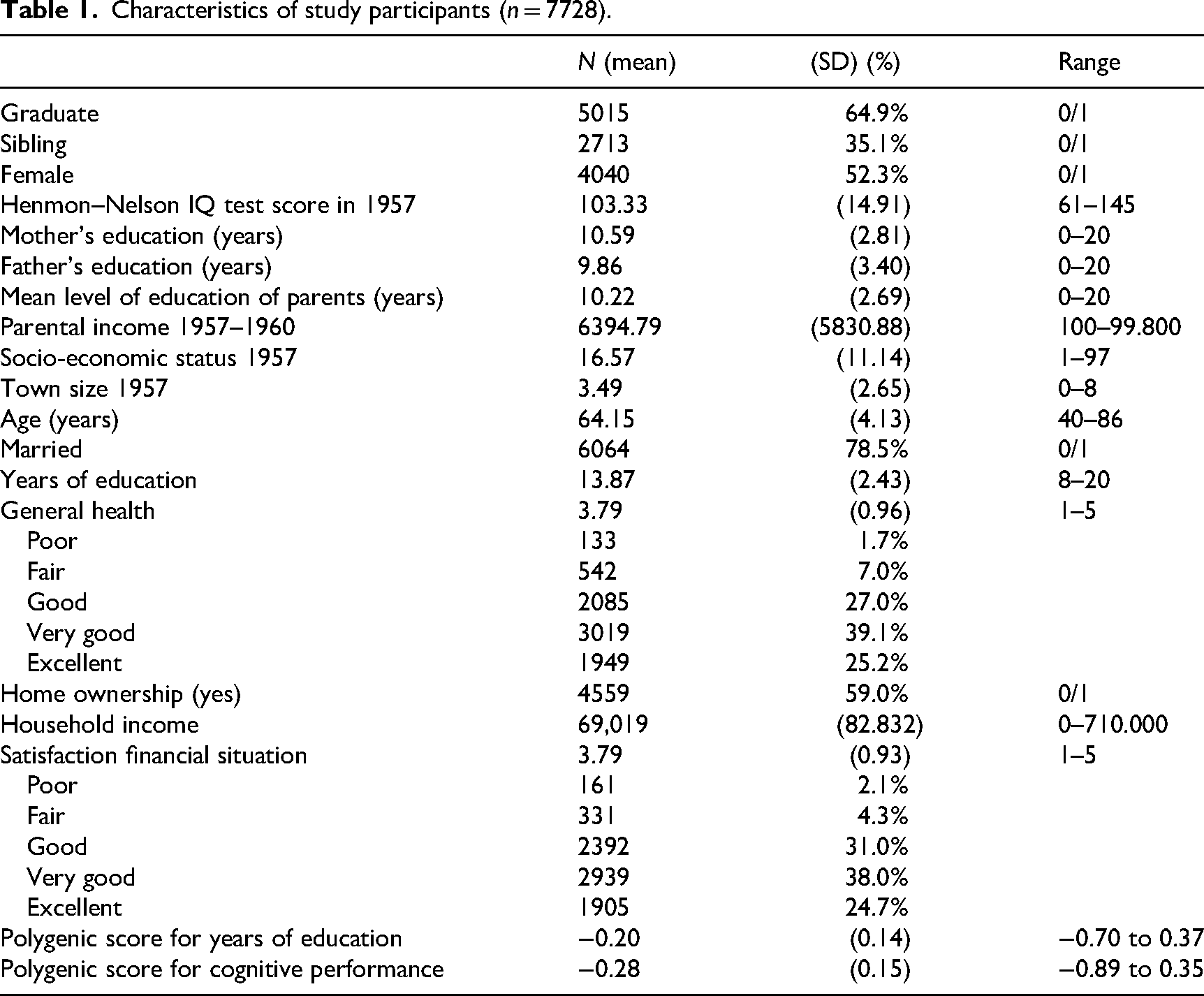
The average level of education of participants’ mothers was 10.6 years (SD: 2.81, range 0–20) and fathers had attained 9.9 years of education (SD: 3.40, range 0–20). The mode for mothers was a high school diploma after 12 years of school (37%). The mode for fathers was 8 years of school (29%). In total, 8% of mothers and 20% of fathers had completed fewer than 8 years of education. A bachelor's degree after 4 years of college had been attained by 6% of mothers and 8% of fathers.
Descriptives for formal and informal prosocial behaviours reported by participants are presented in Table 2. About half reported volunteering in the past year (49.1%), and ever having donated blood (53.3%). Of the participants, 64.7% reported charitable donations in the past year. Daily helping activities were reported by 63.7% of participants, 43.7% reported housework helping, 84.7% reported to have offered emotional/instrument support to friends, neighbours, co-workers, parents, children/grand-children or other relatives and 48.7% offered babysitting/childcare to others.
Description of prosocial behaviours of WLS cohort in the 2004 survey.

WLS: Wisconsin Longitudinal Study.
SEM measurement model
We estimated an SEM measurement model to generate latent variables of prosocial behaviour. The model with two factors, that is, formal and informal prosocial behaviour, reached good model fitness (CFI = 0.96, TLI = 0.93, RMSEA = 0.05 (90% confidence interval (CI) [0.04, 0.05]), BIC = 48,985.4). This two-factor model was superior to a one-factor model containing all indicators of prosocial behaviour (CFI = 0.85, TLI = 0.77, RMSEA = 0.09 (90% CI [0.08, 0.09]), BIC = 49,211.5). We retained the factor scores from the two-factor model and used them as dependent variables, that is, formal and informal behaviours in the analyses below.
Regressions
Hypothesis 1. Participants with higher polygenic scores for educational attainment were more engaged in formal prosocial behaviour, controlling for covariates. This result is in line with our hypothesis. The polygenic score for cognitive performance was not associated with formal prosocial behaviour. This result is evident from Table 3.
Regression analyses of formal prosocial behaviour
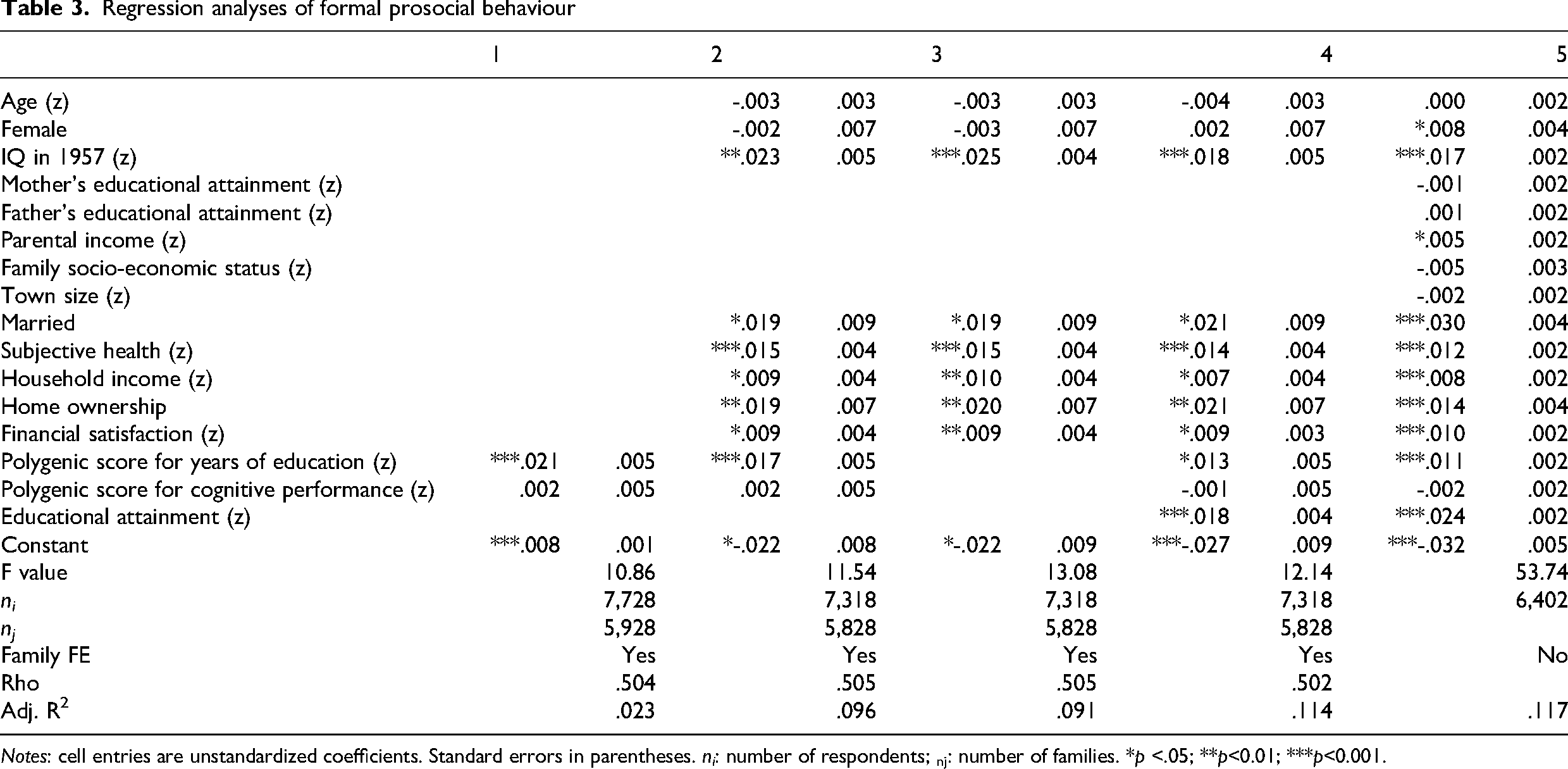
Notes: cell entries are unstandardized coefficients. Standard errors in parentheses. ni: number of respondents; nj: number of families. *p <.05; **p<0.01; ***p<0.001.
The first model shows that the polygenic score for educational attainment is positively associated with prosocial behaviour in formal contexts when shared environmental influences are captured by family fixed effects. The coefficient is the same in a model without family fixed effects (see online Appendix Table S2 in the Supplemental materials). We find no association with the polygenic score for cognitive performance, once the polygenic score for educational attainment is included.
The second model shows that the polygenic score for educational attainment remains associated with prosocial behaviour in formal contexts when covariates are added. The polygenic score for cognitive performance did not change as a result of adding IQ in 1957 to the model. The 1957 IQ score also is positively associated with prosocial behaviour in 2004. Adding the two polygenic scores – primarily the one for educational attainment – increases the proportion of variance explained with 0.005% compared to the third model with covariates only.
Hypothesis 2. The fourth model added educational attainment. The model explains 11% of the variance. Educational attainment was positively related to prosocial behaviour in formal settings. The association between the polygenic score for educational attainment and engagement in formal prosocial behaviour was attenuated by 38%. This result is in line with our hypothesis.
Hypothesis 3. At the same time, the association remained sizeable and significant. This indicates that genetic variants associated with educational attainment are also related to prosocial behaviour through other pathways than through educational attainment itself. This result is in line with our hypothesis.
A comparison of the association between educational attainment and prosocial behaviour in formal settings in Models 4 (0.018) and 5 (0.024) shows that it is weaker by about one-quarter when shared environmental influences are controlled by family fixed effects. However, a test for the equality of coefficients for educational attainment in Models 4 and 3 indicates that the difference (−0.006; SE: 0.004) is not statistically significant (z = −1.43, p = 0.152). Also, we find that the association between IQ in 1957 and prosocial behaviour is similar (0.018 vs 0.017) when shared environmental influences are controlled by family fixed effects. These results suggest that the association of prosocial behaviour in formal settings with educational attainment is partly due to shared environmental effects, but the association with cognitive performance is not.
In a model with only family characteristics, we find that family socio-economic status, father's educational attainment and parental income are positively associated with formal prosocial behaviour (see Model 1, online Appendix Table S9 in the Supplemental materials). Adding the polygenic scores but excluding IQ in 1957, educational attainment and covariates measured in 2004 reduces the associations with family socio-economic status and father's education (see Model 4, online Appendix Table S2 in the Supplemental materials). Adding IQ in 1957 (see Model 5, online Appendix Table S2 in the Supplemental materials) does not alter the associations with measured family background characteristics. Taken together, these results indicate that family socio-economic status, father's education and parental income are three shared environmental influences on formal prosocial behaviour, mediated by the educational attainment of respondents.
At the same time, the association between the polygenic score for educational attainment and formal prosocial behaviour (0.011) does not change much when shared environmental effects are not controlled. If anything, the association is somewhat weaker (−18%). These findings indicate that shared environmental effects may suppress the influence of the polygenic score, while they clearly amplify the effects of educational attainment.
Hypothesis 4. The results of analyses of informal prosocial behaviour are quite different: we find no associations between educational attainment and informal prosocial behaviour (see online Appendix Table S3 in the Supplemental materials), indicating that there is no educational gradient in helping others directly, without the involvement of an organisation. These results are at odds with our hypothesis. At the same time, we do find weakly positive associations with the polygenic score for educational attainment.
Conclusion
We examined associations among polygenic scores for educational attainment and cognitive performance with prosocial behaviours of Wisconsin graduates, tracked since 1957. We found that a polygenic score for educational attainment is associated with prosocial behaviour in formal settings: participants with more genetic variants associated with educational attainment are more intensively engaged in volunteering, more likely to give money to charities and to have ever given blood during their lifetime.
Our findings suggest that the benefits of educational attainment are not confined to the private lives of graduates but rather extend to the communities to which they contribute valuable resources as donors of blood, money and time. Controlling for polygenic scores for educational attainment and cognitive performance, we continue to find a robust association between educational attainment and prosocial behaviour. To some extent, this association may be due to genetic effects on other traits that are selected for in education.
The results show that genetic effects on educational attainment explain a small proportion of the variance in prosocial behaviour: by itself, the polygenic score for educational attainment explained ∼2% of the variance in formal prosocial behaviour. The association was reduced by 38% when educational attainment was added, suggesting that participants with a higher genetic propensity for educational attainment give more time, money and blood partly because they attain a higher level of education. The polygenic score for educational attainment also had a direct positive association with formal prosocial behaviour, indicating that there are other pathways than educational attainment through which the polygenic score is associated with prosocial behaviour. These results are consistent with hypotheses 1 and 2.
Educational attainment did not explain any variance in informal prosocial behaviour – providing emotional and social support and helping family, friends, neighbours and co-workers with daily housework, babysitting or childcare. These results are consistent with hypothesis 4.
The results provide partial support for the selection model, which argues that associations between educational attainment and prosocial behaviour are the result of a sorting process in the educational system, based on shared environmental and genetic factors that facilitate both educational attainment as well as prosocial behaviour. Consistent with this explanation, we find that the association between the polygenic score for educational attainment and prosocial behaviour is substantially reduced once educational attainment is taken into account. In addition to these genetic effects, shared environmental influences are prominent in formal prosocial behaviour. Our findings suggest that parental income, father's education and family socio-economic status are positively associated with prosocial behaviour measured 47 years later. However, we did not find support for the prediction in hypothesis 3 that parental education would remain associated with prosocial behaviour above and beyond the polygenic score for educational attainment and the level of education attained. Unexpectedly, we found that another shared environmental factor, parental income, remained associated with engagement in formal prosocial behaviour even when polygenic scores, cognitive performance, educational attainment and covariates were included.
At the same time, the results do not rule out causative effects of educational attainment on prosocial behaviour. We find that educational attainment is still correlated with prosocial behaviour when a polygenic score is included, even in models including family fixed effects. This finding indicates that educational attainment is not only associated with prosocial behaviour because of genetic effects and shared family background effects on educational attainment, but also in other ways.
The implication of co-twin control studies showing limited evidence for unique environmental effects of education is that the pathways creating the association between educational attainment and prosocial behaviour are likely to be due to both genetic and shared environmental effects. Indeed, we find that the coefficient for educational attainment is reduced by one-quarter when shared environmental effects are controlled. At the same time, we find that the higher level of engagement in formal prosocial behaviour in 2004 by respondents with higher cognitive performance in 1957 is not at all affected by shared environmental characteristics.
Discussion
The association between the polygenic score for educational attainment and prosocial behaviour is limited to formal prosocial behaviours. This finding is consistent with previous research showing that educational attainment is not consistently related to informal prosocial behaviour (Ramaekers et al., 2022; Wilson and Musick, 1997) and implies that informal helping behaviour occurs in all educational strata. The lack of an educational gradient in informal helping is informative because informal helping is strongly correlated with formal helping behaviour (r = 0.48), which must be due to other factors than educational attainment, such as social values or personality characteristics.
The proportion of variance in prosocial behaviour explained by the polygenic score for years of education is relatively small (0–2%). As the number of SNPs in GWAS of educational attainment increases, the association of the polygenic score with educational attainment will also increase (Abdellaoui and Verweij, 2021). To the extent that the association between educational attainment and outcomes such as prosocial behaviour reflects genetic effects, the proportion of variance explained by more comprehensive polygenic scores will also increase.
To some extent, the association of the polygenic score for educational attainment with prosocial behaviour is likely to result from unmeasured genetic effects on other traits. Genetic variants associated with complex traits, identified in large GWAS can be combined into composite measures of so-called polygenic risk scores to predict variation in these complex traits. These polygenic scores typically have small effects and hence contribute only a small fraction to the explained variance (Choi et al., 2020; Dulbridge, 2013). Our value of explained variance is comparable to similar studies explaining complex traits (cf. Lee et al., 2018; Mills et al., 2018). So-called pleiotropy refers to genes that influence several seemingly unrelated traits and create co-occurrence between traits, pointing to underlying shared genetic mechanisms (Watanabe et al., 2019). Educational attainment shows genetic correlations with many traits that are associated with prosocial behaviour, such as fertility, health and income (Mills and Tropf, 2020). We did not include polygenic scores for these traits in the current analyses. This implies that the associations of subjective health, income and financial satisfaction with formal prosocial behaviour should be regarded as reflecting both genetic and unique environmental effects.
A first limitation of our study is that we did not investigate paths from the polygenic score for educational attainment to prosocial behaviour beyond educational attainment. Future research should investigate which traits may explain the remaining association between educational attainment and prosocial behaviour in formal settings. We suggest that personality traits such as neuroticism and self-efficacy are likely to facilitate both educational attainment and prosocial behaviour in formal settings. Neuroticism is associated with lower educational attainment (Smith-Woolley et al., 2019) and shows negative associations with volunteering and blood donation (Bekkers, 2006). Self-efficacy contributes to achieving a higher level of education (Möller et al., 2020) and volunteering (Son and Wilson, 2017). Future research could include additional polygenic scores – for example, for personality characteristics – to identify such genetic effects.
A second limitation is that the cohort of Wisconsin 1957 graduates does not necessarily generalise to the US population or to other countries with different educational systems. As educational systems differ between countries and have changed over time, and prosocial behaviour has changed as well, its associations with educational attainment, genetic variants and shared environmental background will also have changed. Therefore, we call for replications in other cohorts, contexts and countries with the available measures of prosocial behaviour and polygenic scores for educational attainment. Future research on the associations of prosocial behaviour with educational attainment may also consider related traits such as empathic concern, altruism or co-operation, for which GWAS and polygenic risk scores may become available.
A third limitation is that measurement error may bias the estimates, particularly in self-reported variables such as educational attainment, parental education and father's occupation. While previous studies have shown that measurement error in such self-reports are small (Angrist and Krueger, 1999; De Vries et al., 2008), they can still reduce the precision and strength of the estimates. Because the measure of parental income in the WLS was taken from income tax registers, its association with prosocial behaviour was more precisely estimated. Preferably future research relies on observed characteristics, for instance through register data, as much as possible.
Taken together, our study has implications for theory, future research and policy. The theoretical implication of the finding that educational attainment is not related to informal prosocial behaviour is that explanations of the association between educational attainment and prosocial behaviour should take intermediate organisations into account. The lack of an educational gradient in informal prosocial behaviour suggests that educational attainment prepares citizens for prosocial behaviour benefiting society through intermediary organisations. The theoretical implication of the finding that shared family background and genetic variants are responsible for a substantial part of the educational gradient in formal prosocial behaviour is that the pre-adult socialisation model of political participation (Persson, 2015) also holds for formal prosocial behaviour. Thus, future research should investigate pathways from environmental influences that siblings in higher educational background families share with each other. Also, future research should consider potential genetic confounding, not only including educational attainment but also additional personality characteristics to shed light on the antecedents of prosocial behaviour in other samples and contexts.
At the same time, an educational gradient in formal prosocial behaviour remains that cannot be explained by pre-adult socialisation factors. This finding suggests that educational experiences and other environmental benefits of education contribute to prosocial behaviour, implying that collective investments in education are not solely resulting in individual benefits for those attaining higher levels of education, but also result in civic returns to the benefit of all.
Supplemental Material
sj-docx-1-asj-10.1177_00016993251409671 - Supplemental material for Learning to give: Both genetic and shared environmental influences are reflected in the association between educational attainment and prosocial behaviour
Supplemental material, sj-docx-1-asj-10.1177_00016993251409671 for Learning to give: Both genetic and shared environmental influences are reflected in the association between educational attainment and prosocial behaviour by René Bekkers, Eva-Maria Merz and Ting Li in Acta Sociologica
Supplemental Material
sj-docx-2-asj-10.1177_00016993251409671 - Supplemental material for Learning to give: Both genetic and shared environmental influences are reflected in the association between educational attainment and prosocial behaviour
Supplemental material, sj-docx-2-asj-10.1177_00016993251409671 for Learning to give: Both genetic and shared environmental influences are reflected in the association between educational attainment and prosocial behaviour by René Bekkers, Eva-Maria Merz and Ting Li in Acta Sociologica
Footnotes
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the H2020 European Research Council (grant number 802227 to Eva-Maria Merz).
Data availability statement
The data analysed in this article are available at https://researchers.wls.wisc.edu/documentation. A replication package including a pre-analysis plan for this study, the code producing the results reported here and appendices with additional results is available at ![]() .
.
Supplemental material
Supplemental material for this article is available online.
Author Biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.