
Abstract
Nanoparticles and bacteria have received a great attention worldwide due to their ability to remove heavy metals (HMs) from wastewater. The current study is aimed at finding the interaction of HMs-resistance strains (Bacillus cereus and Lysinibacillus macroides) with different concentrations (5, 10, 15, 20, and 25 mg/L) of zinc oxide nanoparticles (ZnO NPs) and how they would cope with HM stress (Pb, Cd, Cr, and Cu). The growth rate and tolerance potential of bacteria were increased at lowered concentrations (5 and 10 mg/L) of ZnO NPs against HMs while it was unaffected at higher concentrations of ZnO NPs. These findings were confirmed by minimum inhibition zone and higher zinc solubilization at lower concentrations of ZnO NPs. Scanning electron microscopy (SEM) revealed that higher concentrations of ZnO NP increased HM accumulation in bacteria cells which had a significant impact on bacterial morphology and caused pores in bacterial membrane while in the case of lower concentrations, the cell remained unaffected. These results were further supported by the less production of antioxidant enzymes (SOD, POD, and CAT), thiobarbituric acid reactive substances (TBARS), and hydrogen peroxide (H2O2) contents at lower concentrations of ZnO NPs against heavy metal stress. This study suggested that synergistic treatment of Bacillus spp. with lower concentrations of ZnO NPs enhances the tolerance potential and significantly reduces the HM toxicity.
1. Introduction
Various methods such as ion exchange, metal precipitation, adsorption methods, and reverse osmosis were used in the past for the removal of HMs but these methods were ineffective, costly, and time-consuming [1, 2]. After that, microbe-based remediation was used to remove heavy metal stress. Bacteria have different detoxifying mechanism, such as biosorption, bioaccumulation, biotransformation, and biomineralization to alleviate the HM stress [3]. However, solubilized state of HMs is not quickly and effectively removed by bacteria [4]. So as a result, there was a great demand to use adsorbents, which have efficient heavy metal binding capabilities and help in the removal of HMs. ZnO NPs have wide application because of their uses in optical systems, solar cells, and storage devices [5]. Similarly, their ability to absorb ultraviolet radiation makes them an essential component in enzymes, sunscreens, toothpastes, and cosmetics [6, 7]. ZnO NPs, on the other hand, have previously been shown to be capable of removing lead (Pb) and cadmium (Cd) from aqueous solutions in different conditions, including time, absorbent dosages, pH, and temperature [4]. ZnO NPs have a porous micro/nanostructure with variety of surface active sites that can effectively attach with HMs and help to grow bacteria faster [8]. Because of their electropositive nature, these nanoparticles have a stronger attraction towards electronegative charge metals including Ag- and S- ions [9]. On the other hand, some Bacillus spp. use lower concentrations of ZnO NPs as Zn ions and solubilize in the cell membrane by making a complex with proton and chelated ligand [10]. Microbes use Zn ions as a cofactor in their metabolic process to meet the zinc requirements for growth. These zinc ions are an essential component for bacteria’s growth and have a beneficial impact on enzymes such as dehydrogenase, thiol peroxidase, and glutathione reductase and increase the bacteria’s tolerance against HMs [11].
Various studies have shown that bacteria remediate the HMs from wastewater but no report has been published, to best of our knowledge, about the combined effect of ZnO NPs and bacteria on wastewater remediation. The present study was therefore conducted to determine the effect of various concentrations of ZnO NPs on Bacillus spp. growth against heavy metal stress by investigating biochemical and antioxidant parameters of bacteria and also observe the combined role of ZnO NPs and bacteria in wastewater remediation.
2. Materials and Methods
2.1. Zinc Oxide Nanoparticles (ZnO NPs)
ZnO NPs were obtained from Pir-Mehr-Ali-Shah (PMAS) Arid Agriculture University, Rawalpindi. It was synthesized by coprecipitation method by using the protocol of Hussain et al. [12] with some modification. ZnO nanoparticles were prepared by mixing 0.05 M Zinc acetate Zn (NO3)2.6H2O (25 mL) solution with 4 mL of plant extract. Solution was heated and continuously stirred. The solution was centrifuged at 12,000 rpm for 15 min. Supernatant was discarded, and the isolated pellet was again suspended in deionized water. The solution was again centrifuged for 5 min and repeated the process three times to remove impurities. ZnO NPs were characterized by Amara et al. [13] through Field Emission Scanning Electron Microscope (FESEM), Transmission Electron Microscopy (TEM), Fourier Transform Infrared Spectroscopy (FTIR), and X-ray diffraction (XRD). ZnO NPs (1 g) was dissolved in 1000 mL deionized water (Milli-Q water) (Sigma-Aldrich, St. Louis, CA, USA) to make a stock solution. The ZnO NP solution was placed in an ultrasonic bath for 30 minutes to break up large aggregates of NPs and made the homogenized solution.
2.2. Bacterial Strain Collection
The bacterial strains Bacillus cereus (PMBL-3) and Lysinibacillus macroides (PMBL-7) were identified by Khattak et al. and then obtained from Plant and Microbial Biotechnology Lab, Kohat University Science Technology, Kohat, Pakistan [14].
2.3. Bacteria Strain Growth
Bacterial strains were grown in a nitrogen broth (0.5% of yeast extract (Sigma-Aldrich, St. Louis, CA, USA), 1% of tryptone (Sigma-Aldrich, St. Louis, CA, USA), and 1% of sodium chloride (Sigma-Aldrich, St. Louis, CA, USA)) dissolved in DW. The pH of the media was 7.4-7.6 at the end. The bacteria were inoculated in autoclaved media from a fresh culture. The bacteria-based culture broth was then centrifuged at 3500 × g for 10 minutes. The pellet contained bacteria biomass was properly washed with physiological saline water. Now these fresh bacterial cells were suspended in 10 mL of sterilized saline water (9 g NaCl) (Sigma-Aldrich, St. Louis, CA, USA) in 1 L DW.
2.4. Bacteria and ZnO NP Interaction/Exposure Experiment
The methodology of batch culture experiment was design by the protocol as described previously by Wan et al. [15]. In a brief, 20 mL of different concentrations (5, 10, 15, 20, and 25 mg/L) of ZnO NPs was sequentially placed in 50 mL sterile Erlenmeyer flasks. From each fresh cultured bacteria strain, 1 mL of suspension was dissolved in 20 mL of DW and then added in flasks containing various concentrations of ZnO NPs. Each flask was sterilized and then heated in a rotatory shaker (Perkin Elmer, Analyst 4000, Waltham, MA, USA) (200 rpm) for 90 minutes at 30°C. The experiment was consists of one positive control (bacteria culture without ZnO NPs) and one negative control (no bacteria culture with only ZnO NPs). This experiment was carried out in replicates. Subsequently, after exposure with ZnO NPs, bacterial suspensions were obtained and their tolerance thresholds were determined against each HM salt (PbCl2, CdCl2.2.1/2H2O, K2Cr2O7, and CuSO4.5H2O).
2.5. Resistance/Tolerance Level of Bacteria against Heavy Metals
The tolerance level of bacterial against HMs (Pb, Cd, Cr, and Cu) was examined by the methodology as described previously by Dhas et al. [16]. After the exposure experiment, bacterial strain’s cells (
2.6. Optical Density (OD)
Bacterial optical density against HMs was measured by using methodology of Peters et al. [17]. From the batch culture assay, bacterial strains (
2.7. Bacterial Growth/Colony-Forming Unit (CFU)
The CFU of bacteria was measured by following the methodology of Hsueh et al. [8] with slight changes. After the exposure experiment, small aliquots (
2.8. Zinc Solubilization Assay
Zinc solubilization experiment was carried out in a mineral salt medium (g/L), as mentioned previously by Saravanan et al. [18]. Bacterial strains were inoculated in mineral salt medium containing dextrose (NH4)2 SO4: KCl:; K2HPO4:; MgSO4 (pH: 7); insoluble zinc compound from ZnO NPs (5, 10, 15, 20, and 25 mg/L); insoluble zinc compound from ZnO NPs (5, 10, 15, 20, and 25 mg/L); and 0.1 percent agar. Fresh bacteria culture (3 L) was spotted in each prepared agar plate containing 100 mg/L of HMs (Pb, Cd, Cr, and Cu) and incubated at 28°C for 48 hours. The zinc solubilization zone was measured in millimeter (mm).
2.9. Zone Inhibition Assay
Combined impact of ZnO NPs-bacteria in a well diffusion experiment was determined by the protocol of Dwivedi et al. [19]. Bacteria were grown in nutrient agar plates, amended with 100 mg/L of HMs (Pb, Cd, Cr, and Cu). After that, plates were solidified and 10 mm sterile cork wells were made. In each well, 100 μL of different concentrations (5, 10, 15, 20, and 25 mg/L) of ZnO NPs was poured in each well and incubated overnight at 30°C. The zone of inhibition in each well was measured (mm) in the next day.
2.10. Bacteria-ZnO NP Interaction
The interaction of bacteria-ZnO NPs in HIE polluted water was studied by Kumar et al. [20]. Various concentrations (5, 10, 15, 20, and 25 mg/L) of ZnO NPs were arranged sequentially in sterile Erlenmeyer flasks (25 mL). Each flask contained an equal amount (25 mL) of freshly cultured bacteria broth and HIE-derived wastewater. Flasks were sonicated for 2 minutes to remove any residual aggregates of ZnO NP. Each flask was sterilized and incubated at 30°C for 24 hours in a shaking incubator (200 rpm). Furthermore, mixture was centrifuged, and the supernatant was removed while the pellet containing the bacterial biomass was screened for HM aggregation and antioxidative enzymatic activities.
2.11. Bioaccumulation of HMs
To determine the HM content in bacterial biomass, the bacterial suspension (2 mL) from each flask was removed and washed twice with sterilized physiological saline (10 mM EDTA-Na2) (Sigma-Aldrich, St. Louis, CA, USA) [21]. The media and bacteria biomass were separated by centrifuging the bacteria suspension at 10,000 rpm. Samples were mixed with 68% HNO3 at 110°C before the solution becomes completely clear. The digestion solution was transferred to a flask and added 2% HNO3 while raised to a final volume by adding10 mL of DW. Atomic absorption spectroscopy was used to examine the concentration of HMs (Pb, Cd, Cr, and Cu) in digested solution (PerkinElmer, Analyst 4000).
2.12. Antioxidant Parameters
Antioxidant enzymes in bacteria biomass were examined by Peters et al. [17]. The bacterial suspension was centrifuged at 12,000 rpm (4°C) for 10 minutes to extract the bacteria biomass. Bacteria biomass was combined with 100 mL of potassium phosphate buffer (pH 7.5) consists of 1 mL of EDTA (Sigma-Aldrich, St. Louis, CA, USA), 3 mL of dithiothreitol (DTT) (Sigma-Aldrich, St. Louis, CA, USA), and 5% polyvinylpyrrolidine (PVP) (Sigma-Aldrich, St. Louis, CA, USA). After that, enzyme was obtained by centrifuging the mixture at 20,000 rpm for 15 minutes. Each antioxidative enzyme was evaluated using the protocols mentioned below.
2.13. Superoxide Dismutase (SOD)
The activity of superoxide dismutase (SOD) in bacteria biomass was investigated by Gonzalez et al. [22]. Substrate was made by adding 15.5 mg of nitro blue tetrazolium (NBT) (Sigma-Aldrich, St. Louis, CA, USA), 0.02 mg of riboflavin (Sigma-Aldrich, St. Louis, CA, USA), 10 mg of sodium ethylene diamine tetra acetic acid (Na2 EDTA) (Sigma-Aldrich, St. Louis, CA, USA), and 485 mg of methionine (Sigma-Aldrich, St. Louis, CA, USA). The substrate was covered and kept at 4°C to prevent the degradation. In the flask, 2.25 mL of reaction substrate and 25 mL of enzyme extract were mixed to give the reaction mixture. The mixture was exposed to UV light for 20 min and then taken in the dark to complete the reaction. The SOD content was then measured at 560 nm by using a spectrophotometer (UV-visible spectrophotometer).
2.14. Peroxidase (POD)
The peroxidase content in bacteria biomass was determined by Vignali et al. [23]. Enzyme extract (100 mL), 100 mL of 1.5% guiacol (Sigma-Aldrich, St. Louis, CA, USA), 100 mL of 4% hydrogen peroxide (Sigma-Aldrich, St. Louis, CA, USA), and 2.75 mL of reaction substrate were mixed to make 3 mL of reaction substrate (50 mM of potassium phosphate buffer) (Sigma-Aldrich, St. Louis, CA, USA). After 10 minutes, the absorbance was measured at 470 nm at room temperature (RT).
2.15. Catalase (CAT) Activity
The catalase content in bacteria biomass was determined by Den Besten et al. [24] method. For this purpose, reaction substrate was made in a cuvette by adding 2.5 mL of 100 mM hydrogen peroxide (Sigma-Aldrich, St. Louis, CA, USA), 100 μl of 25 mM potassium phosphate buffer (PPB) (Sigma-Aldrich, St. Louis, CA, USA), and 100 μl of the enzyme extract. CAT content was determined at 240 nm.
2.16. Thiobarbituric Acid Reactive Substances (TBARS)/Malondialdehyde (MDA) Level
The levels of thiobarbituric acid reactive substances (TBARS) and malondialdehyde (MDA) in bacterial biomass were used to investigate lipid peroxidation. MDA content was analyzed by following the protocol of Hasanuzzaman et al. [25]. Substrate reaction was formed by adding 50% v/v of thiobarbituric acid (Sigma-Aldrich, St. Louis, CA, USA) and 10% v/v of trichloroacetic acid (Sigma-Aldrich, St. Louis, CA, USA) to extract enzyme and boiled at 95°C for 30 minutes. Afterwards, the mixture was cooled by keeping the flask in a cold environment. The photospectrophotometer (Perkin Elmer, Analyst 4000, Waltham, MA, USA) was used to determine the MDA content at 532 and 600 nm.
2.17. Hydrogen Peroxide
H2O2 content in bacteria biomass was determined by Hossain et al. [26]. The substrate was made by adding 1 mL of 50 mM PBS, 1 mL of 25 mM potassium iodide (Sigma-Aldrich, St. Louis, CA, USA), and 1 mL of enzyme extract. The H2O2 content was determined by recording the absorbance at 390 nm.
2.18. Examination of Bacteria Cell Morphology by Scanning Electron Microscopy (SEM)
Bacteria cell morphology was examined using the scanning electron microscopy (SEM) by using the method as described previously by Ghinwa et al. [27] with slight modification. LB media was used to grow Bacillus cereus strain overnight. In next day, the OD of the sample was determined by removing midlog stage cells (109 cells/mL) from the bacteria culture and diluting it until it had an OD less than 0.6 and contained 107 cells/mL. In bacteria broth media, different concentrations (5, 10, 15, 20, and 25 mg/mL) of ZnO NPs were made. After that, the reaction mixture was made by mixing 20 mL of bacteria suspension with varying concentration of ZnO NPs (final volume: 2 mL). The mixture was centrifuged at 15,000 rpm for 5 minutes to remove debris. After gentle shaking, samples were passed through dialyzed membranes in 4 L Milli-Q deionized water at room temperature (30 minutes) for high-quality SEM imaging. To preserve the morphology of the organisms, the bacteria culture was fixed in 4% PFA fixing solution (Sigma-Aldrich, St. Louis, CA, USA) for 2 hours. Cell suspension (200 μl) was spotted on glass coverslips in a DCP with a liquid CO2 ethanol exchanger and air-dried (Dentem vacuum critical point dryer) for 1 hour. The coverslip was attached to the SEM stubs with carbon adhesive tabs. The coverslip was fixed to the SEM stubs with carbon adhesive tabs (LUXORTM benchtop sputter coater). Scanning electron microscopy (SEM) (JSM5910, JEOL, Japan) and digital images of bacteria cells were produced at 25,000 magnification with (10 kV) accelerated voltage.
2.19. Statistical Analysis
Samples were statistically analyzed in the Statistic 9 software using the ANOVA (analysis of variance) test and LSD (least significant difference) at
3. Results
3.1. Effect of ZnO NP on Bacterial Growth
We first investigated the effect(s) of ZnO NPs on the bacterial growth. Different doses of ZnO NPs were used to treat the bacterial strains. Higher concentrations of ZnO NPs inhibited the growth of bacterial strains (Figures 1(a) and 1(b)). As a result, we used lower concentrations (5-15 mg/L) of ZnO NPs in subsequent studies, but higher concentrations were used as a control. In the first experiment, we see whether ZnO NPs can increase bacteria’s resistance against different HMs. Bacterial strains were exposed to different concentrations of HMs to determine the HM tolerance level. It was observed that B. cereus showed a maximal tolerance against Pb (300 ppm) as compared with Cr, Cd, and Cu (125, 175, and 250 ppm), respectively. On the other hand, ZnO NPs, at lower concentrations, greatly increased bacteria resistance (see Table 1). Importantly, in contrast to the control treatment, bacteria tolerance to HMs was greatly improved at 15 mg/L NP concentrations against Pb, Cr, Cd, and Cu stress (400, 225, 275, and 275 ppm), respectively. In contrast, higher concentrations of ZnO NPs (20-25 mg/L) had no effect on bacterial tolerance as compared with lower concentrations (Table 1). Similar results were obtained in the case of L. macroides where lower concentrations of NPs showed maximum tolerance against HMs as compared with control (Table 1).

Tolerance level of bacteria strains (a) Bacillus cereus and (b) Lysinibacillus macroides under different concentrations of ZnO NPs (5, 10, 15, 20, and 25 mg/L).

Effect of different concentrations of ZnO nanoparticles (0, 5, 10, 15, 20, and 25 ppm) on maximum inhibitory concentration (MIC) of Bacillus cereus and Lysinibacillus macroides under heavy metal (Pb, Cd, Cr, and Cu) stress.
We previously observed that lower concentrations of ZnO NP increased bacterial growth. We tried to see if these concentrations could make bacteria grow better when they were exposed to HMs. All bacterial strains (B. cereus and L. macroides) were exposed to a fixed volume (100 mg/L) of HMs in the presence of NPs, and their optical density (OD) was measured at different times. We noted that bacterial growth was severely inhibited at different times (5, 10, 15, 20, and 25 h) under HM (Pb, Cd, Cr, and Cu) stress as compared with control untreated samples (Figures 2(a) and 2(b)). For example, it is evident from Figures 2(a) and 2(b) that B. cereus (0.374, 0.312, 0.353, and 0.232) and L. macroides (0.245, 0.311, 0.332, and 0.353) growth was inhibited after 20 h as compared with untreated bacteria: B. cereus (0.416, 0.521, 0.254, and 0.332) and L. macroides (0.354, 0.365, 0.343, and 0.327 OD600). Surprisingly, we found that bacteria grew faster when exposed to lower concentrations of ZnO NPs (Figures 2(a) and 2(b)). Furthermore, it is also clear from Figures 2(a) and 2(b) that lower concentrations of NPs enhanced bacterial growth against HM stresses. However, the OD measurements should be sufficient to conclude that lower concentration of ZnO NPs will promote bacterial growth. To further confirm, the colony-forming unit (CFU) of bacteria was determined which revealed that the number of viable cells was maximum at lower concentration of ZnO NPs under HM stress. It is clear from Table 2 that the CFU of bacteria was increased at lower concentrations of ZnO NPs in the presence of HMs as compared with control (Table 2). Furthermore, the common pattern of growth was observed for both bacterial strains under HM stress. On the other hand, higher amount of ZnO NPs increased the bacterial growth as compared with bacterial cells treated only with HMs. As a result, our OD and CFU results are consistent, indicating that lower concentrations of NPs had greatly increased bacterial growth under HM stress (see Table 2).

Effect of ZnO NPs (0, 5, 10, 15, 20, and 25 mg/L) on the growth of (a) Bacillus cereus and (b) Lysinibacillus macroides under heavy metal stress: (A) Pb, (B) Cd, (C) Cr, and (D) Cu at different time interval (0, 5, 10, 15, and 20 h). Error bars show means of standard error (± S.E) of three replicates (

Effect of different concentrations of green synthesized ZnO nanoparticles (0, 5, 10, 15, 20, and 25 ppm) on colony-forming unit (CFU) of Bacillus cereus and Lysinibacillus macroides under heavy metal (Pb, Cd, Cr, and Cu) stress.
Values are
As noted in the previous section, lower concentrations of ZnO NPs increased bacterial growth under HM stress. ZnO NPs contained zinc ions which have been reported to have the ability to serve as a micronutrient for bacterial growth. To demonstrate this, we used the zinc solubilization assay. It is clear from Table 3 that lower concentrations of ZnO NPs resulted in increased solubilization of zinc ions under HM stress than higher concentrations of NPs. Consequently, maximum solubilization was observed at 5 mg/L of ZnO NPs in B. cereus (1.4, 1.3, 1.1, and 1.2 mm) and L. macroides (1.1, 1.3, 1.2, and 1.4 mm) under various HM stresses as compared with control treatments (1.5 and 1.3 mm), respectively. Interestingly, solubilization ability of bacteria was decreased at 25 mg/L of ZnO NPs in B. cereus (1.2, 1.1, 1.2, and 1.2 mm) and L. macroides (1.1, 1.1, 1.1, and 1.2 mm) under HM stress as compared with control treatments (1.8 and 1.6 mm), respectively (Table 3).
Effect of different concentrations of ZnO nanoparticles (0, 5, 10, 15, 20, and 25 ppm) on zinc solubilization of Bacillus cereus and Lysinibacillus macroides under heavy metal (Pb, Cd, Cr, and Cu) stress.
Values are
Our zinc solubilization assay and previous studies had shown that NPs enhanced bacterial growth in the presence of HMs. Alternatively, we hypothesized that NPs could reduce the bacteria’s toxicity to HMs. To achieve this, we used a zone inhibition/well diffusion assay for HMs. HMs developed a wide inhibition zone under HM stress in the absence of NPs (Table 4). However, the zones were significantly decreased in the presence of lower concentration of ZnO NPs (Table 4). Large diffusion zone was observed in B. cereus (3.1, 3.8, 2.6, and 3.4 mm) and L. macroides (2.4, 3.9, 2.5, and 2.4 mm) for HMs (Pb, Cd, Cr, and Cu) in the absence of NPs. On the other hand, lower concentrations (5 mg/L) resulted a small inhibition zone in B. cereus (2.5, 2.7, 2.2, and 3.1 mm) and L. macroides (1.8, 2.4, 1.8, and 1.2 mm) under HMs (Pb, Cd, Cr, and Cu). Furthermore as previously noted, the inhibition area was maximized at higher concentrations of ZnO NPs (Table 4).
Effect of different concentrations of ZnO nanoparticles (0, 5, 10, 15, 20, and 25 ppm) on zone of inhibition of Bacillus cereus and Lysinibacillus macroides under heavy metal (Pb, Cd, Cr, and Cu) stress.
Values are
The ability of NPs to reduce the zone of inhibition had been shown in a well diffusion assay. To further confirm that, bioaccumulation assay was conducted. It can be seen in Figure 3 that maximum accumulation of HMs was observed in both B. cereus and L. macroides in the presence of HM stress. Additionally, Cr (1.37 and 0.945 mg/g) accumulation was greater than Pb, Cd, and Cu accumulation in B. cereus (1.15, 0.611, and 1.04 mg/g) and L. macroides (0.654, 0.543, and 0.765 mg/g), respectively. The synergistic effect enhanced bacteria’s ability to use HMs at lower concentrations as compared with higher concentrations of ZnO NPs. Maximum accumulation of HMs (Zn, Pb, Cd, Cr, and Cu) was observed in B. cereus (0.894, 1.298, 1.121, 0.995, and 0.432 mg/g) and L. macroides (0.543, 0.855, 0.743, 0.643, and 0.533 mg/L) at lowered concentration (5 mg/L) of ZnO NPs. Higher concentrations of ZnO NPs (25 mg/L) resulted in minimum accumulation of HMs in B. cereus (1.43, 0.622, 0.411, 0.475, and 0.51) and L. macroides (0.754, 0.532, 0.343, 0.423, and 0.501 mg/g) as compared with lower concentrations of NPs (Figure 3).

Effect of ZnO NPs (0, 5, 10, 15, 20, and 25 mg/L) on the heavy metal (lead, cadmium, chromium, and copper) uptake of (a) Bacillus cereus and (b) Lysinibacillus macroides. Error bars showed means of standard error (± S.E) of three replicates (

Lysinibacillus macroides
3.2. Effect of ZnO NPs on Bacterial Antioxidant Enzymes
3.2.1. Superoxide Dismutase (SOD)
Maximum SOD content in B. cereus and L. macroides was observed at different concentrations of HMs (Pb, Cd, Cr, and Cu) while synergistic treatment with ZnO NPs reduced HMs’ toxic effects and resulted in minimum SOD activity (Figures 4(a) and 4(d)). As compared to Pb, Cd, and Cu in B. cereus (0.318, 0.453, and 0.363) and L. macroides (0.612, 0.543, and 0.453 U/mg protein), Cr showed the highest SOD activity (0.506 and 0.632), respectively. SOD activity was significantly decreased in B. cereus (0.183, 0.254, 0.278, and 0.365 U/mg/protein) and L. macroides (2.450, 3.650, 3.890, and 5.320 U/mg/protein) at lower concentrations (5 mg/L) of ZnO NPs, as compared with higher concentrations (25 mg/L) of ZnO NPs (Figures 4(a) and 4(d)).
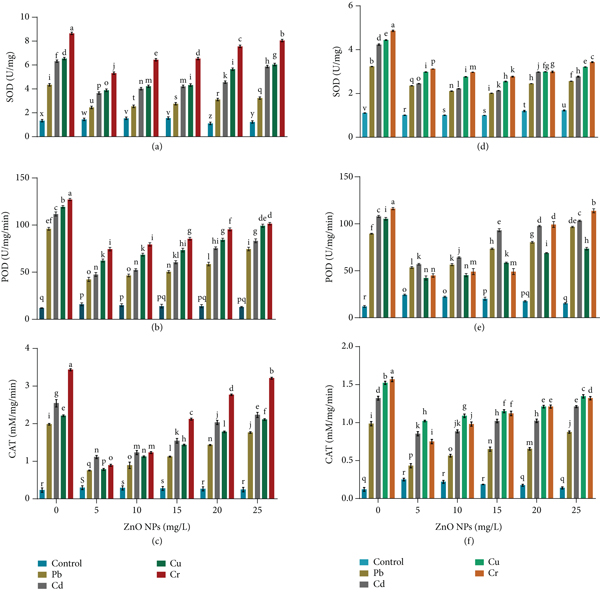
Effect of ZnO NPs (0, 5, 10, 15, 20, and 25 mg/L) on (a) superoxide dismutase, (b) peroxidase, (c) catalase of Bacillus cereus, (d) superoxide dismutase, (e) peroxidase, and (f) catalase of under heavy metal stress. Error bars show means of standard error (± S.E) of three replicates (
3.2.2. Peroxidase (POD)
The effect of HMs (Pb, Cd, Cr, and Cu) on B. cereus and L. macroides revealed maximum POD activity, whereas combined treatment with ZnO NPs reduced the harmful impact of HMs and resulted in the lowest POD activity. Similarly, our findings showed a maximum POD content under Cr stress (127.068 and 116.088) as compared with Pb, Cd, and Cu (96.026, 119.32, and 111.729) stress in B. cereus and L. macroides (107.899, 105.32, and 89.43 U/mg protein), respectively. POD content was also decreased in B. cereus (42.32, 47.54, 62.32, and 47.43 U/mg protein) and L. macroides (53.52, 56.95, 42.32, and 44.93 U/mg protein) under heavy metal (Pb, Cd, Cu, and Cr) stress as compared with higher concentration (25 mg/L) of ZnO NPs in B. cereus (74.43, 83.24, 99.23, and 101.423 μ/g) and L. macroides (89.43, 107.89, 105.32, and 116.08 μ/g), respectively (Figures 4(b) and 4(e)).
3.2.3. Catalase (CAT)
Catalase (CAT) contents were also increased in B. cereus and L. macroides under heavy metal stress while synergistic treatments with ZnO NPs showed minimum CAT content (Figures 4(c) and 4(f)). Results revealed that maximum CAT content was observed under chromium stress (3.434 and 1.568 mM/mg/min) as compared with Pb, Cd, and Cu in B. cereus (2.543, 2.21, and 1.981) and L. macroides (1.523, 0.985, and 1.321 mM/mg/min), respectively. Catalase (CAT) content was also decreased at minimum amount (5 mg/L) of ZnO NPs under Pb, Cd, Cu, and Cr stress in B. cereus (0.754, 1.112, 0.783, and 0.893) and L. macroides (1.764, 2.23, 2.11, and 3.21 mM/mg/min) as compared with higher concentration (25 mg/L) of ZnO NPs in B. cereus (0.432, 0.853, 1.021, and 0.750) and L. macroides (0.985, 1.321, 1.523, and 1.568 mM/mg/min), respectively (Figures 4(c) and 4(f)).
3.2.4. Thiobarbituric Acid Reactive Substances (TBARS)/Malondialdehyde (MDA) Level
Results revealed that MDA content was more increased under chromium stress (6.790 and 5.157) as compared with Pb, Cd, and Cu in Bacillus cereus (3.32, 4.87, and 5.33) and Lysinibacillus macroides (4.32, 3.21, and 4.98 nM/mg), respectively. MDA content was significantly reduced at lower concentration (5 mg/L) of ZnO NPs in Bacillus cereus (1.32, 2.32, 1.87, and 3.23 nM/mg) and Lysinibacillus macroides (1.76, 1.94, 2.87, and 3.23 nM/mg) while increased at higher concentration (25 mg/L) of ZnO NPs in Bacillus cereus (1.76, 1.94, 2.87, and 3.23 nM/mg) and Lysinibacillus macroides (3.21, 4.32, 4.98, and 5.15 nM/mg), respectively (Figures 5(a) and 5(c)).

Effect of ZnO NPs (0, 5, 10, 15, 20, 25 mg/L) on (a) malondialdehyde (MDA) and (b) hydrogen peroxide (H2O2) of Bacillus cereus and (c) malondialdehyde (MDA) and (d) hydrogen peroxide (H2O2) content of Lysinibacillus macroides under heavy metal (Pb, Cd, Cr, and Cu) stress. Error bars showed means of standard error (± S.E) of three replicates (
3.2.5. Hydrogen Peroxide (H2O2)
Hydrogen peroxide content was also increased in bacteria under HM stress (Figures 5(b) and 5(d)). Maximum H2O2 content was observed under chromium stress (3.76 and 11.755 nM/g) as compared with Pb, Cd, and Cu in Bacillus cereus (4.346, 6.11, and 5.903) and Lysinibacillus macroides (8.937, 5.633, and 7.43 μM/g), respectively. H2O2 content was significantly reduced at lower concentrations (5 mg/L) of ZnO NPs under Pb, Cd, Cr, and Cu stress such as Bacillus cereus (2.03, 2.21, 2.89, and 3.76) and Lysinibacillus macroides (3.11, 4.54, 3.43, and 5.43 μM/g) while increased at higher concentration (25 mg/L) such as Bacillus cereus (3.11, 4.54, 3.43, and 5.43 μM/g) and Lysinibacillus macroides (5.63, 7.905, 7.23, and 9.46 μM/g), respectively (Figures 5(b) and 5(d)).
3.2.6. Effect of ZnO NPs on Bacterial Morphology
In order to find the effect of different concentrations of NPs on bacterial morphology, we used scanning electron microscopy (SEM). Figure 6(a) showed bacteria with a rod-shaped shape in control. There was no visible distortion or internal deformation, indicating that the bacteria were healthy in control treatment. After that, we applied ZnO NPs to Bacillus cereus to see how they interact with the bacteria. After 12 hours of incubation, it was observed that lower concentrations of NPs (5 and 10 mg/mL) had no impact on bacterial morphology (Figures 6(b) and 6(c)) in the presence of a large number of NPs on the bacterial surface (shown with arrows). ZnO NPs at higher concentrations (15 mg/mL) adhered better to the bacterial surface, resulting in a small reduction in bacterial biomass (Figure 6(d)). However, higher concentrations of the NPs (20 and 25 mg/mL) greatly damage the bacterial morphology, resulting in bacterial cell deformation and formation of pore in the cell wall (Figures 6(e) and 6(f), shown with yellow arrows).

Effect of ZnO NPs (0, 5, 10, 15, 20, and 25 mg/L) on Bacillus cereus cell morphology in the midlog phase of growth after 12 h: (a) control, (b) 5 mg/L, (c) 10 mg/L, (d) 15 mg/L, (e) 20 mg/L, and (f) 25 mg/L of ZnO NPs.





4. Discussion
Nanoparticles have gained a lot of attention due to their morphological, textural, and structural properties [27]. Antimicrobial properties of nanoparticles, especially ZnO NPs, have previously been investigated [28]. However, there is only a limited literature available on the possible role of ZnO NPs in enhancing bacterial growth and its HM remediation ability. Therefore, the current study was initiated to determine the effect of ZnO NPs on bacterial growth, morphology, and its HM remediation capacity. We started by checking bacteria growth at different concentrations of NPs to find the working concentrations of NPs that could be used in future experiments (Figure 1). According to our findings, bacterial growth gradually increased at lower concentrations of ZnO NPs (5-15 mg/mL). On the other hand, higher concentration seems to be toxic to bacteria and limiting their growth on agar plates (Figure 1). This result is consistent with previous report, which suggest that higher NP concentrations can be detrimental to bacterial strains [29]. Surprisingly, our findings revealed that lower concentrations of ZnO NP increased the B. cereus and L. macroides resistance level against HMs (Table 1) confirmed by the findings of optical density (OD) (Figure 2) and colony-forming unit (CFU) (Table 2). These results showed that lower concentrations of ZnO NPs promote bacterial growth and increased resistance against HM stress These results are confirmed by other scientists [30], and they revealed that bacteria can convert ZnCl2 to Zn+2 ions, which are then used as an essential cofactor for bacteria’s structural and regulatory functions in toxic environments. Furthermore, bacteria have been shown to maintain Zn2+ homeostasis by importing and exporting Zn2+ through zinc efflux systems [31]. As a result, porin enables the passive absorption of zinc ions into cells, aiding in cellular ingestion of nutrients and inducing cell-cell contact [29]. Furthermore, zinc ions have been shown to bind with maltodextrin (a major source of sugar in bacteria) and promote its entry into bacterial cells through LamB and other porin proteins [30]. Bacteria use Zn+2 ions as micronutrients in the range of 0.01-1 mM to enlarge and maintain the membrane, macromolecules, numerous steroid receptors, and carbohydrate metabolism [32]. On the other hand, bacteria behave differently when exposed to high concentrations of ZnO NPs. For example, bacteria have been shown to activate the envelop stress response (ESR) to protect themselves against higher concentrations of NPs and HMs. Gram-positive bacteria integrate D-alanine into their cell walls during ESR system activation, reducing net negative charges on their surfaces and thereby reducing their contact with positively charged NPs. On the other hand, gram-negative bacteria have lipopolysaccharides (LPS) (Lipid A) on their cell membranes, which they added with phosphoethanol amine (PEA) to increase the surface’s positive charge and decrease NPs [33].
According to the zinc solubilization assay, bacteria solubilize zinc ions from ZnO NPs, at lower concentrations under higher HM stress (Table 3). Furthermore, well diffusion assay revealed that lower concentration of ZnO NP revealed minimum HM diffusion in agar plates (Table 4). According to previous studies, bacteria secrete metabolites, amino acids, protein extrusions, annealing agents, sulphuric acid, nitric acid, and ascorbic acid, to solubilize the zinc ions by forming a complex with proton and chelated ligands [29]. Furthermore, our findings are supported by the observation of Bong et al. [34] that Bacillus spp. produce organic acid by changing the pH of the culture broth and solubilizing the zinc ions. On the other hand, higher concentrations of NPs revealed the maximum inhibition zone by causing the cell surface saturation and destroying the cell membrane [35]. These findings were further confirmed by other report [36] that higher concentrations of ZnO NPs revealed a maximal inhibition zone for Pseudomonas aeruginosa and Escherichia coli. It was observed that lower concentrations of ZnO NP increased the efficiency of bacteria to accumulate maximum HMs as compared with the higher concentration (Figure 3). Bacteria have been shown to have an extracellular sequestration mechanism (in which bacteria secrete environment stress response (ERS) and immobilized toxic ions to prevent them from contact with other bacteria) to protect themselves against HMs by inducing intracellular sequestration proteins such as the SilG gene and SiL operons, which contain all toxic ions in the cytoplasm [37, 38].
Hormesis is a bacterial process that consists of two levels: an enzymatic level and a short-term method. The intra- and extracellular concentrations of HMs are lowered by scavenging enzymes in the enzymatic system, allowing bacteria to retain balanced ion concentrations and hence improved bacterial growth [28]. SOD, POD, and CAT play an important role in bacterial resistance mechanisms against oxidative stress. We observed that higher concentrations of ZnO NPs under HM stress content revealed maximum lipid peroxidation activity and hydrogen peroxide content, while lower concentrations of ZnO NPs showed minimum effect (Figure 4). Similar results were observed by Feris et al. [39] that antioxidative enzyme content increased in Ametryn- and Clomazone-resistant bacteria under HMs. Smaller nanoparticles have also been shown to penetrate in the bacterial cell membrane and inhibit respiratory enzymes (NADH dehydrogenase II), which are a well-known cause of ROS [40]. It was also observed that NPs create holes in the surface of bacteria and generate reactive oxygen species [41]. Furthermore, our results are compatible with previous findings that toxic metals increased ROS in Acinetobacter baumannii, resulting in membrane lipid peroxidation and reducing sugars, proteins, and cell viability. Toxic metal ions impair cell membrane permeability by causing oxidative stress, which inhibits cell development, destroys biomolecules (DNA and protein), and induces cell lysis [29]. As per our results, minimum concentrations of ZnO NPs act as a nutrient for bacteria and reduce oxidative stress. These results are confirmed by earlier findings of Nakano and Asada [42], who discovered that Bacillus spp. cell walls have a (thick) peptidoglycan coating that reduces oxidative stress susceptibility and stabilizes antioxidative enzymes. Bacteria secreting extracellular matrix, such as flagellin-containing (Pyoyanin), reduce NP agglomeration and inactivate toxic ions released by HMs, reducing peroxidase activity [43]. Zinc ions are antioxidative enzyme and are needed for different transporters, zinc-binding proteins, and metallothionine [44]. Bacteria control the electric charges on their surface to repel higher concentrations of NPs. According to the results, nanoparticles with neutral and negative charges have a lower binding affinity for surface bacteria with negative charges, resulting in fewer oxidative stress [45]. Our results are related with the previous findings, as we found that minimum treatment of ZnO NPs has no toxic impact on bacteria and causes lower MDA activity. Zinc ions used by bacteria have been shown to alter the expression of porin genes involved in HM entry as well as the confirmation and fluidity of unsaturated fatty acids by blocking ion passage [46]. According to study, bacteria use zinc ions as a cofactor for a number of enzymes to scavenge hydrogen peroxide [47].
According to scanning electron microscopy (SEM), higher concentrations of ZnO NPs influenced B. cereus morphology, as compared with lower concentrations of ZnO NPs (Figure 6). These findings showed similarity with Naja et al. [48] that when bacterial cells are exposed to higher concentrations of nanoparticles, their morphology changes, resulting in the formation of an irregular cell membrane. ZnO NPs were also found to alter the spiral shape of Campylobacter jajuni into a coccoid form [49, 50]. According to study, a higher concentration of NPs has antimicrobial effects that allow bacteria to alter size and morphology by destroying the cell membrane [51, 52]. ZnO NPs can bind more molecules per unit of surface area by releasing Zn+2 and bind to the phosphate and carboxyl groups of bacteria [53, 54]. It was observed [55, 56] that bacteria use zinc ions as a micronutrient to improve growth and stabilize their membranes by releasing surfactants into the environment by binding with different charges of NPs. Recent findings showed resemblance with the recent report [57, 58] that ZnO NPs remediate the HMs from polluted water.
5. Conclusion
In the present study we have concluded that lower concentrations of ZnO NPs can act as micronutrients for bacterial growth which is subsequently helpful in enhancing the tolerance level of bacteria against HMs by reducing the antioxidative enzyme (SOD, POD, and CAT) and stress biomarker (TBARS and H2O2) levels. Furthermore, from scanning electron microscopy (SEM) data, we could conclude that ZnO NPs interact with bacteria in all different concentrations. However, higher concentrations of NPs severely affected the bacterial morphology and made pores in bacterial cells, while in the case of lower concentrations, the cell remained largely unaffected. It is therefore concluded that lower concentrations of ZnO NPs not only enhance the tolerance potential of plant but also significantly remediate the HM toxicity.
Footnotes
Data Availability
The data used to support the findings of this study are included within the article.
Conflicts of Interest
The authors have no potential conflicts of interest.
Acknowledgments
This study was partially supported by the Higher Education Commission, Pakistan, through research grant no. 20-4372.