
Abstract
On the basis of more than a decade of studies on the cellular effects of ethosuximide, currently, the most prudent view is that together with a block of the low threshold, T-type Ca2+ current, a reduction both of the noninactivating Na+ current, and the Ca2+-activated K+ current in thalamic and cortical neurones contribute to the overall therapeutic action of this antiabsence medicine.
The Original Observation
In 1989, work in thalamocortical (TC) neurons (acutely dissociated and in slices) showed that therapeutically relevant concentrations (250 to 750 μM) of ETX decreased the peak amplitude of the low-threshold T-type Ca2+ current (IT) (2–4). A similar effect was later reported for the IT of neurons in the nucleus reticularis thalami (NRT) (5). This action of ETX was relatively specific (as high voltage-activated Ca2+ currents were unaffected), dose dependent (with a maximal reduction of approximately 32% at 750 μM), and voltage dependent (with a larger reduction for currents evoked between −60 and −40 mV), but there was no effect on the kinetics and steady-state (in)activation properties. ETX was also shown to decrease the Ca2+-activated K+ current [IK(Ca)] in TC neurons (2,3).
The ability of ETX to reduce IT of thalamic neurons soon gained popularity in clinical and experimental reviews and textbooks as one of the main mechanisms of action of the succinimide class of antiabsence medicines. The rationale of this mechanistic link between ETX and IT simply stemmed from the view that a low-threshold Ca2+ spike (LTS), the main voltage expression of T-type channel activation, underlies the action potential firing that is recorded extracellularly from TC neurones during the characteristic EEG spike and SWDs in some experimental models of absence epilepsy (6,7) (discussed later here).
The Contradictory Results
During the same period, evidence against this partial block of IT was accumulating. Different groups were unable to detect any action of therapeutically relevant concentrations (250 to 750 μM) of ETX on IT of nonthalamic cells, including human neocortical neurons (8), three classes of rat hippocampal neurons (9), rat dorsal root ganglion (DRG) cells (10,11), and GH3 pituitary cells (12). In DRG cells, however, a reduction of 91% and 45% of T-type and L-type Ca2+ current, respectively, was observed with 10 μM ETX by Kostyuk et al. (13).
Furthermore, direct attempts to reproduce the block of IT by ETX in TC and NRT neurons from cats and Wistar and Sprague-Dawley rats, as well as from a genetic rat model of absence epilepsy and its nonepileptic control strain, showed either no effect of 0.25 to 1.0 mmol/L ETX (14,15) or a reduction of 19% at 5 mmol/L (16). In one of these studies (15), two additional actions of ETX on TC neurons were observed: (a) a partial reduction of the noninactivating Na+ current (INaP) (60% at 750 μM) (Fig. 1A1), with no effect on the transient Na+ current, and (b) a partial decrease of IK(Ca) (40% at 500 μM) (Fig. 1B1), confirming previous data (2,4).
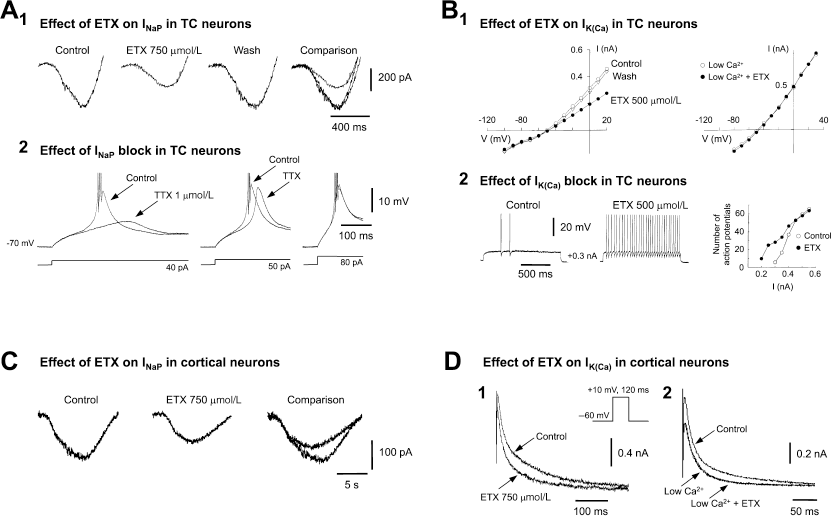
Effect of ethosuximide (ETX) on Na+ and K+ currents of rat thalamocortical (TC) and cortical neurons. (A1.) Whole cell currents elicited by a voltage ramp protocol (from −100 to 50 mV) in a TC neuron show the reversible reduction of the persistent Na+ current (INaP) by 750 μM ETX. A2, LTS, and associated action potential burst firing (top traces) produced by the current steps of increasing amplitude (bottom traces) from a membrane potential of −70 mV, in control conditions and in the presence of 1 μM TTX. Note the smaller and delayed low threshold Ca2+ potential in the presence of 1 μM TTX for the two smallest current steps: this is due to the block by TTX of the non-inactivating Na+ current (INaP) underlying the LTS (see ref. 19 for details). Action potentials are truncated for clarity. (B1.) Steady-state current/voltage plot show the reversible reduction by 500μM ETX of the sustained outward whole cell current (left plot). In another TC neuron recorded in a low Ca2+ (0.5mM) - high Mg2+ (8mM) medium (right plot), 500μM ETX has no effect on the sustained current, indicating that its action is on the Ca2+-activated K+ current (Ik(ca)) component of the sustained outward current. (B2.) Two depolarizing current (0.3nA) pulses show an increased tonic firing in the presence of 500μM ETX. The plot on the right illustrates how this action is restricted to the smallest input currents (0.2–0.4nA). (C.) Whole cell currents elicited by a voltage ramp protocol (from −90 to 50mV) in a layer V cortical pyramidal neuron show the reduction of INaP by 750μM ETX. (D1.) Whole cell currents from another layer V pyramidal neuron show the reduction by 750μM ETX of outward K+ current(s). (D2.) This effect is abolished when ETX is applied in the presence of a low Ca2+ (0.5mM) - high Mg (8mM) medium, indication that ETX is acting on Ik(ca). Panels A and B reproduced with permission from references 15 and 19; (c) 1998 by Society for Neuroscience.
Data from Cloned T-Type Channels
The small effect of ETX (10% at 1 mmol/L and 15% at 3 mmol/L) detected in mouse (17) and human (18) cloned T-type channels, respectively, had been interpreted as indicating that “both α1G and α1H currents are relatively insensitive to ETX, similar to other reports … in DRG and in thalamic neurons” (18). These findings have been enlarged by Gomora et al. (1), who have shown that 1 mmol/L ETX reduces the current generated through human cloned α1G by 5% and 30% when the holding potential is −100 and −75mV, respectively (Fig. 7E in reference 1) (for details, see the previously mentioned commentary). In addition, they observed a preferential reduction (50% at 600 μM) of the steady-state (or “window”) component of the T-type current. Gomora et al.'s interpretation of these data is that they support the notion that native T-type channels are decreased by ETX.
Is the Discrepancy Resolved?
While on the strength of their data one can accept Gomora et al.'s view, the issue remains as to why only one group has so far been able to show a similarly weak block of native thalamic channels by ETX, whereas different groups have failed to do so, particularly as a similar age range, intracellular and extracellular solutions, and types of preparation and thalamic nuclei were used. Gomora et al. (1) suggested that their finding and those of Lacinova et al. (17) provide an explanation to resolve this apparent contradiction, as “studies that reported no effect used very negative holding potentials (–110 mV) where channels are less sensitive to block” (1). This explanation, unfortunately, is not correct, as very negative holding potentials were used in the thalamic studies that did detect an effect of ETX: −100 mV (Fig. 3A and 4 in reference 4), −110 mV (Fig. 1A and 2B in reference 2), −112 mV (Fig. 6B, in reference 3), and −120 mV (Fig. 1A in reference 5).
What is striking, instead, is that the steady-state (in)activation curves of the cloned (human and animal) T-type channels are 15 to 25 mV more depolarized (1,17,18) than those of the native channels (3,4,15). Thus, in contrast to Gomora et al. (1) and Lacinova et al. (17), who tested the ETX block at holding potentials of −60 and −75 mV, respectively, no study on native channels could have correctly used these holding potentials, as even at −75 mV less than 15% of the native current is deinactivated (Fig. 6B in reference 3; Fig. 6b in reference 4; Fig. 3B in reference 15). Although one cannot exclude that a better voltage control is achieved in HEK 293 cells than in acutely dissociated TC neurons, the magnitude of the voltage discrepancy would make this possibility unlikely. Alternatively, the cloned channels might require an auxiliary protein to express similar (in)activation properties as the native channels: In this case, however, it remains to be seen whether cloned channels with more negative (in)activation properties retain a small sensitivity to ETX. Finally, whether caused by the high strength of the charge carrier (5 mmol/L Ca2+ or 10 mmol/L Ba2+), a potential screening effect at the mouth of the channel might have contributed to the block of the recombinant channels by ETX needs further investigation.
Do T-Type Channels Contribute to the Firing of TC and NRT Neurons during SWDs?
In principle, the small block of native IT produced by therapeutically relevant concentrations of ETX would alone affect TC cell excitability by sufficiently decreasing the amplitude of LTSs, thus dampening their ability to evoke action potential bursts (5). A decrease in LTS, however, will also occur because the 50% reduction by ETX of INaP (Fig. 1A1) (15) will remove part of the depolarization required to activate IT, thus resulting in a smaller LTS (Fig. 1A2) (19). As the threshold for activation of IK(Ca) is more depolarized than that of INa>P (15), the reduction by ETX of IK(Ca) (Fig. 1B1) becomes physiologically important in the membrane potential region close to the firing threshold. This resulting increase in tonic firing (Fig. 1B2) would contribute to a disruption of TC loop synchronized activity during SWDs.
As there is no established in vitro model that is capable of reproducing SWDs, ETX has only been shown to decrease hypersynchronous thalamic network activities that rely on the presence of LTSs (5), whereas its ability to block the bicuculline-induced thalamic paroxysms (20) has not been tested. In vivo, however, the majority (60%) of cat TC neurons are completely silent during SWDs (21), and in a rat genetic model of absence, no LTSs are recorded in TC neurons during the majority (90%) of SWDs (22,23). Thus, whether the weak block by ETX of this small T-type component of TC neuron firing during SWDs really represents an important element of its therapeutic action remains questionable. Note that the notion of a small contribution by LTSs to TC neuron firing during SWDs is not in conflict with the recent finding that α1G knockout mice are resistant to pharmacologically induced SWDs (24), as this inability may reflect the concomitant decrease of LTSs in cortical cells.
On the other hand, all NRT neurons in vivo show an LTS (and associated prolonged burst of action potentials) at each cycle of a SWD (25): Thus, the small ETX-induced reduction in the LTSs of NRT neurons (via the small decrease in IT and possibly INaP) would be functionally more important for blocking a paroxysm than an equivalent action in TC neurons. In this respect, it is interesting that ETX evokes a more substantial block of the window component than the peak of IT (1) and that α1I, the most prevalent T-type α subunit in NRT neurons, has the largest window component (26). Furthermore, as IK(Ca) is much larger and functionally more important (for the repolarization that follows an LTS) in NRT than in TC neurons (27), if the ETX block of this current is confirmed in NRT neurons, it is likely that most of the thalamic action of ETX occurs mainly via NRT and not TC neurons. Ultimately, the full therapeutic mechanism of ETX undoubtedly comprises not only these actions on thalamic neurons but also similar effects on cortical cells, which too are endowed with ETX-sensitive INaP and IK(Ca) (Fig. 1C and Fig. D) (28) and possibly IT.