
Abstract
Zebrafish are an important and expanding experimental system for brain research. We describe a noninvasive electrophysiology technique that can be used in living larvae to measure spontaneous activity in the brain and spinal cord simultaneously. This easy-to-use method uses a commercially available multielectrode array to detect local field potential parameters, and allows for relative coordinated (network) measurements of activity. We demonstrate sensitivity of this system by measuring activity in larvae treated with the antiepileptic drug valproic acid. Valproic acid decreased larval movement and startle response, and decreased spontaneous brain activity. Spinal cord activity did not change after treatment, suggesting valproic acid primarily affects brain function. The observed differences in brain activity, but not spinal cord activity, after valproic acid treatment indicates that brain activity differences are not a secondary effect of decreased startle response and movement. We provide a step-by-step protocol for experiments presented that a novice could easily follow. This electrophysiological method will be useful to the zebrafish neuroscience community.
Electrophysiological measurements of the central and peripheral nervous system can be assessed by invasive and noninvasive methods. From a single patched cell to whole brain electroencephalogram (EEG), the neuroscience field has greatly benefited from the numerous techniques available for studies, including seizure propensity, excitatory/inhibitory balance, and pain. Multielectrode arrays (MEAs) are noninvasive systems that offer sensitive measurement of signals, which is widely used for in vitro cultured neurons, cardiac cells, and tissue slices. Microelectrode arrays were largely developed in the 1970s as a way to bridge single-cell properties and population coding in cellular networks. 1 The MEA system also acts as an EEG platform for detection and analysis of transient electrical signals. Discharge, or potentials, are registered as relative voltage changes on the MEA electrodes. The obtained measurements are the superposition of all ionic flow and relative voltage discharge. These potentials are interpreted as local field potentials (LFPs), EEG-like signals, to detect and amplify activity and observe simultaneous activity across multiple electrodes (relative network activity). 2 Measuring LFPs allow for continuous streaming of data from multiple electrodes.
Zebrafish (Danio rerio) are an excellent system for translational studies of the nervous system. With high physiological and genetic homology to humans, the zebrafish field has rapidly developed targeted studies of high-risk neurodevelopmental and mental health genes. 3 From single- to multichannel systems, sophisticated techniques for electrophysiological recordings of the brain in both larvae and adult zebrafish have been previously described.4–17 Some of these electrophysiology systems have used invasive electrodes for recording brain activity,4,6–8,10 or reporter transgenic zebrafish lines.5,11,12 In an investigation from Meyer et al., electrical activity in the larva brain was measured noninvasively over long-term recordings in a perfusion system. 13 These authors employed a “slice anchor” to help immobilize larvae by pressure, but this may cause distress to the animal, and there was no description of anchor construction. Hong et al. designed a microfluidic system for long-term electroencephalographic recordings 14 ; however, the system is not commercially available. Homemade systems, custom-designed electrodes, and systems set up on an electrophysiology apparatus create a barrier for novice laboratories interested in acquiring these methods, and for exploring electrophysiological techniques.4,8,10–12,14–16
We aimed to provide an electrophysiological method accessible to the zebrafish community, that can be easily adapted for laboratories with little to no electrophysiological training. Step-by-step details of the method are described in Supplementary Data. We adapted a MEA technique on a commercially available MEA system, from Axion Biosystems, to measure activity from the nervous system in live zebrafish larvae. In addition, this method measures brain and spinal cord activity simultaneously for useful analysis of the entire central nervous system. Zebrafish larvae were assayed at 7 days postfertilization (dpf), an optimal timepoint for zebrafish behavior and electrophysiology. Larvae are able to live solely on nutrients derived from the yolk until at least 7 dpf, and as long as 14 dpf.
18
This removes potential caveats from differences in feeding. To test sensitivity of live larval electrophysiology, we defined responsiveness to the drug valproic acid as a positive control for diminished brain activity. Valproic acid is a broad-spectrum anticonvulsant that increases turnover of the inhibitory neurotransmitter γ-aminobutyric acid, modulates N-methyl-
We tested the effect of 500 μM valproic acid on 7 dpf larvae locomotor and light-responsive sensorimotor startle behavior. Startle response in zebrafish is a complex behavior that involves sensorimotor gating through several regulatory circuits.21,22 Startle response, which yields kinematic measures, including C-bend angle, distance traveled, and trajectory, can be measured after several different stimuli. 22 Startle response is a relatively conserved mechanism across species and initiates in reticulospinal neurons.23–25 Tactile startle response, induced by touching of the head or tail of a larva, is present at 2 dpf. 26 Acoustic startle response is present after 5 dpf to sound frequencies >200 Hz. 27 Visual startle response develops by 3 dpf. 28 Single-cell resolution of 5–6 dpf zebrafish hindbrain indicated glycinergic neurons are recruited during locomotion, and different GABAergic clusters in the hindbrain and cerebellum are correlated with locomotion after visual stimulus. 29 Additional behavioral paradigms, hardware, and applications are summarized in a recent review by Basnet et al. 30
As described in Supplementary Data, larvae were incubated in E3 embryo medium, with or without addition of valproic acid, for 30 min before testing. Startle response behavior was measured on the DanioVision (Noldus) tracking system, using a noninvasive test where the light source is extinguished for 5 s, at 10-min intervals, for a testing period of 70 min. This method is widely utilized and described by Basnet et al. as a behavioral model for high-throughput screening of neuroactive compounds. 30 Startle activity is observed after each dark stimulus, indicated by increased movement (distance traveled). With addition of valproic acid, we observed a significant decrease in startle response compared with control (Fig. 1A). During the first 10 min of recording, before dark stimulus, we found that larvae treated with valproic acid showed a significant decrease in total movement compared with control (Fig. 1B). This analysis defined a suitable timepoint for electrophysiological testing. Diminished movement of treated larvae persisted over the length of the startle response behavioral testing compared with control (Fig. 1A).
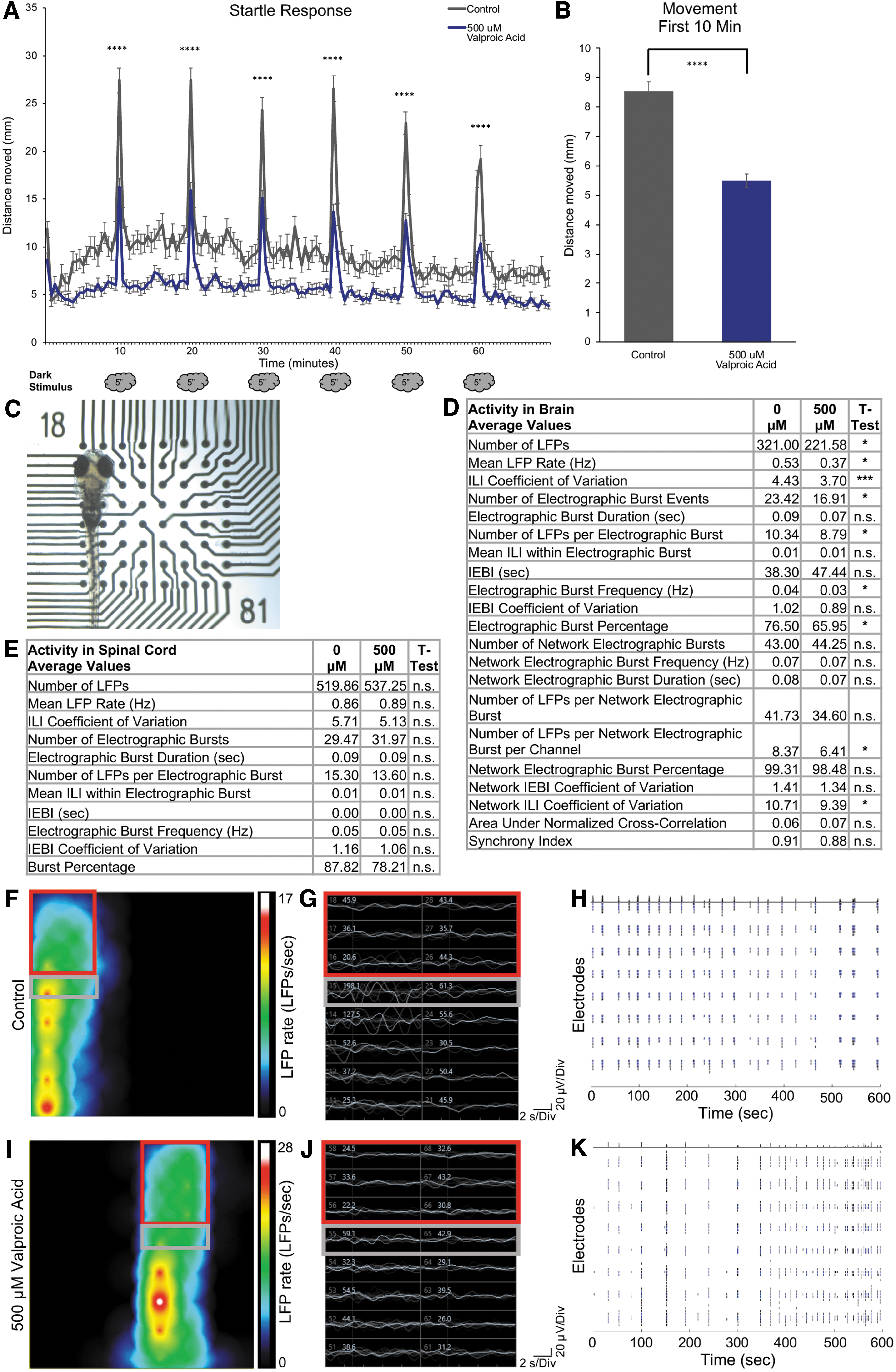
For electrophysiological testing, we compared control and valproic acid-treated 7 dpf larvae. Treated larvae were immersed in precooled 1.5% low-melt agarose in E3 solution and mounted in a 64-electrode six-well CytoView plate (Axion Biosystems) (Fig. 1C, and Supplementary Data). Low-melt agarose for restraining larvae is widely used in electrophysiological recordings.4,8,9,12,14,15 As summarized by Reis et al., EEG artifacts from human brain studies include movement of the head and neck, insufficient electrode contact, cardiac activity, and eye movement. 31 Low-melt agarose will gently restrain the larva to minimize eye and body movement. Niell et al. were able to perform two-photon imaging of tectal neurons after visual stimuli without anesthesia, as the agarose they used was adequate to prevent eye movement. 32 Therefore, we performed the following studies in the absence of a muscle paralytic.
Before solidification of the low melt agarose, larva was positioned with the dorsal side of the head in contact with electrodes. With available electrode configurations, 4–6 electrodes come in contact with the head when placed accordingly. Larval positioning on electrodes is illustrated in Figure 1C. An empty well, coated with agarose, was used as a control for recordings. Once the agarose was solidified, recordings of spontaneous activity were made for a 10-min period on the Maestro Edge (Axion Biosystems). Larva needs to be completely immobilized and in contact with the electrodes as activity is not observed if the larva is moving. We propose that the recordings measured EEG-like signals for the following reasons. Movement artifacts have been found to disrupt EEG-like LFP recordings, supporting that the data we collected are the result of brain and spinal cord activity. 31 Electromyograms (EMGs) measure electrical activity produced by skeletal muscles. The amplitudes of EMG signals are in the millivolt range, whereas the amplitudes of EEG are in the microvolt range.33,34 Therefore, EEG detection requires more sensitive sensors due to the smaller amplitudes of the signals. The Axion Maestro system and software (AxIS) are adequately sensitive to detect and process these smaller EEG-like signals. After recording, we noted which specific electrodes were in contact with the head (brain) or more posterior (spinal cord) region, to assess activity from brain or posterior regions.
AxIS can readily compile the data collected from recordings. However, as the larval recordings are not direct measurements of voltage changes, the data needs to be interpreted as LFPs and electrographic burst events (Supplementary Data). The activity represents a summation of brain and spinal cord regions surrounding an individual electrode, and larva could be specifically placed to measure relative activity surrounding the forebrain, midbrain, and hindbrain regions. Data were collected for each region, analyzing LFPs (firing frequency in Hz), electrographic burst events (minimum 5 LFPs/100 ms) and relative network activity (minimum 3 LFPs detected simultaneously between a minimum of two electrodes). LFP detection was filtered at 6 × standard deviation to remove potential artifacts. Methods are expanded in the Supplementary Data.
We observed a significant decrease in the average number of LFPs, mean LFP rate, and inter-LFP interval (ILI) coefficient of variation (that indicates regularity of LFPs), after treatment with 500 μM valproic acid compared with control (Fig. 1D). The average number, frequency, and percentage of electrographic burst events, along with LFPs per electrographic burst, were significantly decreased after treatment. No difference was observed in electrographic burst duration or in inter-electrographic burst events properties, indicating valproic acid decreased LFP propensity but not overt LFP properties (Fig. 1D). This coincides with reported valproic acid action on glutamatergic excitatory neurons. 19 We were additionally able to measure relative network activity across the brain. Overall, we did not observe significant changes to network activity after treatment, but there was a significant decrease in number of LFPs per network electrographic burst/channel, and network ILI coefficient of variation, indicating decreased detection of LFPs across the brain yet with less variability (Fig. 1D). A representative heat map illustrates a snapshot of LFP rate and differences between head and body activity between control and treated larva (Figs. 1F, 1I). Representative traces of LFP amplitude (white text) and raster plot (total LFP activity over time) illustrate differences in overall activity in treatment compared with controls (Fig. One gram, H, J, K). We did not detect any signal in the millivolt range, indicating further that we were not measuring EMGs. Conversely, all signals detected were in the microvolt range, consistent with EEG signals (Fig. 1G, J). Collectively, the overall decrease in LFPs after treatment with valproic acid aligned to the decreased seizure and field potential data collected in Hong et al. 14
Another advantage of this MEA system is simultaneous measurement of brain and spinal cord activity. Data were analyzed and pooled for electrodes in contact with the spinal cord region anterior to the swim bladder (Fig. 1C, E–J). Significant differences in activity were not observed between valproic acid treatment; however, we noted overall activity is substantially higher compared with head measurement activity. Furthermore, the observation of diminished brain activity, but not spinal cord activity, after treatment with valproic acid supports brain activity differences are not a secondary effect from decreased startle response and movement. From these findings, we propose that muscle paralytics are not essential to acquire LPFs from live zebrafish larvae under these experimental parameters.
For these studies, we utilized the Noldus DanioVision for larvae tracking and the Axion Maestro Edge for zebrafish LFP measurements. The DanioVision is an attractive system widely used for high-throughput single and multiple body point tracking. The EthoVision XT software easily compiles tracking data for analysis. For electrophysiological studies, users need to be proficient in electrophysiological techniques to set up recordings and analyze data properly. We present the Axion Maestro system for live whole animal zebrafish measurements regardless of electrophysiology background, with options to add on hardware such the Axion Lumos multiwell optical stimulation system. In addition, we tested this system on 14 dpf larvae and observed increased overall activity with maturation of the nervous system (data not shown). However, we recognize that the six-well CytoView plate only allows for between 4 and 6 electrodes to come into contact with the 7 dpf larval head. A more suitable, smaller electrode layout with a higher density of electrodes in a given area would be excellent to measure specific brain regions. Laboratories without the Noldus and Maestro Edge can adapt these experiments on similar systems. Alternative cost-effective tracking and electrophysiological systems can be assembled according to Selvaraj and Santhakumar (2017) 35 and Garma et al. 17
Zebrafish have multiple outstanding attributes for studies related to human neurological and neurodevelopmental disorders, including conservation of genes with human, and disease-associated phenotypes. The ability to readily perform electrophysiology allows measurements of phenotypes that include seizure propensity after chemical induction. 36 Using the apparatus, electrodes can also be used to induce activity, allowing analysis of neuronal circuits after stimulation. 37 The ability to record brain and spinal cord LFP activity in live larvae with a commercially available system allows for accessible electrophysiological studies for all zebrafish investigators.
Footnotes
Acknowledgments
The authors thank Olivier Paugois for excellent fish care. They thank Axion Biosystems, specifically Carina Peritore and Stacie Chvatal, for help in modifying parameters to best suit the zebrafish system.
Disclosure Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
This work was supported by a grant from the Simons Foundation to the Simons Center for the Social Brain at MIT, National Institute of Mental Health (NIMH) of the National Institutes of Health (1R01MH119173-01A1), Balkin-Weinberg-Markell Fellowship, and additional support from Jim and Pat Poitras.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.