
Abstract
Immunoglobulin (Ig) G4-related disease (IgG4-RD) is a newly recognized systemic fibroinflammatory disease with characteristic histological findings and high serum IgG4 levels. Epstein–Barr virus (EBV) is a persistent herpesvirus in B lymphocytes, and we previously reported EBV reactivation-induced Ig production. We showed that EBV reactivation induced the production of thyrotropin receptor antibodies, the causative antibodies of Graves' disease. In the present study, we investigated whether EBV reactivation induced IgG4 production and if EBV-positive B cells or IgG4-positive plasma cells are present in the thyroid tissues of Graves' disease patients with lymphoplasmacytic infiltration. EBV-encoded small RNA1 (EBER1) in situ hybridization and immunohistochemistry for IgG and IgG4 were performed on seven resected thyroid tissues with lymphoplasmacytic infiltration collected from the thyroids of 11 Graves' disease patients. We then cultured the lymphocytes of 13 Graves' disease patients and 14 controls and induced EBV reactivation to measure IgG4 levels in culture fluids. We detected EBER1-positive cells and IgG4-positive plasma cells in the same area of thyroid tissues. EBV-reactivated cells with IgG4 on their surface were observed in culture cells, and IgG4 production was detected in culture fluids. The IgG4/IgG percentage was higher than that in normal serum level. A subset of Graves' disease is an IgG4-RD-like condition, not an IgG4-RD. EBV reactivation stimulates IgG4 production, which may result in high serum IgG4 levels and promote IgG4-positive plasma cell infiltration. EBER1 needs to be examined when an increase in IgG4-positive plasma cell numbers is noted.
Introduction
I
IgG4 is a low-affinity antibody (35) that weakly binds to Fcγ receptors (2) and does not bind to complement 1q (5). The location and mechanisms by which IgG4 is produced have not yet been elucidated. It also remains unclear whether IgG4 contributes to the pathogenesis of IgG4-RD or is only produced as a consequence of the disease. Furthermore, the involvement of autoimmune mechanisms in the development of IgG4-RD is still debatable (5,13,29).
Epstein–Barr virus (EBV) is a common human herpesvirus that latently infects most adults (16). EBV stably infects, but sometimes changes its mode of infection from latent to lytic, and this is referred to as reactivation (3,11). EBV mainly infects B lymphocytes and thus is speculated to affect the antibody production by host B cells. Various serum autoantibodies have been observed in the acute phase of infectious mononucleosis (8,21), which suggests a relationship between EBV and autoimmunity.
Graves' disease was reported to show high serum IgG4 in some cases (31). We previously demonstrated the presence of EBV-infected B cells with a thyrotropin receptor antibody (TRAb) as the surface globulin [TRAb(+)EBV(+) cells] in peripheral blood mononuclear cells (PBMCs) from Graves' disease patients and controls (19). TRAbs are causative autoantibodies of Graves' disease. We induced EBV reactivation on PBMCs containing TRAb(+)EBV(+) cells and observed the production of IgG, IgM, IgE, and TRAb as well as the expression of activation-induced cytidine deaminase (AID) and its mRNA (AICDA) (Fig. 1) (20,22). Our findings suggested that EBV reactivation stimulates plasma cell differentiation of host B cells and Ig production. Therefore, we proposed an EBV reactivation-induced Ig production mechanism (Fig. 1): (1) EBV infection and polyclonal B cell activation, (2) AID expression and class-switch recombination (CSR), and (3) EBV reactivation, plasma cell differentiation, and Ig production. The Igs produced may be low in affinity for antigens, but some show an isotype change from the IgM on naive B cells.
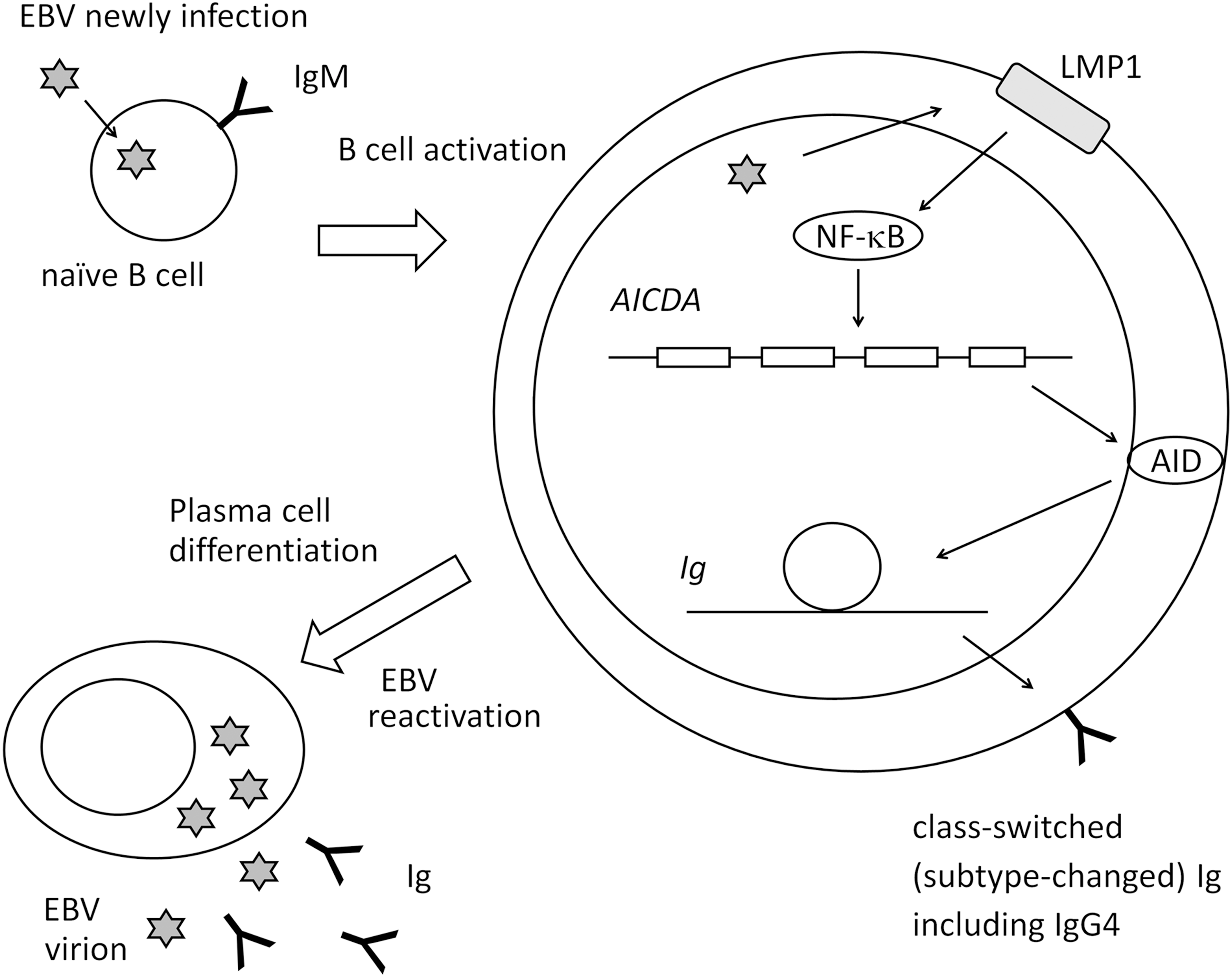
Pathway for EBV reactivation-induced Ig production. B cells newly infected by EBV become latency 3 and express LMP1. LMP1 activates host cells (polyclonal B cell activation). LMP1 stimulates AID expression through NF-κB, which enables host cells to class-switch Ig. EBV reactivation induces plasma cell differentiation and Ig production. AID, activation-induced cytidine deaminase; EBV, Epstein–Barr virus; Ig, immunoglobulin; NF-κB, nuclear factor-kappa B.
We herein examined the presence of IgG4-positive plasma cells and EBV-infected cells in resected thyroid tissue of Graves' disease and investigated whether IgG4 is produced by an EBV reactivation-induced Ig production mechanism in Graves' disease patients and controls.
Materials and Methods
Subjects
Twenty-four Graves' disease patients and 14 healthy controls participated in the present study (Tables 1 and 2). All subjects provided written informed consent for participation in the study, and the study protocol was approved by the Medical Ethics Committee for Human Subject Research (No. 707, 707-1-13) at the Faculty of Medicine, Tottori University, Yonago, Japan. Eleven patients had also participated in our previous study (22).
AICDA, mRNA of activation-induced cytidine deaminase (AID); NT, not tested; TRAb, thyrotropin receptor antibody.
Each plasma cell count is an average of three corresponding fields for IgG4 and IgG.
EBER1, Epstein–Barr virus-encoded small RNA 1; ISH, in situ hybridization; lymphocyte infiltration (+), hyperplastic thyroid with moderate lymphoplasmacytic infiltration; lymphocyte infiltration (+/−), hyperplastic thyroid with no or few lymphoplasmacytic infiltration; TRAb, thyrotropin receptor antibody; HPF, high power field; NT, not tested.
Eleven of the 24 patients underwent thyroidectomy. Nine of the 13 patients without surgery were receiving treatments with antithyroid drugs (methylmercaptoimidazole or propylthiouracil). Control subjects voluntarily enrolled. Their thyroid functions were normal, and they had no family history of thyroid disease.
The mean ages (±SD) of Graves' disease patients treated with thyroidectomy (11 cases; Table 2), those treated with antithyroid drugs (nonsurgical group) (13 cases; Table 1), and healthy controls (14 cases; Table 1) were 42.6 ± 15.4, 41.5 ± 8.6, and 35.9 ± 11.6 years, respectively.
Sampling protocol
PBMCs were cultured for 2 days in RPMI1640/10% FBS with 0.1 μg/mL cyclosporin A at 37°C, as described previously (4,19,20), to suppress T cell function and enrich the B cell population. They were then transferred to a culture at 33°C to induce EBV reactivation and were regarded as day 0 samples. A culture at 33°C has been shown to physiologically induce EBV reactivation (9,20,26,27).
On days 0, 5, 10, and 12, half of the culture fluid was sampled and replaced with fresh medium (Fig. 2). Culture cells were collected on days 0 and 5 for flowcytometry (FCM) and confocal laser microscopy.
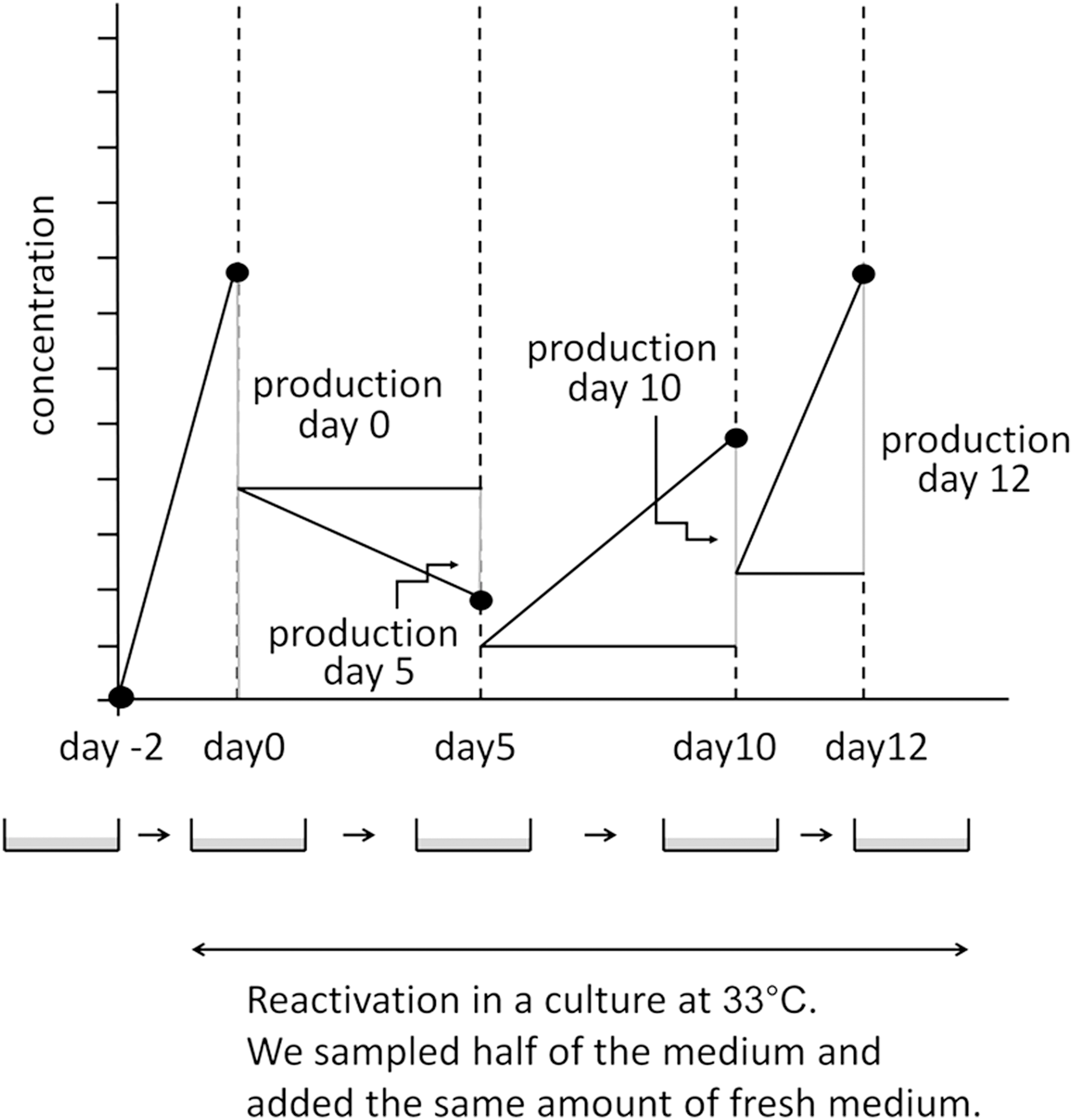
Sampling protocol. We cultured PBMCs at 33°C to induce EBV reactivation. On days 0, 5, 10, and 12, we collected half of the medium for examination and added the same amount of the fresh medium. Ig production at each sampling point was assessed by subtracting half of the values of the previous point from those at the sampling point. Adapted from Nagata et al. (20). PBMCs, peripheral blood mononuclear cells.
In the 11 common subjects of our previous and present studies, the culture fluids used in this study were the same aliquots as those in the previous study.
Ig measurements in culture medium and serum
We measured IgG4 concentrations in the culture medium using an ELISA kit (Affymetrix eBioscience, Vienna, Austria) and IgG concentrations using ELISA (Bethyl No. E80-104, E80-100, E80-108; Montgomery, TX) according to the manufacturer's instructions. On the days when the culture medium was sampled, we collected half of the medium for examinations and added the same amount of the fresh medium (Fig. 2). Ig production at each sampling point was assessed by subtracting half of the values of the previous point from those at the sampling point.
FCM and confocal laser microscopy
The culture cells collected were fixed by 2% paraformaldehyde and fluorostained by a 72A1 mouse monoclonal antibody (Millipore, Temecula, CA) to detect glycoprotein 350/220 (gp350/220), which is expressed in the late phase of EBV reactivation. Since gp350/220 is also expressed on the host cell surface in this phase (26,34), we used it for FCM (Supplementary Fig. S1; Supplementary Data are available online at
Cells were incubated with 3 μL/106 cells of 72A1 at 4°C for 30 min. After the incubation, they were washed and incubated with 1 μg/106 cells of goat anti-mouse IgG H&L (Alexa Fluor® 488) (Abcam, Cambridge, United Kingdom) as a secondary antibody.
We used 1 μg/106 cells of an anti-human IgG4 rabbit monoclonal antibody, clone RM120 (Abnova, Taipei, Taiwan) to stain surface IgG4, and 1 μg/106 cells of goat anti-rabbit IgG (H + L) (Alexa Fluor® 647) (Invitrogen, Carlsbad, CA) as a secondary antibody.
Fluorostained samples were analyzed and sorted with the cell sorter, MoFlo XDP (Beckman Coulter, Fullerton, CA).
IgG4(+)72A1(+) cells were sorted and examined using the confocal laser microscope, TCS SP-2 (Leica, Wetzlar, Germany).
In situ hybridization and immunohistochemistry
Seven of the 11 cases underwent thyroidectomy showed focal moderate lymphoplasmacytic infiltration in the hyperplastic thyroid gland. In situ hybridization and immunohistochemistry were performed on these seven cases only.
The in situ hybridization of EBV-encoded small RNA1 (EBER1) was performed with EBER1-PNA Probe/Fluorescein (Y5200; Dako, Glostrup, Denmark), and immunohistochemistry for IgG4 and IgG was conducted with the anti-human IgG4 rabbit monoclonal IgG, clone RM120 (Abnova), and anti-human IgG mouse monoclonal IgMκ (A57H) (Nichirei, Tokyo, Japan). Envision Dual Link System-HRP (K4063) (Dako) was used for a secondary antibody.
Statistical analyses
Statistical analyses were performed using SPSS Statistics 21 (IBM, Armonk, NY).
Friedman's test was adopted to analyze time course variations in Ig concentrations. The Mann–Whitney U test was used to compare IgG4 percentage (IgG4/total IgG%) peaks.
Results
IgG4 production was stimulated by EBV reactivation
We added three controls and four patients to the common subjects and obtained the same significant change of total IgG (Friedman's analysis: p = 0.000 for controls and p = 0.003 for patients) as previous report (Fig. 3A).
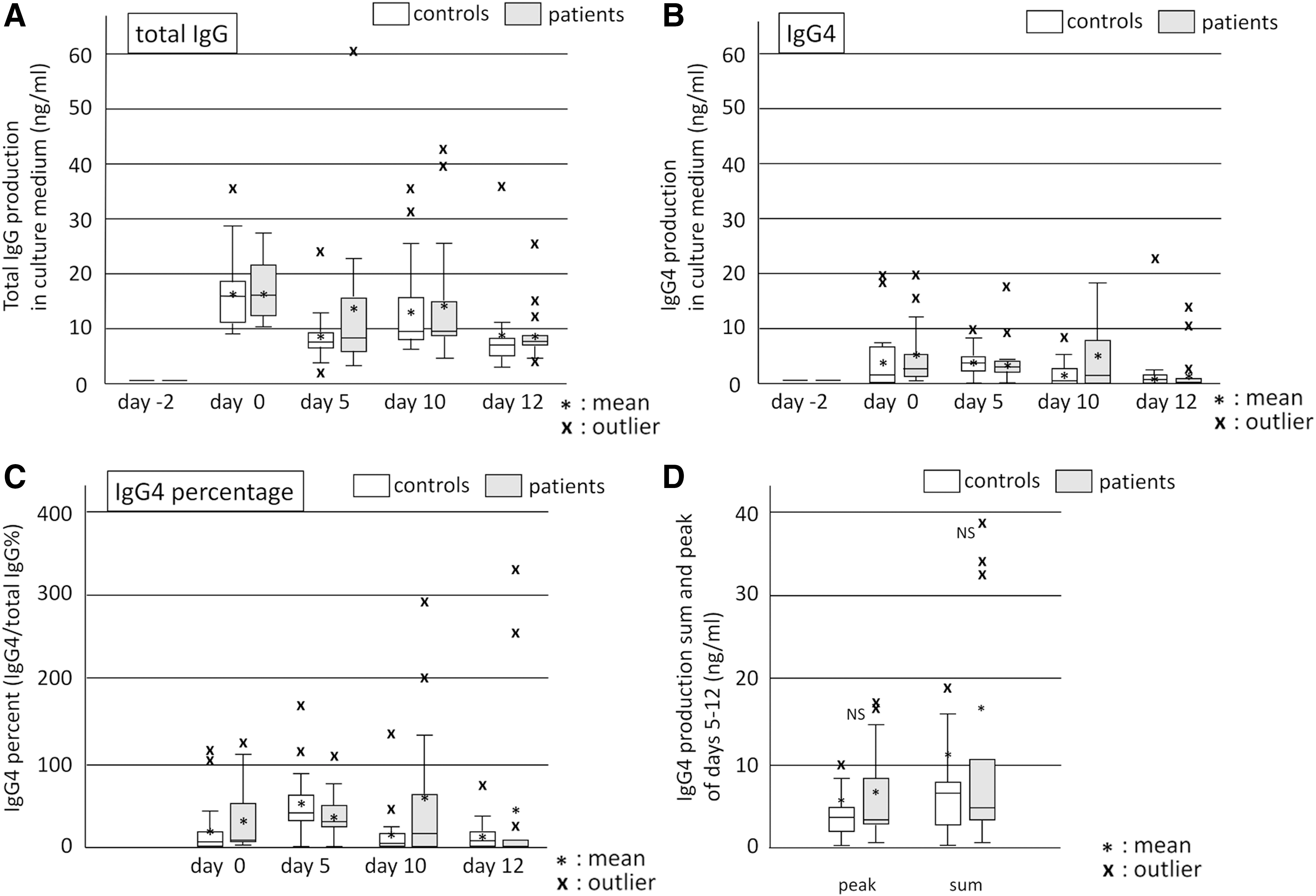
Production of IgG and IgG4 during the EBV reactivation period. Culture fluids were sampled on days 0, 5, 10, and 12, and IgG
IgG4 levels significantly changed during the sampling period (Friedman's analysis: p = 0.005 for controls and p = 0.007 for patients) with the first peak being observed on day 0 (Fig. 3B). Since the induction of EBV reactivation started at day 0, PBMCs on day 0 did not receive stimulation for reactivation. We confirmed that these PBMCs on day 0 contain 4–12% of CD38(+) cells or CD138(+) cells in our previous study (22). Therefore, the day 0 peak represented the production from plasma cells or activated B cells that were already present, and the production stimulated by EBV reactivation was that with peaks on days 5, 10, and 12. Patients and controls showed production with peaks on day 5 or 10, indicating IgG4 production stimulated by EBV reactivation.
IgG, IgM, and IgG4 levels on day-2 (start point) were the same as those of the negative control (0 ng/mL of ELISA standard); therefore, we disregarded the effects of Igs originally in the culture medium.
Culture cells on day 5 contained IgG4(+)72A1(+) double-positive cells
There were 9.88% of 72A1(+) cells in day 5 cells (mean of 4 samples), suggesting that cells on day 5 were affected by the induction of reactivation (Supplementary Table S1).
We noted red spots of IgG4 on IgG4(+)72A1(+) double-positive cell surfaces and fine green dots on cell surfaces and in the cytoplasm that represented the 72A1 antibody binding to EBV-gp350/220 (Fig. 4 and Supplementary Fig. S1).
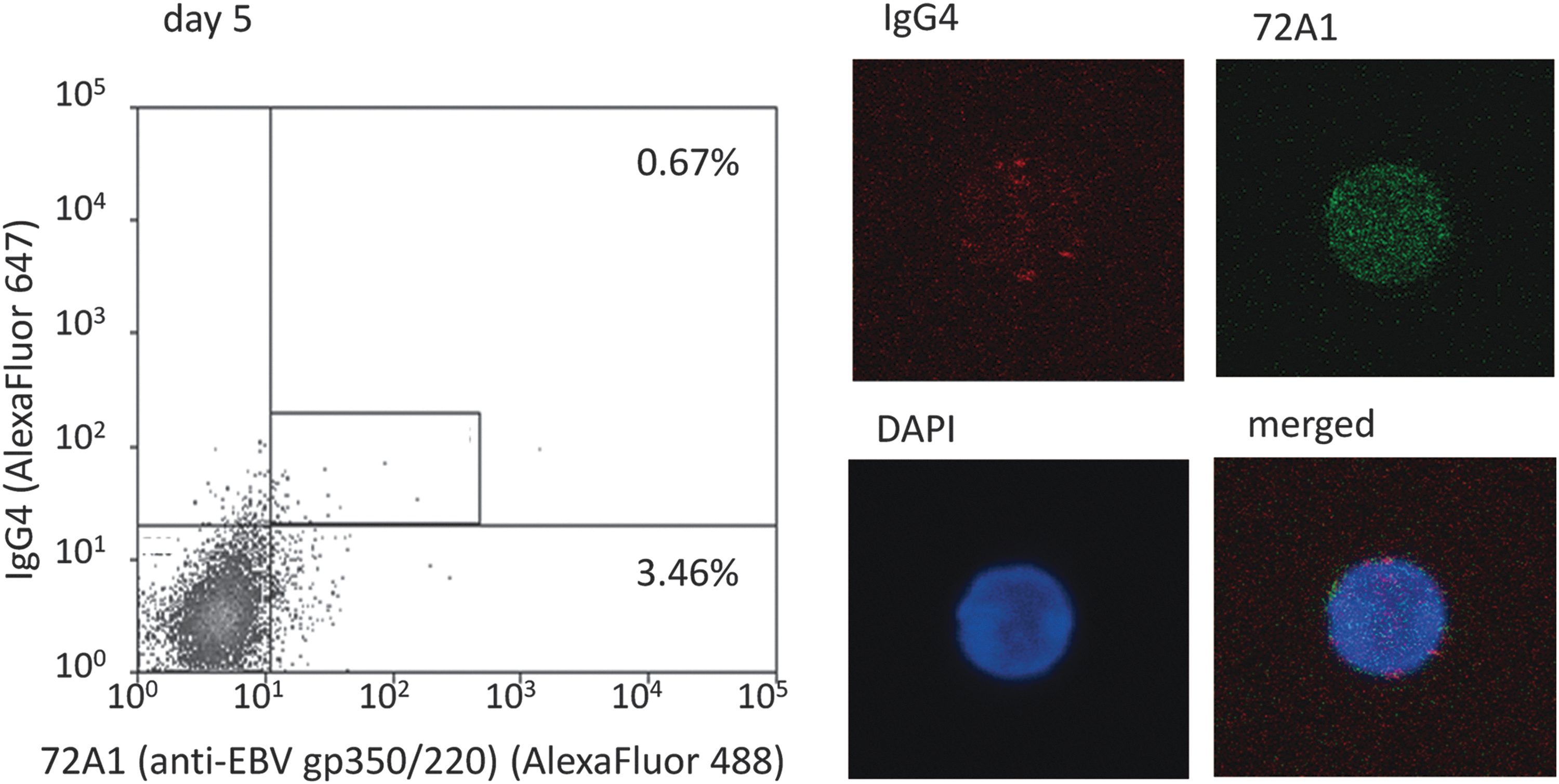
Detection of IgG4(+)72A1(+) double-positive cells in culture cells on day 5. We detected IgG4-positive and EBV-reactivated [IgG4(+)72A1(+)] cells in culture cells on day 5 and confirmed sorted cells by confocal laser microscope. Red spots are surface IgG4 and fine green dots are 72A1 antibodies binding to EBV-gp350/220. The frequency of 72A1(+) cells of this sample (representative case; patient 3) was 4.13% (0.67 + 3.46), and the mean frequency of four samples was 9.88% (Supplementary Table S1).
IgG4 percentage (IgG4/IgG) in culture fluids increased during EBV reactivation
On day 0 (before reactivation), mean IgG4 percentages in patients and controls were ∼35.88% and ∼23.99%, respectively. During reactivation, they increased to ∼60.59% in patients (day 10) and ∼56.19% in controls (day 5) (Fig. 3C).
The normal value of serum IgG4/IgG has been reported to be approximately 4% (25). IgG4 production induced by EBV reactivation resulted in the greater percentage than serum levels.
Comparison of IgG4 production between patients and controls
We compared IgG4 production in patients and controls by peaks and sums during reactivation. They were slightly higher in patients than in controls, but they were not significant (Fig. 3D).
EBER1-positive cells and IgG4-positive cells were observed in the same areas
The pathological findings of resected tissues were the diffuse hyperplasia of thyroid glands in all 11 cases, with 7 of the 11 cases showing the focal moderate infiltration of lymphoplasmacytic cells, and most cases also had lymphoid follicles. However, we did not detect tumefactive lesions, storiform fibrosis, or obliterative phlebitis. The immunohistochemistry of seven cases with moderate lymphoid cell infiltration revealed that EBER1-positive cells and IgG4-positive plasma cells were present in the same area with lymphoid cell infiltration (Fig. 5; Table 2). Six of the seven cases had a large number of IgG4-positive plasma cells (over 10 cells in a high power field [10 < HPF <]) and the ratio of IgG4(+) cells/IgG(+) cells was greater than 40%.
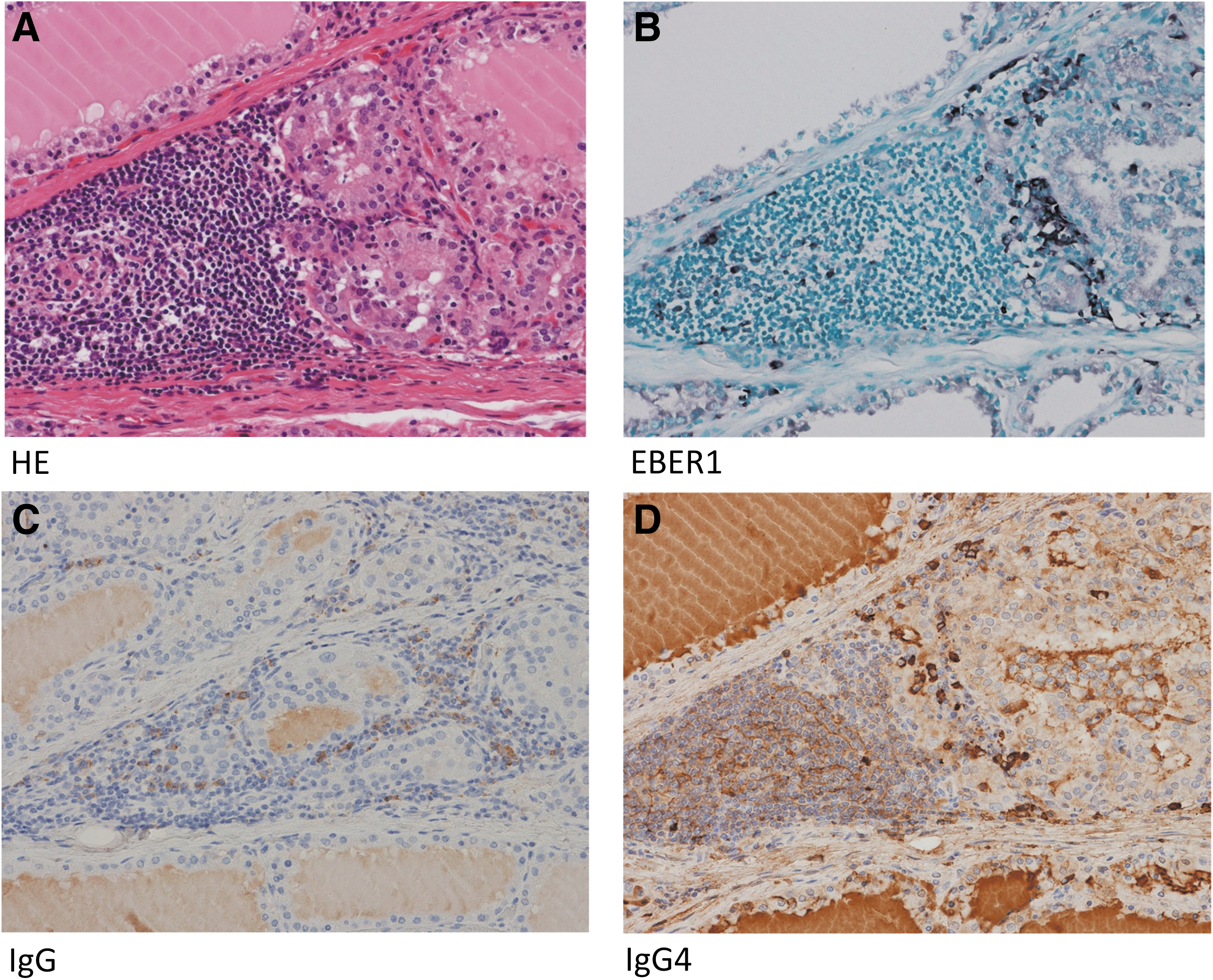
EBER1-positive cells and IgG4-positive cells were observed in the same areas. Resected tissues showed the diffuse hyperplasia of thyroid follicular epithelial cells with the focal infiltration of lymphocytes, but not tumefactive lesions, storiform fibrosis, or obliterative phlebitis
Discussion
Eleven of the 24 Graves' disease patients underwent thyroidectomy due to difficulties associated with controlling their thyroid function with antithyroid drugs. Eleven resected specimens exhibited diffuse hyperplasia, and 7 of the 11 cases had the focal moderate infiltration of lymphocytes with lymphoid follicle formation (Fig. 5A). This lymphoid cellular infiltration contained a similar number of IgG4-positive plasma cells to that of IgG4-RD (Fig. 5D; Table 2). IgG4 serum levels in our patients who underwent thyroidectomy were high, particularly in three patients with levels higher than 135 mg/dL, which were also similar to IgG4-RD (Table 2). However, we did not detect tumefactive lesions, storiform fibrosis, or obliterative phlebitis. Therefore, Graves' disease is not an IgG4-RD, but a subset of Graves' disease is an IgG4-RD-like condition.
This is consistent with previous findings showing that some patients with Graves' disease (31) show high serum IgG4 levels. Most of our cases of thyroidectomy had very high levels of TRAbs as well as IgG4.
During the induction of EBV reactivation on PBMCs from patients and controls, we observed EBV-reactivated cells with IgG4 on their surface [IgG4(+)72A1(+) cells]. In the culture fluid of PBMCs containing IgG4(+)72A1(+) cells, we detected IgG4 in addition to IgG, IgM, and IgE (Table 1; Fig. 3) [previous report: (22)], which indicated that IgG4 was also produced by an EBV reactivation-induced Ig production, the alternative system of antibody production. AID, necessary for CSR, is supplied during EBV reactivation because EBV newly infected B cell can be activated and express EBV-LMP1 that promotes the expression of AICDA mRNA through nuclear factor-kappa B (NF-κB) (7,18,22,36) (Fig. 1). The CSR to IgG4 (Supplementary Fig. S2) requires TH2 cytokines and IL-10 from regulatory T cells (Treg) (1,24). The EBV-BCRF1 gene product exhibits high homology to IL-10 (8), and EBERs stimulate the expression of human IL-10 from host B cells (28). Viral and human IL-10 may be attributed to IgG4 production instead of Treg cells.
In contrast to the small percentage of normal serum IgG4 in total IgG (approximately 4%) (25), the mean values of IgG4/IgG in the culture fluid during the observation period were more than 15% (Fig. 3C). Several cases showed that most of the IgG produced was IgG4, accounting for differences in the sensitivities of the antibodies used in IgG4 ELISA and IgG ELISA. Our results suggest that EBV reactivation-induced Ig production generates a higher percentage of IgG4 than typical Ig production.
In typical Ig production, plasma cells produce high-affinity IgG antibodies (17,25), but IgG4 is not a high-affinity antibody (2,33,35). Therefore, it may only rarely be selected in affinity maturation. IgG4-positive plasma cells that infiltrate inflammatory tissues may have differentiated through another process, not the germinal center reaction.
Our results showed that EBV reactivation stimulates IgG4 production in a subset of Graves' disease, and in some cases, it may be the main source of IgG4. Therefore, high serum IgG4 levels and the infiltration of IgG4-positive plasma cells observed in IgG4-RD-like conditions may be present as a consequence of chronic inflammation and EBV reactivation.
Immunohistochemistry and in situ hybridization revealed that EBER1-positive cells were present in the thyroid glands of Graves' disease patients with lymphocyte infiltration (Fig. 5B; Table 2). We detected IgG4-positive plasma cells at more than 10 cells/HPF in the same area (Fig. 5D; Table 2). These results are consistent with previous reports showing that the number of EBV-positive cells increased in IgG4-related lymphadenopathy (30) and that a case of Graves' disease with severe hypothyroidism had characteristics of IgG4-RD (23).
Histologically, there are two types of Graves' autoimmune hyperthyroidism, the one with lymphocytic infiltration and the other with no significant inflammatory infiltration. In this study, we noticed EBER1-positive cells and IgG4-positive plasma cells in the same area with lymphocyte infiltration. Besides, we did not find any tumors or inflammatory diseases except for thyroid gland in all our subjects clinically. Therefore, it is one interpretation that IgG4 may be induced by EBV reactivation in Graves' disease with lymphocytic infiltration as a thyroid-specific IgG4 thyroiditis (10,14,15,23). However, as we showed EBV reactivation-induced IgG4 production in cultured PBMCs, IgG4 production may occur in circulation as well as in local thyroid tissue with lymphoplasmacytic infiltration in a chronic inflammatory condition with EBV reactivation.
Therefore, EBV may also contribute to the mechanisms underlying IgG4 production, even in systemic IgG4-RD (12,32); however, further investigations are required. We need to examine EBER1 in situ hybridization to confirm the infiltration of EBV-infected B cells when an increase in IgG4-positive plasma cells is noted.
Footnotes
Acknowledgments
We are grateful to Dr. Shuji Fukata (Kuma Hospital) and former professor Takeshi Sairenji (Tottori University). We also thank the Medical English Service (Kyoto, Japan) for proofreading this article. This work was supported by the Tottori University Faculty of Medicine Research Grant (2015), Discretionary funds of the director of Tottori University Hospital (2016), and JSPS KAKENHI Grant No. 17K08694 (K.N.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.