
Abstract
In April 2021, a plague outbreak was identified within one Marmota himalayana family shortly after emerging from hibernation, during plague surveillance in the M. himalayana plague foci of the Qinghai-Tibet Plateau. A total of five marmots were found dead of Yersinia pestis near the same burrow; one live marmot was positive of Y. pestis fraction 1 (F1) antibody. Comparative genome analysis shows that few single nucleotide polymorphisms were detected among the nine strains, indicating the same origin of the outbreak. The survived marmot shows a high titer of F1 antibody, higher than the mean titer of all marmots during the 2021 monitoring period (W = 391.00, Z = 2.81, p < 0.01). Marmots live with Y. pestis during hibernation when the pathogen is inhibited by hypothermia. But they wake up during or just after hibernation with body temperature rising to 37°C, when Y. pestis goes through optimal growth temperature, increases virulence, and causes death in marmots. A previous report has shown human plague cases caused by excavating marmots during winter; combined, this study shows the high risk of hibernation marmot carrying Y. pestis. This analysis provides new insights into the transmission of the highly virulent Y. pestis in M. himalayana plague foci and drives further effort upon plague control during hibernation.
Introduction
Plague is a zoonotic disease caused by the Gram-negative bacterium Yersinia pestis, with pneumonic plague, bubonic plague, and septic plague as the primary plague syndromes (Yang 2018). Cases of plague in humans often occur after being bitten by an infected flea. The clinical manifestations are mostly bubonic plague, accompanied by fever and local lymphadenopathy, while a few cases of bubonic plague may develop into secondary pneumonic plague through sepsis (Glatter and Finkelman 2021). Unlike bubonic plague, pneumonic plague can spread from person to person and readily cause plague outbreaks.
Although the early use of effective antimicrobial therapy can markedly reduce the risk of death from the plague, case fatality rates have remained high, at ∼10% and 40% in previous bubonic plague and pneumonic plague epidemics between 2010 and 2015, respectively, as reported by the World Health Organization (WHO) (Anonymous 2016). In addition, the natural foci of plague are widely distributed in Asia, Africa, and the Americas. In recent years, the reported cases may pose a potential risk of population transmission (Forrester et al. 2017, Andrianaivoarimanana et al. 2019, Zhou and Guo 2020).
Marmota himalayana, the main natural host of plague, is widely found in the Plague foci of M. himalayana in Qinghai–Tibet Plateau, especially along the Qilian Mountains and Altun Mountains in Gansu Province (Shuli 1988, He et al. 2021). Fleas are important vectors of Y. pestis between the host (wild animals or synanthropic rodents) and humans (Chouikha and Hinnebusch 2012). Oropsylla silantiewi and Callopsylla dolabris are the main vector insects in this focus. The marmots live in alpine scrub and alpine meadow at the altitude of 2700–5450 meters and go in to hibernate from September-October till the following end of March-early April each year. The frequent epidemics of plague among marmots in this area pose a significant risk to local herders (Ge et al. 2015).
A human plague pandemic is unlikely to be triggered because the area is sparsely populated. However, the possibility of plague spreading to densely populated areas through tourism and road development cannot be excluded. Employed shepherds are primary victims of the plague, some of whom even dig hibernating marmots out during their hibernation period despite the risk of infection. Human plague case caused by excavating, skinning, and eating hibernating M. himalayana has been reported previously, but Y. pestis was not isolated due to no marmot remains. This study provides direct evidence of hibernating marmots carrying Y. pestis.
During surveillance of the marmot plague foci of Akesai Valley in the Altun Mountains, a plague outbreak was discovered among an M. himalayana family in April 2021. The epidemiology, etiology, and serological characteristics of this outbreak are described herein, which will facilitate understanding of plague epidemics in the M. himalayana plague foci of the Qinghai-Tibet Plateau. Combined with the hypothesis that Y. pestis lurks in hibernating animals (Bizanov and Dobrokhotova2007), a new understanding is provided here on the preservation and transmission of Y. pestis in hosts and vectors during hibernation and on the plague epizootics after awakening in the spring. This study reflects the practical significance of the mechanism of Y. pestis circulation in natural plague foci and the potential for human infection from hibernating rodents or their fleas. When rising to 37°C, Y. pestis will experience the temperature of flea (25–28°C) and rodents (37–40°C) in sequential order. Y. pestis fast proliferates at flea temperature (Brenner et al. 2005) and initiates low-calcium response (LCR) and virulence expression at rodent temperature.
Materials and Methods
Marmota himalayana plague foci in the Qilian and Altun Mountains
Yersinia pestis was isolated from M. himalayana in Xiahe County in 1959, which confirmed the existence of natural plague foci in Gansu Province for the first time. To date, two independent marmot plague foci have been discovered in the Qilian Mountains-Altun Mountains and the Gannan Plateau, respectively. The region of the Qilian Mountains and Altun Mountains is composed of parallel mountains, with altitudes of 3000–4500 meters. The plague foci surveyed in this study have an alpine and semiarid climate, with an annual average temperature of less than 4℃, annual precipitation of 100–500 millimeters, and good vegetation for natural pasture.
Specimen source and strain isolation
In April 2021, five marmots (namely No. 4, No. 5, No. 6, No. 7, and No. 14) were found dead in the environment, and one marmot (No. 16) was captured in the vicinity of the same burrow at an altitude of about 3200 meters in the Akesai Valley (Fig. 1 and Fig. 2), during plague surveillance of the marmot plague foci in the Altun Mountains in Gansu Province. The marmots found dead in the environment referred to the marmot that had been found dead at the time of sampling, and usually referred to the dead marmot that died of disease or unknown cause in the field.
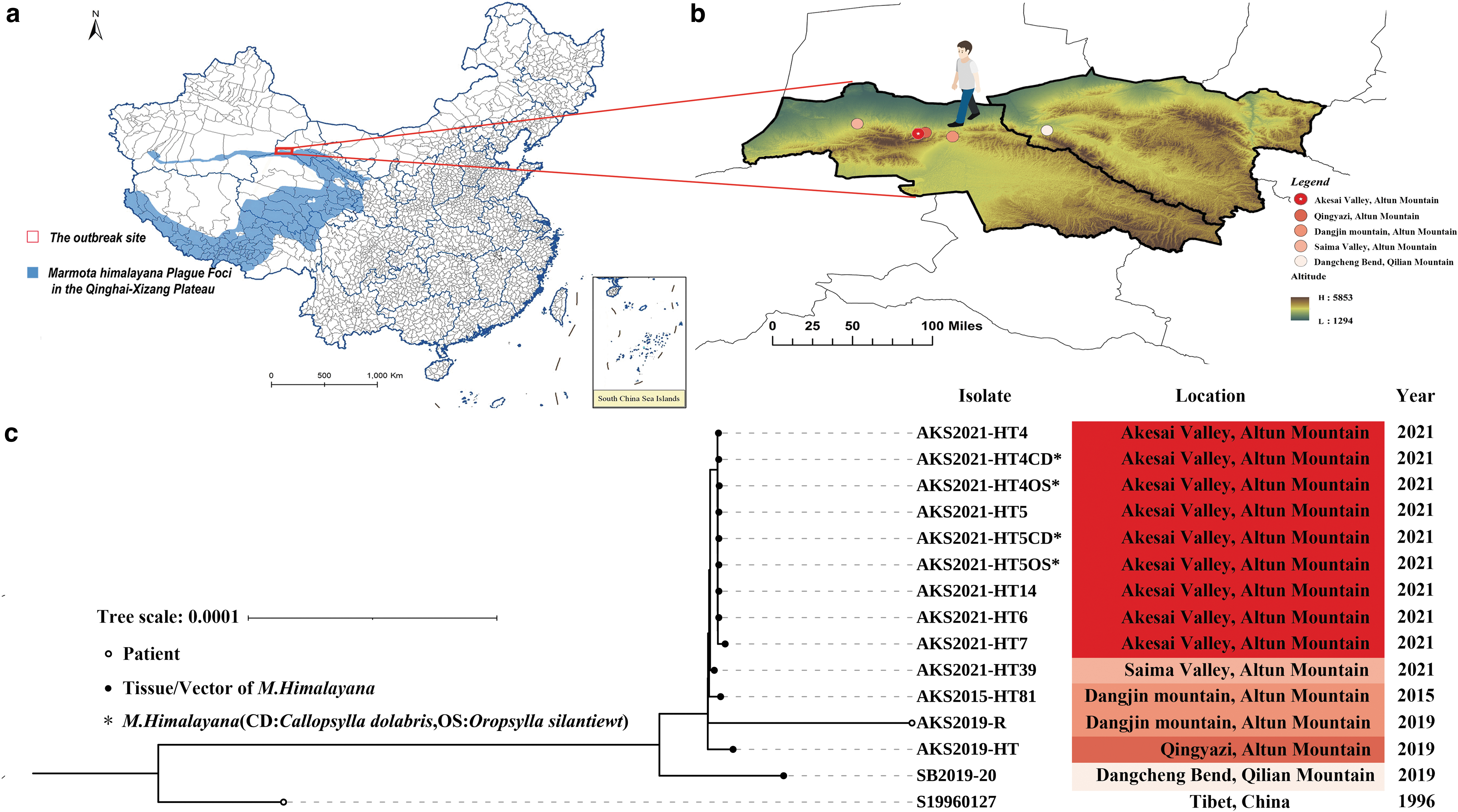
Geographical distribution and phylogenetic tree of the strains included in the whole genome sequencing and evolutionary analysis.
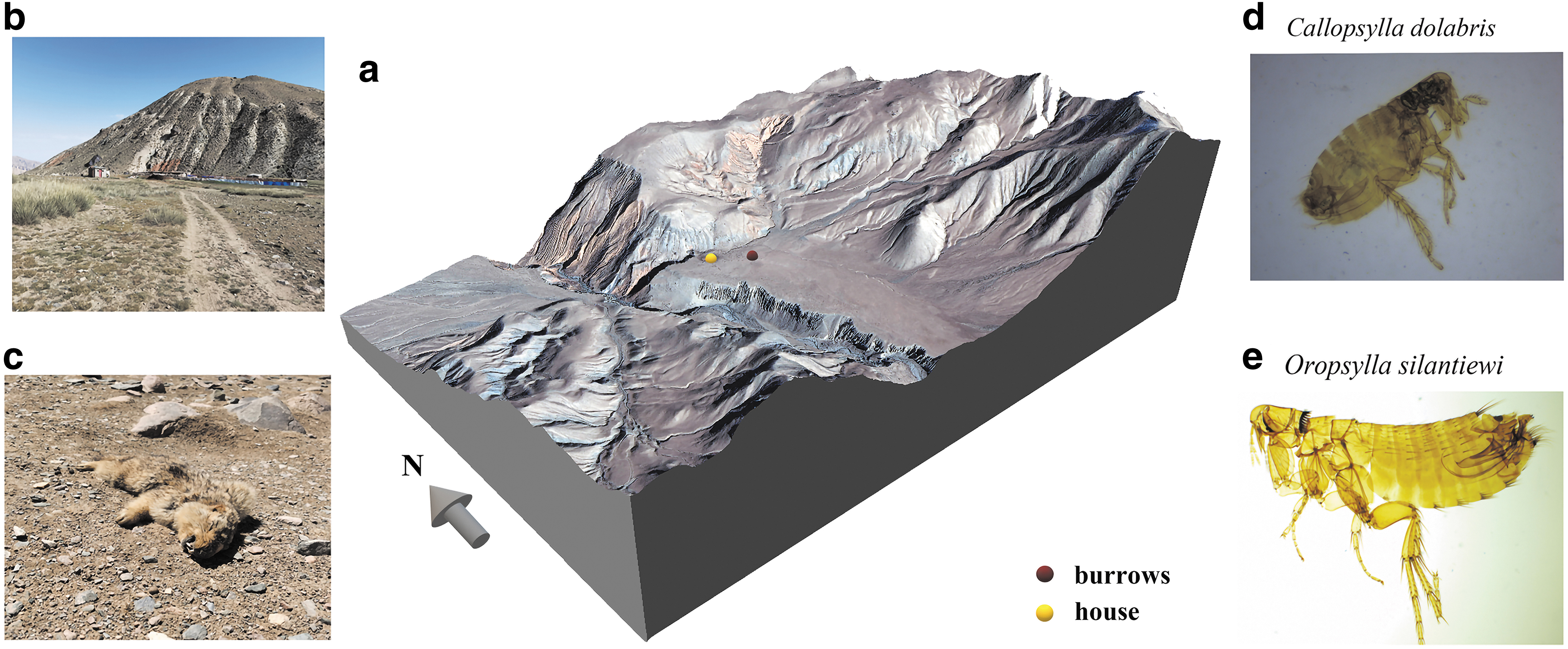
The location of the plague outbreak in an M. himalayana family and the microscopic image of the fleas collected from the marmots found dead in the environment in this study.
Specimens, including heart, liver, spleen, lung, kidney, intestine, tongue, and bone, were collected, and fleas were collected from the marmots and identified under microscopy by professionals at the local Centers for Disease Control and Prevention (CDC). The No. 5 marmot had splenomegaly and hyperemia, No. 7 had subcutaneous hyperemia, and No. 14 had an enlarged heart. Samples of the dead marmots and their fleas were inoculated onto Cepulodin Irgasan Novobiocin selective medium. The presence of Y. pestis was identified by bacteriophage lysis tests. Nucleic acids of all strains were then extracted and tested genes pla and caf1 (Bertherat and Jullien 2021).
Whole-genome sequencing and evolutionary analysis
The strains of Y. pestis isolated in this study and from the plague surveillance were all subjected to whole-genome sequencing. Complete genomes were sequenced by Pacbio Sequel; generated reads are assembled using the Hierarchical Genome Assembly Process algorithm. Draft genomes were sequenced by Illumina NovaSeq; generated reads were assembled using the SPAdes algorithm. A phylogenetic tree based on the core genome was constructed with the neighbor-joining algorithm in the MEGA X software package. Bootstrap analysis was performed with 1000 replicates. Evolutionary distances were computed using the Poisson correction method and are in units of the number of amino acid substitutions per site. All positions containing gaps and missing data were eliminated.
Pairwise single nucleotide polymorphisms (SNPs) were calculated between the compared strains. In addition to outbreak strains, five strains isolated from Altun Mountain and Qilian Mountain during plague surveillance were compared: AKS2015-HT81, isolated from a marmot found dead in Dangjin Mountain in 2015; AKS2019-R, isolated from a patient who died of plague after returning from a pastoral land in Dangjin Mountain; AKS2019-HT, isolated from a dying marmot in Qingyazi in 2019; AKS2021-HT39, isolated from a marmot found dead in Saima Valley; and SB2019-20, isolated from the region of Qilian Mountain (Table 1). The reference strain S19960127 of a plague patient was isolated from the Tibet Autonomous Region of China in 1996.
Information on Yersinia pestis Strains in the Marmot Familial Outbreak and Other Strains Compared in the Study
NCBI, National Center for Biotechnology Information.
The genome accession number of the outbreak strains
The complete genome sequences of the following three strains were deposited to Genbank under accession nos.: AKS2019-HT (CP084339-CP084342), AKS2019-R (CP084336-CP084338), and SB2019-20 (CP084343-CP084346). The draft genome sequences of 11 strains were deposited as: AKS2021-HT4 (JAIWLY000000000), AKS2021-HT4OS (JAIWMD000000000), AKS2021-HT4CD (JAIWME000000000), AKS2021-HT5 (JAIWLZ000000000), AKS2021-HT5OS (JAIWMG000000000), AKS2021-HT5CD (JAIWMH000000000), AKS2021-HT6 (JAIWMA000000000), AKS2021-HT7 (JAIWMB000000000), AKS2021-HT14 (JAIWMC000000000), AKS2021-HT39 (JAIWMI000000000), and AKS2015-HT81 (JAIWMF000000000).
Fraction 1 antibody and antigen detection
Blood of the marmot (No. 16) was sampled, and the serum was separated. The preliminary screening result of colloidal gold was positive, and the specific fraction 1 (F1) capsular antigen titer was then determined by indirect hemagglutination. The positive rates of F1 antibodies in the sera of marmots monitored from April to September of 2021 were compared. In this study, the antibody titer was transformed into reciprocal and analyzed by Wilcoxon signed-rank test.
Spleen index analysis
The ratio of spleen weight to the body weight of the six marmots found in the current study was combined with an analysis of the spleen weight index of marmots in 2021, and correlation with factors such as bacteria present in the animals was explored. A Mann–Whitney U test was used to determine whether there were differences in the proportion of spleen weight between the Y. pestis isolation group and the Y. pestis nonisolation group. The calculation formula for the spleen weight index was (Hadidi et al. 2008):
Ethics statement
The study was approved by the Laboratory Animal Welfare and Ethics Committee of the National Institute for Communicable Disease Control and Prevention of the Chinese Center for Disease Control and Prevention.
Results
Yersinia pestis strains isolated and compared
During this familial outbreak located at Akesai Valley, nine Y. pestis strains were isolated (see Table 1 for details): five isolated from five marmots found dead in the environment were designated as AKS2021-HT4, AKS2021-HT5, AKS2021-HT6, AKS2021-HT7, and AKS2021-HT14, respectively. Four strains isolated from fleas collected from marmots No. 4 and No. 5 were designated as AKS2021-HT4OS, AKS2021-HT4CD, AKS2021-HT5OS, and AKS2021-HT5CD (OS: O. silantiewi, CD: C. dolabris), respectively. Other strains compared in this study were four strains from Altun Mountains, one strain from Qilian Mountain, and the reference strain S19960127 from a plague patient in Tibet, China (Fig. 1).
Phylogenetic analysis
The phylogenetic trees based on 4007 core genes and 1,176,775 positions showed that the nine outbreak strains were tightly clustered (Fig. 1c). The strains isolated from Altun Mountain and Qilian Mountain of Gansu province are located on different branches of the phylogenetic tree, consistent with their geographic distance. The reference strains isolated from the Tibet autonomous region are located outside of all the strains from Qilian-Altun Mountain.
Pairwise SNP analysis
Pairwise SNP comparisons based on the reference strain revealed that there were 0–16 SNP differences among the outbreak strains and 2–115 SNP differences between the outbreak strain and other Qilian-Altun Mountain strains (Table 2).
Pairwise Single Nucleotide Polymorphism Differences Between Compared Strains in This Study
Analysis of F1 antibody titer
According to the analysis of surveillance data obtained in 2021, the positive rate of plague antibodies in serum of marmots was 28.00% (42/150), and the geometric mean titer (GMT) was 1:178 (Supplementary Table S1). The antibody titer of No. 16 marmot in Akesai Valley was 1:512, higher than the GMT of 1:178 (W = 391.00, Z = 2.81, p < 0.01).
The spleen index of marmots found dead infected with the plague was higher
The spleen index of the strain isolation group was higher compared with the strain nonisolation group. The distribution of the spleen index differed between the strain isolation group and the strain nonisolation group of Y. pestis from marmots found dead in the environment (U = 225.00, Z = −2.97, p < 0.05). The median of spleen index in the strain isolation group was 2.61, of which the upper and lower quantile are 3.70 and 2.09, respectively; and the median of spleen index in the strain nonisolation group was 1.62, while the upper and lower quantile are 2.57 and 0.81, respectively (Fig. 3).
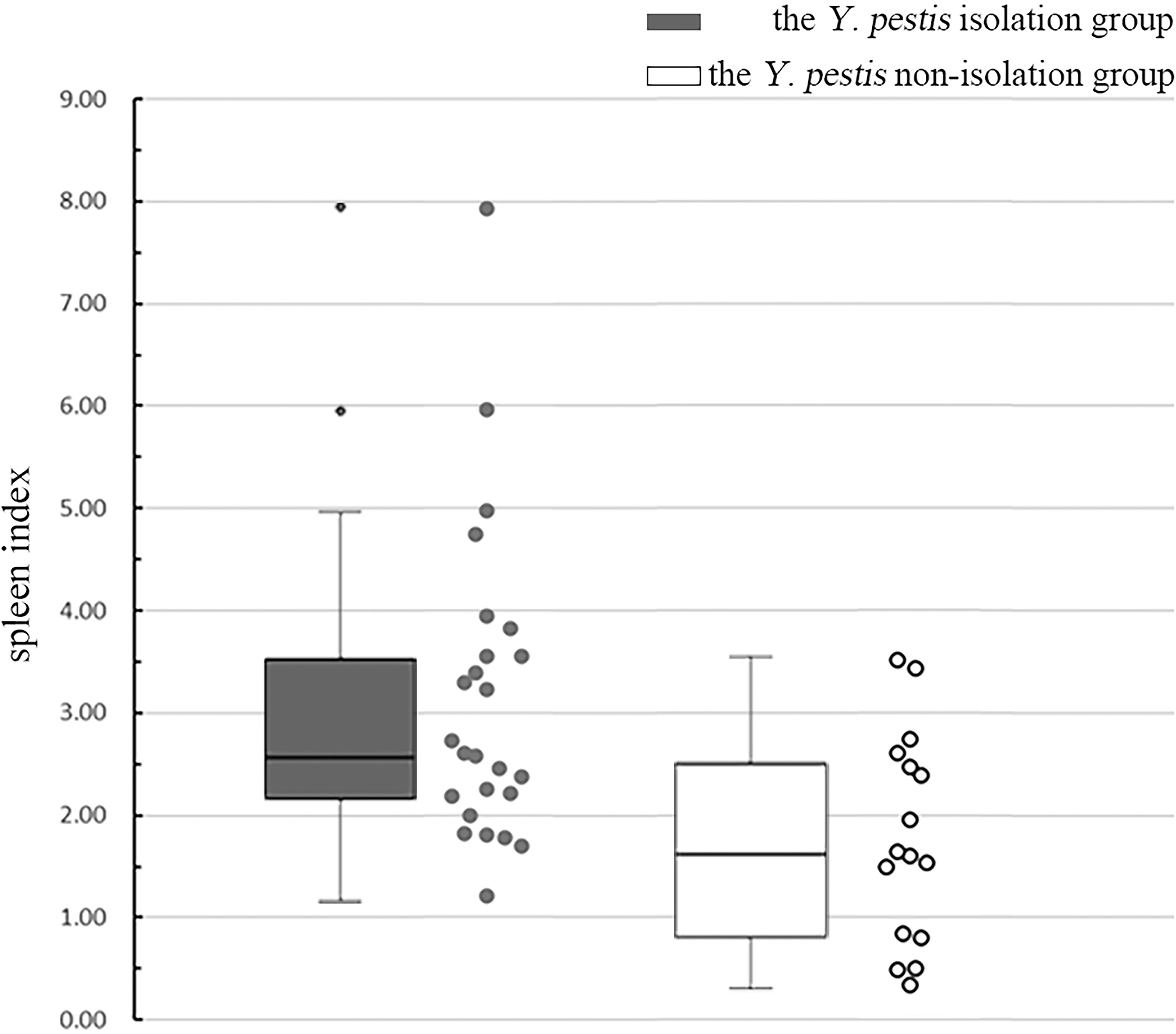
Box plots of spleen index of the strain isolation group and the strain nonisolation group of Yersinia pestis from marmots found dead in the environments. The middle region of each box plot (box body) covers 50% of the individuals, and the region between the upper and the lower transverse lines covers 100% of the individuals, whereas the points outside of the box represent transgressive individuals.
Discussion
In the M. himalayana plague foci of the Qinghai-Tibet Plateau, animal plague epidemics occur every year, sometimes with large-scale animal outbreaks and human cases. The spread of human plague epidemics from 2008 to 2017 was predominantly through contact with or skinning M. himalayana and was concentrated from June to December, with obvious seasonality (Xianhe et al. 2018). This is approximately the same as the epidemic period of the M. himalayana plague (May–September). The importance of surveillance of plague outbreaks in the M. himalayana plague foci is self-evident. Surveillance data of the Himalayan marmots showed that there were two epidemic peaks in the indirect hemagglutination assay (positive rate and F1 antibody titer): June was the main peak and September was the minor peak (Jie et al. 2011). M. himalayana hibernates from September to October each year through to the end of March or mid-to-early April of the following year (Liu and Tan 2000). This indicates that there is a possibility that some marmots infected in September may hibernate with bacteria.
In the current study, a family plague outbreak among M. himalayana was found in Akesai Valley in Gansu Province in April 2021, meaning that these six marmots had been infected with Y. pestis when they emerged from hibernation, and five of them were infected and dead. This further supports that M. himalayana may carry bacteria during hibernation (Fig. 2). Comparative genome analysis showed that Y. pestis strains isolated from the marmots found dead in the same burrow are highly identical (Fig. 1c); each strain has no more than 16 SNPs between each other (Table 2). The activity range of marmots is ∼3 km away from their cave (Gong and Shengwen 2012); therefore, isolates from marmots can reflect the local epidemic of Y. pestis, and phylogenetic analysis based on the core genome can identify outbreak strains of the same clone origin. The serum antibody titer of the live marmot from the same burrow was 1:512, and no strain was isolated from the marmot, which is a survivor of the familial outbreak. Together, these demonstrated the preservation and transmission of plague between marmot during and soon after hibernation.
Hibernation may be a factor in the prolongation of Y. pestis infection in rodents. The induction and expression of many genes of Y. pestis are mediated by temperature (Yother et al. 1986, Han et al. 2004, Korhonen et al. 2013, Chen et al. 2016, Quinn et al. 2019). Temperature control in Y. pestis inevitably involves mentioning the LCR phenomenon, which is the phenomenon that pathogenic Yersinia with Ysc-Yop type III secretion system (T3SS) genes requires for millimolar concentrations of calcium to grow at 37°C. In vitro, the LCR is observed as the inability to sustain growth, accompanied by the production of two virulence-associated antigens (V and W) when cultures free of Ca2+ are subjected to temperatures above 34°C, while LCR does not occur below 34°C, regardless of Ca2+ concentration (Yother et al. 1986). LCR and virulence of Y. pestis KIM5 are mediated by the trtA and trtB genes located on the 75.4-kilobase (kb) plasmid pCD1, and expression of these two genes is linked to the temperature-related positive regulatory gene lcrF (Yother et al. 1986).
Furthermore, temperature regulation of Ysc-Yop T3SS is mediated by controlling the transcriptional activator LcrF (Chen et al. 2016). In addition, the increase in temperature reduces the acylation of lipid A (Kawahara et al. 2002, Knirel et al. 2005). Temperature-mediated reduction of lipopolysaccharide (LPS) acylation leading to reduced cytokine inducibility is one of the mechanisms proposed to prevent the host immune system from recognizing pathogens in the early stages of infection (Korhonen et al. 2013). Changes in temperature and LPS are also suggested to affect the function of the gene pla. Plasminogen activation and autoprocessing of pla were higher in Y. pestis cells grown at 37°C compared with those grown at 20°C (Suomalainen et al. 2010). Moreover, RovA, the main regulator related to metabolism, stress adaptation, and virulence, and PsaA, a key Y. pestis mammalian virulence determinant that forms fimbriae, are both susceptible to temperature (Chen et al. 2016, Quinn et al. 2019).
In summary, the virulence of Y. pestis is related to temperature, and the expression and transcription of Y. pestis virulence genes will be upregulated at 37°C. When the body temperature of marmot is maintained at 5–10°C during hibernation, the reproduction of Y. pestis is relatively low (Shuli 1988). However, fleas of the genus Oropsylla are highly active vectors of plague at low ambient temperatures, 5–10°C. The vigor of rodent fleas and the transmission efficiency of plague are affected by environmental temperature, rainfall, and relative humidity (Ben-Ari et al. 2011, Cordes et al. 2020). Infected fleas at low temperatures can still cause an epidemic among animals (Baltazard and Bahmanyar 1960, Bazanova et al. 2007), which helps Y. pestis survive winter in the flea intestine and may cause infection during the hibernation or in the next transmission season (Schotthoefer et al. 2011, Williams et al. 2013). In addition, marmots in burrows wake up to 15–20 times during hibernation (Arnold 1988). At the same time, in the torpor state, their body temperature is 5–10°C, and when waking up in a state of euthermia, the temperature becomes normal, about 37°C.
To sum up, during marmot hibernation, infected Oropsylla fleas actively transmit the causative agent of plague between sleeping marmots. When waking up several times during hibernation with body temperature gradually returning to 37°C, the marmots go through the optimal growth temperature of Y. pestis. At this time, Y. pestis multiplies and increases virulence in infected marmots. So marmots may die out in whole families both during hibernation and after leaving wintering burrows in the spring months (Fig. 4). Therefore, during the winter months, marmots are prone to exacerbation of the infectious process.
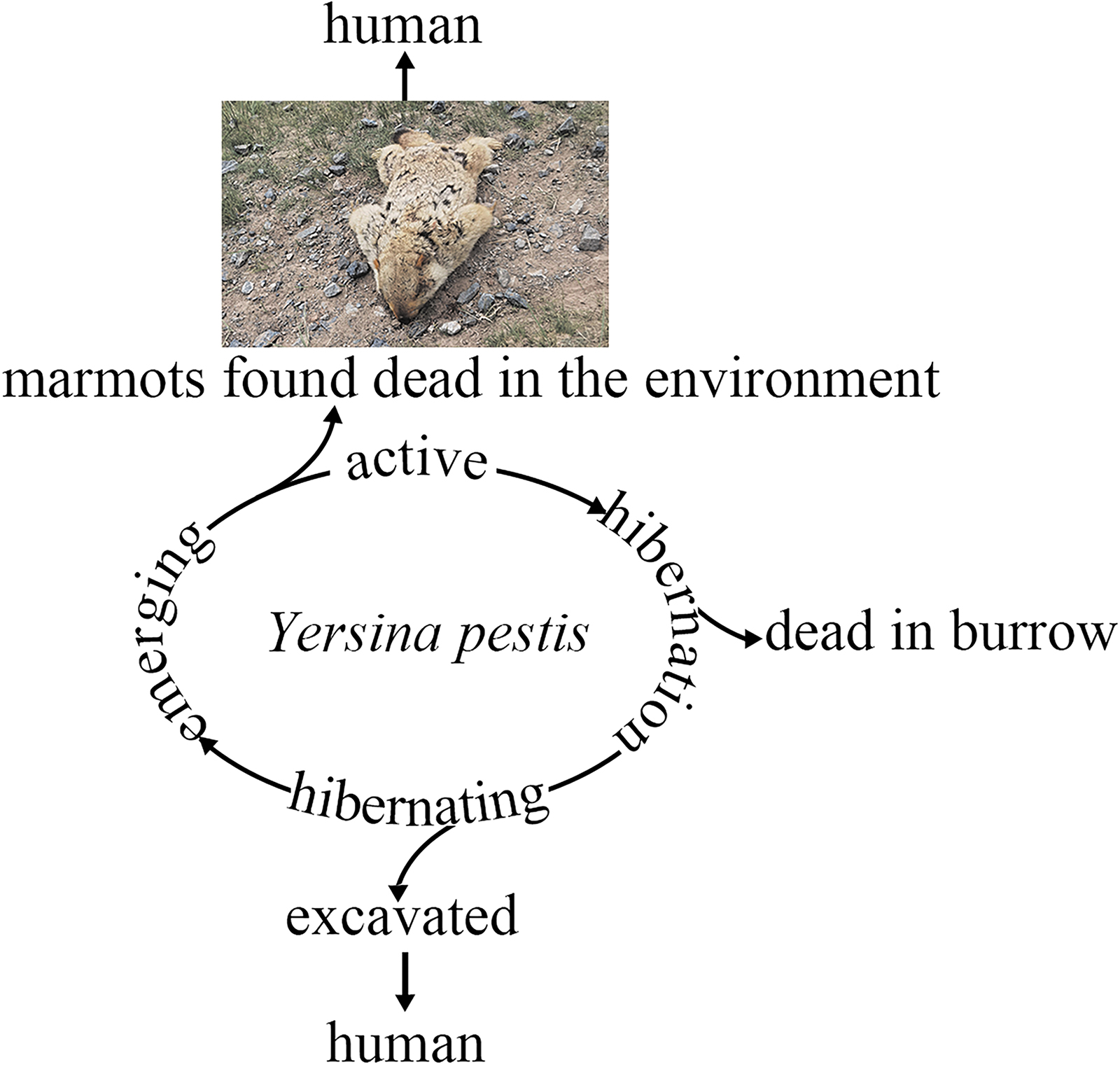
The life cycle of Y. pestis in M. himalayana and the risk of infection toward human during and after the hibernation period. The ring represents the life cycle of M. himalayana, and the upper and lower branches represent the transmission risk during the period of emergence and hibernation, respectively.
Whole-genome sequencing confirmed the outbreak of the marmot plague. Marmots are social family animals, and fleas still have a certain transmission ability at low temperatures. Therefore, it cannot be excluded that one of the six marmots in this study was infected with Y. pestis, and the marmots subsequently infected each other in the hibernating burrows resulting in an outbreak after hibernation. In this study, the strains AKS2021-HT4OS, AKS2021-HT4CD, AKS2021-HT5OS, and AKS2021-HT5CD obtained from fleas were highly homologous with the isolated strains of marmots in this family outbreak, which further supports the hypothesis that fleas spend the winter by parasitizing the hibernating M. himalayana.
There are 0–16 SNPs in a pairwise comparison between the outbreak strains, and even no SNP has been found between flea-isolated strain and marmot-isolated strains: AKS2021-HT4CD and AKS2021HT7, AKS2021-HT4CD and AKS2021-HT5 (Table 2). Other local strains isolated from the years 2019 and 2021 indicated that the epidemic strains in this region were relatively conserved and only have a small degree of variation. A possible factor for the continued prevalence of epidemic strains among animals in this region is that hibernation did not stop the prevalence of Y. pestis. The preservation mechanism of Y. pestis during animal hibernation may have played an important role in this continued prevalence.
F1 antibody titer reflects the epidemic level of plague. The antibody titer of marmot No. 16, a family member of the outbreak, reflected recent infection and a high possibility of infection during hibernation. The titer was higher than the GMT (1:178) of the surveillance data in the same year (W = 391.00, Z = 2.81, p < 0.01). This implies that the marmot was in a relatively high epidemic of plague state in the year.
Plague makes the spleen index larger. The distribution of spleen index differed between the strain isolation group and the strain nonisolation group of Y. pestis from marmots found dead in the environment (U = 225.00, Z = −2.97, p < 0.05); the spleen index of marmots found dead in the strain isolation group was higher than that in the nonisolation group. After an animal is infected by Y. pestis, the weight of its immune organs such as the spleen may change due to immune response, which is consistent with the characteristics of Y. pestis tending to invade the lymphatic system. The reactive proliferation of splenic lymphoid tissue, neutrophil infiltration in the sinusoid lacunae, and bacteria in phagocyte cells and consequently were observed in animal experiments (Heine et al. 2013, Wang et al. 2014, Li et al. 2017). Consequently, the spleen index also changes.
Hibernating marmots with Y. pestis have the potential to transmit the plague and may cause an outbreak after hibernation. Marmota himalayana hibernates from September-October to the following end of March-early April. The marmots gradually emerged in April, and the Y. pestis in the marmot and its vectors were easy to get the plague. The surviving marmots showed elevated serum antibody titers in the following months and reached peak strength in September for the next winter hibernation period (Fig. 4). In this study, a few tens of meters from the burrow where the dead marmots were found was a herdsman's house, used during grazing in winter (Fig. 1a). Fortunately, there are few people in this area in summer when the marmots are out of hibernation; if crowds of people and shepherd dogs were nearby at this time, or even if shepherds excavated M. himalayana, it would increase the risk of plague transmission.
Reviewing the sources and thematic studies of human plague cases from 2000 to 2014 in Gansu revealed that apart from the direct transmission of plague by skinning and eating marmots, the shepherd dogs are also important intermediate hosts that cannot be ignored in the plague transmission process. Consequently, herdsmen are more exposed to the wild than the general population, and the activities of shepherd dogs also increase the risk of infection in herdsmen (Wang et al. 2014, Ge et al. 2015). The location of the outbreak so close to the herdsman's residence further suggests that prevention and control of plague should not be relaxed in winter. Moreover, during the hibernation of marmots, some unique situations may occur, such as herdsmen peeling and eating the hibernating marmots and sheepdogs contacting hibernating marmots, which may cause human plague infection, and this could lead to further widespread transmission in crowds.
This investigation of a family outbreak of animal plague among M. himalayana shows during or soon after waking up from hibernation; marmots are sensitive and prone to Y. pestis and can die out in whole families. Active plague-infected fleas and several times of waking up in marmots during hibernation help Y. pestis transmission and reproduction, which is critical for the natural preservation of Y. pestis. Marmots die of plague not only after coming out of hibernation but also during the winter months in the burrow; plague control during the hibernation period cannot be let down.
Footnotes
Acknowledgment
The authors thank Charlesworth author services (paper no. 88929) for their critical editing and helpful comments regarding our article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by National Science and Technology Major Project (2018ZX10713-003-002 and 2018ZX10713-001-002).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.