
Abstract
Influenza A viruses (IAV) are zoonotic pathogens that can cause significant illness in wild and domestic animals, and humans. IAV can infect a broad range of avian and mammalian species, depending on subtype, and avian IAV can be moved over relatively long distances by migratory birds. Although spillover infections from wildlife or domestic animals to humans are an important part of the transmission cycle that can drive outbreaks, the relevance of companion animals, specifically dogs and cats, is not fully understood. A novel pandemic H1N1 reassortant (H1N1pdm09) emerged from swine in 2009, infecting humans, and wild and domestic animals worldwide. During a 2016 human influenza outbreak in Kyiv, subtype H1N1pdm09 predominated and was associated with severe disease and deaths; however, H3N2 and influenza B viruses were also detected. No case of avian influenza in humans was detected. To investigate potential involvement of companion animals, animals in a veterinary hospital (116 cats and 88 dogs) were randomly selected, and sera were tested using a commercially available IAV nucleoprotein enzyme-linked immunosorbent assay. Twelve of 203 serum samples were positive for influenza antibodies (5.7% of dogs and 6.08% cats). These are the first data to demonstrate influenza A infection of pets in Ukraine, highlighting the potential risk of infection of companion animals from close contact with humans.
Introduction
Influenza A
Canine influenza results from two main subtypes: H3N8 originated in horses, and H3N2 originated in birds (Parrish et al. 2014). Both subtypes are currently circulating in America, Europe, and Asia (Knesl et al. 2009, An et al. 2010, Bunpapong et al. 2014, Schulz et al. 2014, Sen et al. 2016, Zhou et al. 2016, Voorhees et al. 2017, Watson et al. 2017, Aydin et al. 2018). The H3N8 equine influenza strain circulating among horses adapted to dogs without genetic reassortment, generating a new canine-specific H3N8 lineage (Waddell et al. 1963). Cross-species infections of dogs have occurred independently on several occasions, generating specific lineages of H3N8 canine influenza virus (CIV), suggesting there may be adaptive barriers (Crawford et al. 2005, Daly et al. 2008, Crispe et al. 2011).
In the United States, a serological survey of dogs with respiratory disease in 2004–2005 found antibodies to H3N8 CIV (Dubovi and Njaa 2008, Gibbs and Anderson 2010). In 2007, severe respiratory disease in dogs was reported in South Korea. This zoonosis was caused by a distinct avian-derived H3N2 subtype CIV, which became established in South Korean and Chinese canine populations, with persistent dog-to-dog transmission and a different internal gene constellation from human seasonal H3N2 viruses (Song et al. 2008, 2009).
Human pandemic subtype H1N1 IAV, seasonal subtypes H3N2 and H1N1, and avian subtypes H5N1, H5N2, H5N8, and H9N2 influenza viruses have also occasionally infected dogs (Chen et al. 2010, Lin et al. 2012, Zhan et al. 2012, Sun et al. 2013, Song et al. 2015, Yuk et al. 2017). Interestingly, porcine H1N1 viruses have frequently been transmitted to dogs and reassorted with avian-origin H3N2 CIV. Consequently, in Asia, four novel reassortant CIV genotypes, namely H1N1r, H1N2r, H3N1r, and H3N2r, have evolved (Song et al. 2012, Chen et al. 2018).
Respiratory tract infections in cats are often generally referred to as “cat flu.” In 2011, canine influenza H3N2 was isolated from cats in Korea, suggesting a potential role for cats as an alternate host for H3N2 IAV. In addition, several studies reported cases of highly pathogenic avian influenza (HPAI) H5N1 in naturally infected domestic cats in Thailand, Germany, Austria, and Iraq (Songserm et al. 2006a, Wolf et al. 2006, Yingst et al. 2006, Leschnik et al. 2007). Experimental infection of cats with avian H5N1 isolated from a human case is frequently fatal, with documented cat-to-cat transmission (Rimmelzwaan et al. 2006). Sporadic infection of cats has been reported for several IAV subtypes, including avian-like H7N2, H5N2, and human-like H1N1pdm09 (pandemic H1N1), and seasonal H3N2 and H1N1 (Sponseller et al. 2010, Ali et al. 2011, Fiorentini et al. 2011, Chow and He 2014, Hai-xia et al. 2014, Lee et al. 2017).
Emergence of new influenza viruses in domestic carnivores with the potential for zoonotic infection may constitute a public health threat (Song et al. 2011, Jeoung et al. 2013). IAV in domestic canine and feline companion animals could represent additional intermediate or reservoir hosts for generation of new pathogenic strains of IAV through genetic reassortment. Moreover, as many human diseases have zoonotic origin, surveillance in wild and domestic animals can serve as an early warning to anticipate a human health threat (Neo and Tan 2017). With this in mind, we conducted a serological survey to determine if there is exposure of companion domestic dogs and cats to IAV in Ukraine. We found evidence of IAV infection of these species, highlighting their potential role in the transmission of IAV.
Materials and Methods
From March to May 2016, a total of 203 serum samples from 88 domestic dogs and 115 domestic cats were collected and biobanked during standard clinical procedures in a veterinary hospital in Kyiv, Ukraine, with approval of pet owners for routine serum-based diagnostic procedures. Animals were sampled randomly based on availability and none displayed influenza-like symptoms. Among the 203 sera, 91, 73, and 39 serum samples were collected in March, April, and May, 2016, respectively. All samples were immediately frozen and stored at −20°C until transport to the Institute of Veterinary Medicine (IVM, Kyiv) for serological testing. Upon receipt, samples were thawed, centrifuged at 1073 × g (4000 rpm/6 cm) for 15 min at room temperature, aliquoted, and stored at −80°C until tested for antibodies against IAV.
A specific competitive enzyme-linked immunosorbent assay (epitope-blocking ELISA) was performed utilizing an Influenza A Virus Antibody Test kit (IDEXX) according to manufacturer's protocols. ELISA pates were analyzed on an iMark Microplate Absorbance Reader (Bio-Rad). The IDEXX Influenza A Virus Antibody Test kit was used as a first screening test to detect serum antibodies (IgG) against IAV nucleoprotein (NP). Samples with S/N value (sample to negative control ratio) less than 0.6 were considered positive. NP is highly conserved and enables detection of antibodies to any IAV subtype across avian, swine, equine, canine, feline, and other mammals.
All sera tested positive by the NP ELISA were retested with a competitive N1 ELISA (ID Screen Influenza N1 Antibody Competition). The results are expressed in percentages and samples with a value less than 60% were considered positive. Originally, the kit was developed to analyze serum from avian species for the presence of anti-N1 IAV antibodies. However, the kit potentially can be used as a multispecies assay using a monoclonal antibody against influenza A N1 surface glycoprotein, without the need for species-specific secondary antibodies (conjugate). This kit has successfully been employed as an additional serological test for influenza N1 antibodies in domestic animals during serological surveys for influenza H1N1pdm09 in Germany (Damiani et al. 2012) and Differentiation of Infected from Vaccinated Animals (DIVA) analyses (Dundon et al. 2007).
To determine whether or not the N1 ELISA could produce false-positive results, a panel of NP ELISA-negative sera was assayed with the N1 ELISA kit (Damiani et al. 2012). In our study for that purpose, 30 cat and 30 dog NP-negative sera were used, as determined by the IDEXX NP ELISA kit, and no false-positives result was observed. The 95% exact binomial confidence interval of the percentage of positive samples was assessed by using the epitools package (Aragon 2017) and the R environment for statistical computing (R Core Team 2019).
Results
Twelve of the 203 serum samples were positive for influenza A antibodies by the NP ELISA test for a seroprevalence of IAV of 5.7% in dogs and 6.08% in cats (Table 1). Employing N1 subtype-specific ELISA, antibodies against N1 IAV were detected in six of the samples: 1/5 dog sera and 5/7 cat sera (Table 2). At the time of sampling, the seropositive animals had no apparent signs of respiratory or parasitic diseases, and complete blood cell counts, serum biochemistry, and infectious disease test results were normal or negative for each of the 12 cases (Table 2). The sample collection protocol precluded the ability to question owners on their health status. However, serological evidence of the N1 in these pets could suggest that transmission of influenza H1N1pdm09 may have occurred, possibly by close contact with IAV-infected owners during the peak period of virus infection in the human population in late 2015, or early 2016 (Fig. 1).
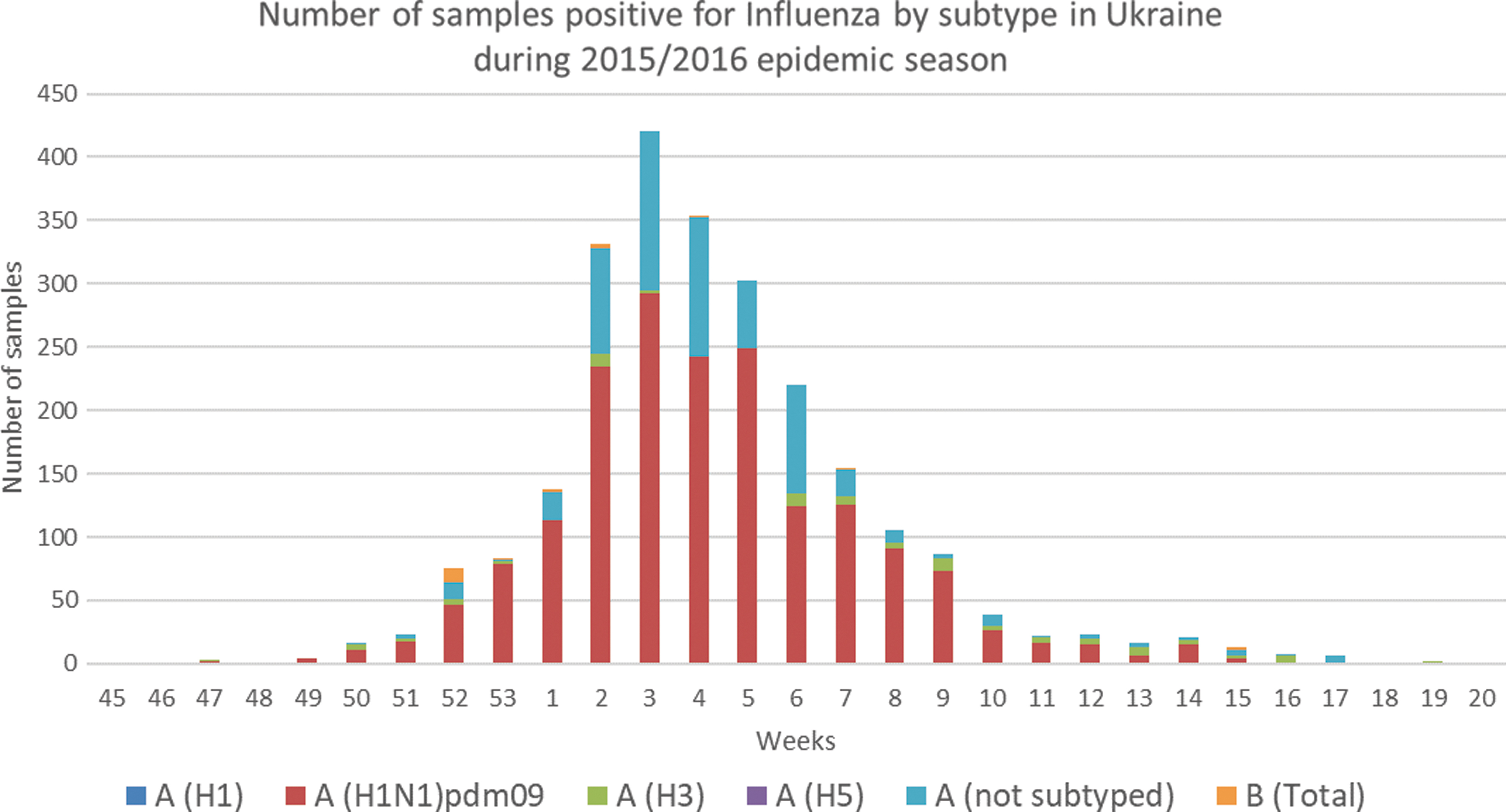
Epidemic curve of influenza virus in the human population in Ukraine during 2015/2016 epidemic season. Data were collated from a publicly reported source: FluNet (
Seroprevalence of Influenza A Virus in Domestic Carnivores in Kyiv, Ukraine
IDEXX Influenza A NP blocking ELISA, results are expressed as S/N values (sample to negative control ratio): neg (≥0.6); pos (<0.6) indicating positive for IAV anti-NP antibodies.
Statistical analysis by the epitools package and the R environment for statistical computing.
CI, confidence interval; ELISA, enzyme-linked immunosorbent assay; IAV, influenza A viruses; NP, nucleoprotein.
Characteristics of Animals Seropositive for Influenza A Antibodies by Nucleoprotein-Blocking Enzyme-Linked Immunosorbent Assay
IDEXX Influenza A NP blocking ELISA, results are expressed as S/N values (sample to negative control ratio): neg (≥0.6); pos (<0.6).
ID Screen® Influenza N1 ELISA competition percentages (ODsample/ODneg × 100): neg (>60%); pos (<60%).
Discussion
With respect to zoonotic diseases, the health of companion animals is of particular concern because of their close contact with humans and risk for reverse zoonoses, particularly for human H1N1pdm09 IAV. Dogs and cats are a popular companion animal and, although there is no centralized registration process, it is believed that more than half of Ukrainians currently own pets and perceive their pets as family members (
In this report, we provide evidence for domestic dog and cat exposure to IAV using a competitive ELISA method for serological testing. Twelve serum samples were positive for IAV NP antibodies by the NP ELISA test, strongly suggesting prior infection of these pets, as the animals were not overtly symptomatic at the time of sample collection. Since samples were collected during/after the human 2015–2016 influenza season, we speculate, but cannot confirm that infections probably resulted from transmission through close contact with infected owners or contact with other animals carrying influenza viruses.
This is not unprecedented, as several studies have suggested that infection of cats and dogs resulted from close contact with humans. The first cases of lethal pandemic H1N1pdm09 influenza virus infection in domestic cats in 2009 in the United States were likely infection by contact with infected owners (Lὅhr et al. 2010). A serosurvey of cats in the USA during the 2009–2010 influenza season found seroprevalences of pandemic H1N1pdm09 (21.8%), human seasonal H3N2 (25.6%), and H1N1 (41.9%) influenza viruses (McCullers et al. 2011). At the end of 2009, China first reported detecting H1N1pdm09 in domestic dogs with clinical signs, and H1N1pdm09 was confirmed in a dog associated with a sick pet owner in the United States (Keenliside 2013).
In 2009, pandemic IAV H1N1pdm09 was also directly isolated from dead cats in Italy. Subsequent serological investigation showed that 55.0% of the animals tested had antibodies against the H1N1pdm09 strain, concurrent with the peak of the human flu pandemic in Italy (Fiorentini et al. 2011). Seroprevalence of domestic cats (1.93%) and dogs (0.13%) to pandemic H1N1pdm09 in Germany in 2010 and 2011 suggested humans as the source of infection (Damiani et al. 2012).
Serological evidence for H1N1pdm09 with a relatively high prevalence was reported for cats in China in 2011–2013, suggesting that transmission occurred by close contact with infected owners or other companion animals (Su et al. 2013, Zhao et al. 2014). In 2012 in China, a high prevalence (24.7%) of antibodies against influenza H1N1pdm09 was also reported in dogs. The researchers hypothesized that close, prolonged contact between dogs and influenza-infected humans led to this significant seroprevalence (Su et al. 2014). In Japan, a seroprevalence survey of cats and dogs between 1997 and 2008 found antibodies against the human H1, H3, and N2 subtypes of IAV at a relatively low prevalence (Said et al. 2011).
In the epidemiology of influenza viruses, interspecies transmission plays a key role, and the close contact between companion animals and humans should not be overlooked. In dogs, H3 subtypes of CIV are frequently found. Serological evidence of H1N1pdm09 influenza virus infection alone and in combination with the canine H3N2 was found in serum samples collected from dogs in Korea in 2010 and 2011 (Song et al. 2015). The same results were reported in the USA; two dogs were seropositive for two subtypes (H1N1 and H3N2 and H1N1 and CIV H3N8) (Jang et al. 2017). These studies indicate that although dogs can be infected with more than one virus subtype, retrospective serology studies cannot discriminate between simultaneous and serial infections. However, reassortants apparently can be found in dogs. For example, a novel reassortant canine H3N1 between pandemic H1N1 and canine H3N2 IAV (Song et al. 2012), and swine-origin H1N1 viruses reassorted with CIV H3N2, creating three novel strains (H1N1r, H1N2r, and H3N2r). The authors noted that specific factors in China, in particular dog meat markets and a large number of feral dogs, make canine-to-human transmission of IAV more likely, representing a potential pandemic threat (Chen et al. 2018).
The recent study in Poland has shown 2.21% prevalence of IA antibodies in dogs using commercial ELISA kit, and with additional hemagglutination inhibition assays also found that the most frequently detected antibodies were to swine H3N2 (Kwasnik et al. 2020).
In this report, antibodies against N1 IAV were detected in one of the five positive dogs and five of the seven positive cats, but given the low overall seropositivity in our study, the difference in detection rate of N1 IAV between dogs and cats is not significant. This suggests that transmission could have occurred by contact with their infected owners during the 2015/2016 epidemic season in Ukraine. A country-specific risk assessment confirmed the 2015/2016 influenza season in Ukraine was more severe and started earlier than the previous four seasons (Newitt et al. 2018). The predominant strain circulating was influenza virus H1N1pdm09 and was associated with severe disease and deaths (Fig. 1). While the intensity of H1N1pdm09 activity was estimated to be higher than usual, influenza H3N2 and influenza B viruses were also detected (World Health Organization 2019).
Our study reports the first serological evidence demonstrating antibodies to N1 subtype IAV in domestic carnivores living in Kyiv, suggesting these animals were exposed to an N1-carrying mammalian IAV subtype, likely H1N1pdm09. This is the only circulating N1 virus in mammals in Ukraine, as far as is known. Although influenza activity was reported across the country, the capital Kyiv was the most affected area with the highest incidence (Worldwide Influenza Centre and Francis Crick Institute Report 2016, Newitt et al. 2018). Further studies are needed to determine the incidence of infection of cats and dogs in other regions, and to determine whether the frequency of infection is correlated with the incidence of human influenza cases. Interestingly, the risk assessment of the WHO European Region confirmed that there was higher influenza activity in Eastern Europe in 2015/16 compared to Western Europe with a strong predominance of H1N1pdm09 (Tjon-Kon-Fat et al. 2016).
In our study, the four dogs and two cats that were seronegative for N1 subtype clearly had antibodies against influenza NP, suggesting exposure to other subtypes of IAV. Further influenza A subtyping, including analysis of HA (H1, H3, H5, etc.) subtype, was not carried out in our study due to the limited number of reference sera and reagents available. While we detected anti-N1 antibodies in companion animals, we do not believe that H5N1 infection occurred in cats or dogs, since no case of the H5N1 in birds or humans was registered in Kyiv or other regions of Ukraine in that period.
However, dogs and cats can be infected with other strains of IAV. Avian-origin influenza viruses have been identified particularly in cats, including HPAI H5N1, H5N6, and H5N8, LPAI H5N2, H6N1, H7N2, and H9N2 (Songserm et al. 2006b, Leschnik et al. 2007, Zhan et al. 2012, Sun et al. 2013, Hai-xia et al. 2014, Lin et al. 2015, Yu et al. 2015, Yuk et al. 2017, Hatta et al. 2018). Infection of domestic carnivores by avian IAV occurs most likely through the ingestion of or close contact with infected poultry or wild birds (Reperant et al. 2009). In one case, feline H7N2 viruses spread among cats and also infected humans (Marinova-Petkova et al. 2017, Poirot et al. 2018).
Before the onset of the peak influenza season, we believe it is important that pet owners and household members are advised of the potential risks that they may pass influenza virus not only to other humans but also to other animals, including pets. This concept of “reverse zoonosis” raises concerns about mammalian adaptation of avian IAV, antigenic shift mutations, reassortment, and emergence of novel viral subtypes. Complete IAV subtyping was not conducted in our study due to the limited availability of reference sera and reagents. Thus, what other subtypes may be circulating in Ukraine remains unclear. However, our study demonstrates that domestic cats and dogs were exposed to IAV, likely the H1N1pdm09 subtype.
This is the first study that reports serological evidence for IAV infection in domestic pets in Ukraine. When and how seropositive animals were infected could not be determined. We assume that the contact with IAV might have happened by close communication between infected owners and their pets during the intense 2015/2016 human influenza season in Ukraine. It is reasonable to suppose that many more cases may have been present, and we believe that continued surveillance of IAV in domestic animals is warranted.
Conclusions
Our study reports the first serological evidence for antibodies to IAV in domestic dogs and cats in Kyiv, Ukraine. Although serological surveys are limited by available antigens and reference sera, they are used as a first step in exploring key signals and evidence for zoonotic infectious disease transmission. We assume that the contact with IAV might have happened by close communication between infected owners and their pets during the 2015/2016 influenza season in Ukraine. As influenza viruses cross species barriers, carrying out a serological surveillance in dogs and cats is also warranted, taking into account the close contact between humans and companion animals, and potential for circulation among cats and dogs. These results highlight that further studies in these species are needed to determine the precise role they could play in the natural cycle of influenza virus transmission.
Footnotes
Acknowledgments
We express our special thanks to the veterinarians from the veterinary hospital in Kyiv for excellent technical assistance and serum sampling. Dr. Stephen Higgs (Kansas State University) and Eric Bortz (University of Alaska Anchorage) generously provided their mentorship throughout the development and review of this article. The authors would like to thank the Defense Threat Reduction Agency (DTRA) and Biological Threat Reduction Program (BTRP) in Ukraine for the support in developing this publication. While DTRA/BTRP did not support the research described in this publication, the Program supported the writing mentorship provided within the Science Writing Mentorship Program and the publication effort. The content of this publication is the responsibility of the authors and does not necessarily reflect the views of DTRA or the United States Government.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the National Academy of Agrarian Sciences of Ukraine.