
Abstract
In the Mekong Delta in southern Vietnam, rats are commonly traded in wet markets and sold live for food consumption. We investigated seroprevalence to selected groups of rodent-borne viruses among human populations with high levels of animal exposure and among co-located rodent populations. The indirect fluorescence antibody test (IFAT) was used to determine seropositivity to representative reference strains of hantaviruses (Dobrava virus [DOBV], Seoul virus [SEOV]), cowpox virus, arenaviruses (lymphocytic choriomeningitis virus [LCMV]), flaviviruses (tick-borne encephalitis virus [TBEV]), and rodent parechoviruses (Ljungan virus), using sera from 245 humans living in Dong Thap Province and 275 rodents representing the five common rodent species sold in wet markets and present in peridomestic and farm settings. Combined seropositivity to DOBV and SEOV among the rodents and humans was 6.9% (19/275) and 3.7% (9/245), respectively; 1.1% (3/275) and 4.5% (11/245) to cowpox virus; 5.4% (15/275) and 47.3% (116/245) for TBEV; and exposure to Ljungan virus was 18.8% (46/245) in humans, but 0% in rodents. Very little seroreactivity was observed to LCMV in either rodents (1/275, 0.4%) or humans (2/245, 0.8%). Molecular screening of rodent liver tissues using consensus primers for flaviviruses did not yield any amplicons, whereas molecular screening of rodent lung tissues for hantavirus yielded one hantavirus sequence (SEOV). In summary, these results indicate low to moderate levels of endemic hantavirus circulation, possible circulation of a flavivirus in rodent reservoirs, and the first available data on human exposures to parechoviruses in Vietnam. Although the current evidence suggests only limited exposure of humans to known rodent-borne diseases, further research is warranted to assess public health implications of the rodent trade.
Introduction
I
Given the frequency and intensity of human and livestock contact with rodents, we aimed to explore potential risks associated with these exposures through seroprevelance studies. We determined seroprevalence to five groups of known zoonotic viruses in rodents representing the common species sold in wet markets and present in peridomestic and farm settings. In parallel, we determined seroprevalence to the same set of viruses in a cohort of individuals with high levels of occupational and residential exposure to rodents and domestic livestock. The viruses of interest (namely flaviviruses, hantaviruses, poxviruses, arenaviruses, and rodent parechoviruses) were selected on the basis of their known or probable endemicity, their documented associations with rodent-borne disease, and considerations of feasibility and resources (i.e., access to reference strains and well-established diagnostic protocols). In addition to serological screening, we tested rodent liver and lung tissues by RT-PCR using generic primers for flaviviruses and hantaviruses).
Methods
Rodents
Rodent surveys were conducted as described by Hoang et al. (2014). Briefly, rats were purchased from markets in five provinces of the Mekong Delta (Dong Thap, Tien Giang, An Giang, Vinh Long, and Can Tho) during October of 2012, and field trapping was conducted in peridomestic and forest habitats in Dong Thap province during March of 2013. Specimens used in this investigation comprised sera or whole blood and tissues from 275 individual rats (150 purchased in wet markets, 125 field trapped). Five rodent species were represented: Rattus argentiventer (n=104) Bandicota indica (n=65), R. tanezumi (n=61), R. norvegicus (n=29), and R. exulans (n=16) (Table 1).
Human sera and data collection
Sera were obtained from a cohort of 245 people living in farming communities in Dong Thap Province. Ethical approval was obtained by the Oxford Tropical Research Ethics Committee and the People's Committee of Dong Thap Province, and the study was implemented by the provincial Preventive Medicine Center (PMC). Participants were eligible if they gave informed consent and were involved in raising, slaughtering, or processing livestock or wildlife, with a minimum of 10 h exposure per week. Family members with residential exposures to livestock were also recruited (minimum 1 year of age). Phlebotomy was performed using sterile technique by a study nurse. After collection, all specimens were aliquotted and preserved at −80°C. Information was collected on sociodemographic factors, occupation, house structure, hygiene, water source, food consumption, household animals, detailed animal contact histories, vaccination, and admission to hospital in the 12 months prior to the survey.
Serological screening
Human and rodent sera, and rodent dried blood spots on filter papers were screened using an indirect immunofluorescence antibody test (IFAT) for antibodies against Seoul (SEOV) and Dobrava (DOBV) hantaviruses (representatives of rat and mouse type host lineages, respectively); flaviviruses (tick-borne encephalitis [TBEV] serocomplex); arenaviruses (lymphocytic choriomeningitis virus [LCMV]); poxviruses (cowpox virus); and parechoviruses (Ljungan virus). IFAT spotslides and the assay procedures were conducted as previously described (Kallio-Kokko et al. 2006, Jääskeläinen et al. 2013) using Vero E6 cells for SEOV, DOBV, TBEV, LCMV, and Vero cells for cowpox and Ljungan viruses. Viral reference strains were 80-39 for SEOV (Antic et al. 1991), Ano-Poroia for DOBV (Nemirov et al. 1999), Armstrong for LCMV (Kallio-Kokko et al. 2006), Kumlinge A52 for TBEV (Brummer-Korvenkontio et al. 1973; Whitby et al. 1993), CPXV/FIN/T2000 for cowpox virus (Pelkonen et al. 2003), and LV 145 SLG for Ljungan (Tolf et al. 2009). Sera were diluted 1:20 in phosphate-buffered saline (PBS) for initial screening, and repeat testing with three-fold serial dilutions was performed on all human putative seropositives. Whole blood collected in filter papers was eluted in PBS (∼1 cm2 of filter paper in 1 mL of PBS). For rodent sera, the secondary antibody was polyclonal rabbit anti-mouse fluorescein isothiocyanate (FITC) conjugate diluted 1:30 (Dako A/S, Copenhagen, Denmark); for human sera, the secondary antibody was anti-human immunoglobulin G (IgG) FITC conjugate diluted 1:100 (Jackson Immuno Research Laboratories, West Grove, PA). Each slide was examined independently by two researchers using an Olympus BX51 microscope. Human sera with evidence of TBEV reactivity were further examined using differential IFAT screening in parallel for dengue (DENV) and Japanese encephalitis virus (JEV), as previously described (Tonteri et al. 2011).
Molecular screening
Virological screening for flaviviruses and hantaviruses was performed on all rodent liver and lung samples. Briefly, 1-cm2 tissue samples were homogenized using a bead mill (TissueLyser II, QIAGEN), and nucleic acid extracted using MagNA Pure 96 Viral NA Small Volume Kit (Roche) on an automated extractor (Roche). Following extraction, nucleic acid samples were pooled by species (five samples per pool). cDNA from lung samples was screened for hantavirus with conventional PCR (L segment) (Klempa et al. 2006). cDNA from liver tissues was screened for flaviviruses using consensus NS5 primers (Kuno et al. 1998). When pools tested positive, original extractions from individual samples were further analyzed. Amplicons of the expected band size were sequenced commercially (Macrogene, Seoul, Korea).
Statistical analyses
Comparison of human seropositivity to hantavirus (DOBV and SEOV combined), pox, LCMV, and Ljungan viruses was carried out using a chi-squared test. Risk factor analyses to explore associations between demographic/occupational variables, animal contact histories, and seroprevalence were investigated using logistic regression and multivariable models (Dohoo et al. 2003). All statistical analyses were carried out using R statistical software (
Results
Rodent serology
Among the 275 tested rodents, seropositivity was detected in four of the five virus groups, with moderate levels of reactivity to TBEV (5.4% seropositive) and hantaviruses (6.9% seropositive), but little or no evidence for immunological exposure to cowpox (1.1%), arenavirus (0.4%), or Ljungan virus (0%) (Table 2). Among the rats with TBEV antibodies, B. indica had the highest prevalence (12/65, 18.4% positive), followed by R. argentiventer (2/104, 1.9%) and R. tanezumi (1/61, 1.6%) (p<0.001). To examine whether the TBEV seropositives represented true reactivity to TBEV serocomplex viruses or cross-reactivity to the other regionally endemic flaviruses, additional serial titrations (five dilutions) were run in parallel against DENV and JEV for the 15 putative TBEV seropositives and for five seronegatives. The differential IFAT titration showed similar reactivity to TBEV, DENV, and JEV, and all samples tested negative to the three viruses at 1:320 dilution.
Values within parentheses are percentages.
Overall seropositivity to hantaviruses was 6.9% (19/275). In spite of the supposed cross-reactivity among murine hantaviruses, only three individuals were positive for both hantaviral antigens (six positive for DOBV only, 10 positive for SEOV only). This differential reactivity may indicate the presence of several hantaviruses. Seropositive host species included R. norvegicus, R. argentiventer, R. tanezumi, and B. indica. None of the R. exulans sera (n=16) showed antibody reactivity to the test viruses.
Rodent molecular screening
None of the 275 rodent liver samples (including those from the 15 TBEV-seropositive rodents) tested positive when screened with the generic NS5 flavivirus primers. From hantavirus screening of lung tissues, one R. argentiventer market sample yielded an amplicon of appropriate band size in the hantavirus conventional RT-PCR assay. A 310-bp sequence of the L segment confirmed the presence of a hantavirus with a 95–98% match to all SEOV sequences in GenBank, the closest one being strain Jakarta/Rn137/2000 from Indonesia (new GenBank acc. no. KP055755).
Human serology
Seropositivity was detected to all five virus groups, with high levels of seropositives to TBEV (116/245, 47.3%), moderate seropositivity to Ljungan virus (18.8%, 46/245), and lesser rates of seroposivity to cowpox virus (4.5%, 11/245), hantaviruses (3.7%, 9/245), and LCMV (0.8%, 2/245) (Table 3). Regarding the nine individuals with hantavirus antibodies, there were two positive only for DOBV, five positive only for SEOV, and two positive for both. Seropositivity to TBEV and poxviruses was associated with increased age, whereas seropositivity to Ljungan virus was higher in younger individuals (Fig. 1). There were no significant associations between seropositivity and occupation. TBEV antibody detection was higher in males (62.4%) versus females (36.8%) (p<0.001). Differential IFAT titrations against DENV and JEV were performed on a subset of 60 human sera, including 30 TBEV seropositives and 30 TBEV seronegatives. Of the 30 TBEV positive samples, 12 had equivalent titers to JEV and DENV, 17 had higher relative titers to JEV than DENV, and only one had a higher titer to TBEV than to the JEV and DENV.
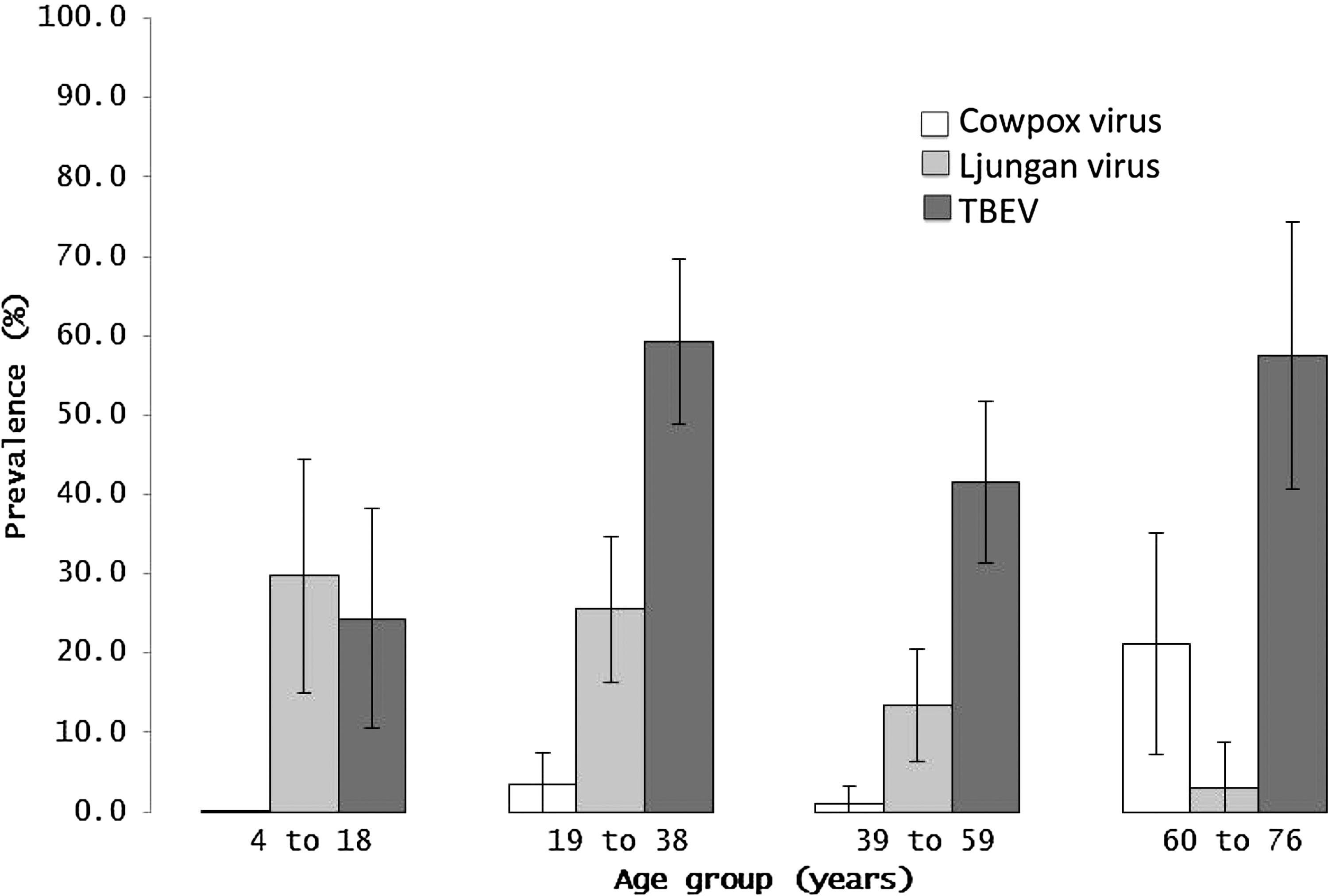
Human seroprevalence to flavivirus (TBEV), parechovirus (Ljungan virus), and poxvirus (cowpox virus) stratified by age group. Cohort age groups were 4–18 years (n=37), 19–38 years (n=86), 39–59 years (n=89), and 60–76 years (n=33). Bars indicate 95% confidence intervals.
Values within parentheses are percentages.
Associations between seroprevalence and animal contact
Of the 245 participants, the most frequently reported animal contact (>10 h per week) involved pigs (174/245, 71%), poultry (175/245, 71%), and dogs (149/245, 61%) (Table 4). Contact with ducks, muscovy ducks, and cats was also common (36%, 28%, and 37%, respectively). Twenty people reported contact with rats, most commonly through work activities, food preparation, or direct consumption. Three cohort members reported maintaining bat roosts to produce guano for fertilizer. Risk factor analysis involved stratifying people as exposed or unexposed to the above-mentioned animal species and examining associations with seroprevalence to the five virus groups. Overall, people with bat contact had higher seropositivity to cowpox virus (relative risk [RR] 8.07, 95% confidence interval [CI] 1.46, 44.67); however, the sample size was extremely small (n=3). Rat contact was associated with an elevated risk of LCMV seropositivity (RR 11.25, 95% CI 0.73, 173.16). However, except for the relationship between bat contact and cowpox seropositivity (p<0.001), none of the above-mentioned associations reached statistical significance.
TBEV, tick-borne encephalitis virus; pos., positive; RR, relative risk; CI, confidence interval; DOBV, Dobrava virus; SEOV, Seoul virus; LCMV, lymphocytic choriomeningitis virus.
Discussion
Rodents are known to host a wide spectrum of human viral and bacterial pathogens; however, investigations of rodent-borne diseases are relatively rare, and this diverse group of infections is not frequently prioritized by biomedical or public health research communities. Hantaviruses (family Bunyaviridae) are arguably the rodent-borne viruses of greatest public health signficance in Asia, where they are associated with an estimated 150,000–200,000 cases of hemorrhagic fever with renal syndrome (HFRS) each year (Bi et al. 2008). The hantavirus genus comprises at least 24 recognized virus species, several of which are pathogenic to humans. Although hantaviruses are known to circulate in Vietnam, few clinical cases have been reported (Huong et al. 2010), and the relative scope of the associated disease burden remains poorly characterized.
Our study indicated an overall 8.3% hantavirus seroprevalence among the five rodent species tested from southern Vietnam. This prevalence is consistent with previous reports from Vietnam (approximately 2.8% and 8.6% seropositivity in rodents from the Ho Chi Minh City and northern provinces, respectively) (Truong et al. 2009, Luan et al. 2012). We found hantavirus antibodies in all rat species with the exception of R. exulans. Despite the relatively high seroprevalence for hantaviruses, and the current understanding that most reservoir hosts develop chronic persistent infections (Easterbrook et al. 2007, Vaheri et al. 2013), only one rat sample (R. argentiventer) was confirmed to be hantavirus positive, with conventional PCR indicating SEOV. R. argentiventer has not previously been identified as a host of hantaviruses, and the worldwide distribution of SEOV has been almost exclusively associated with R. norvegicus (Lin et al. 2012).
Murine hantaviruses are believed to cross-react strongly; however, to ensure detection of a broad spectrum of potential hantavirus antibodies, we used two types of antigens (SEOV and DOBV), representing rat or mouse hosts, respectively. (It should be noted, however, that hosts of DOBV, Apodemus mice, are not found in Vietnam.) Interestingly, most of the rodent sera revealed variable reactivity to DOBV and SEOV viruses, with only five of 19 (26.3%) sera testing positive for both, suggesting that multiple hantaviruses may be circulating in the area. It is known from other studies in southeast Asia that B. indica carries Thailand hantavirus (Hugot et al. 2006), R. tanezumi carries Serang virus (Plyusnina et al. 2009), and R. norvegicus carries SEOV.
Regarding the human cohort, we detected seropositivity to both SEOV (7/245, 2.9%) and DOBV (4/245, 1.6%), again suggesting that variable hantaviruses may circulate. The results are consistent with previous low seroprevalence rates detected from humans in northern Vietnam (Truong et al. 2009). Risk factor analysis did not suggest any significant associations between hantavirus seroprevelance and contact patterns with any particular animal species. Our results indicate that one or more hantavirus species is endemic in common rat species of Vietnam; however, despite relatively intense and common human exposures to rats, human seroconversions to hantavirus are not common.
The Flavivirus genus (family Flaviviridae) comprises approximately 70 virus species, including a number of important human pathogens and a clade of species presumed to be transmitted primarily by rodents, other small mammals, and bats (the so-called “no known vector” group) (Kuno et al. 1998). Within southeast Asia, the only two recognized human clinical diseases caused by flaviviruses are dengue and Japanese encephalitis, and infections with tick-borne encephalitis (TBEV) have not been reported. Although Ixodes spp. ticks have been described in Vietnam (Kolonin et al. 1995), direct observation of ectoparasites on rats suggests little prevalence of ticks on rats of the Mekong Delta region.
Our IFAT screening results indicated overall TBEV seropositivity in humans of 47% and lesser rates in the rodents of 5.4%. Given that flaviviruses induce life-long immunity, that dengue and JEV are both highly endemic in southern Vietnam (e.g., approximately 53% dengue seroconversion in humans by age 7, increasing to 88% by age 13) (Thai et al. 2005), and that JEV vaccination is now widely administered throughout the country (Yen et al. 2010), our observed TBEV immunological reactivity in the humans was not at all surprising and most likely reflects the notorious cross-reactivity of flaviviruses (Roehrig, 1986, Basile et al. 2013). In contrast, the rodent seroreactivity to TBEV (15/275, 5.4%) was more intriguing. Rodent TBEV positives are much less likely to be due to dengue, because the principal dengue mosquito vector, Aedes aegypti, is almost exclusively anthropophilic (Harrington et al. 2001). Rodents have never been suggested to play a role in JEV transmission; however, given the high levels of endemicity in farms and periurban areas of the Mekong (Lindahl et al. 2013), the observed seroreactivity in rodents could be explained by spillover infections from avian reservoirs vectored by various Culex species. Our failure to identify viral genomes in the liver tissues of any of the 275 rodents suggests that the observed seroconversions were caused by transient infections, and/or the tropism of the virus is nonhepatotropic. The present results leave open the possibility that non-DENV, non-JEV flaviviruses circulate endemically within rodents of the Mekong, perhaps involving tick vectors, as has been found in Malaysia (Smith 1956) and Thailand (Bancroft et al. 1976).
None of remaining virus groups included in our survey, namely cowpox virus, arenaviruses, and rodent parechoviruses, showed evidence of high seroreactivity in the rodents. Cowpox viruses belong to the genus Orthopoxvirus that is immunologically cross-reactive and cross-protective, and cowpox infection in rodents has been reported across Europe from a wide range of rodent species, although R. norvegicus and bank voles are thought to be the principal reservoirs (Essbauer et al. 2010). In the present study, poxvirus antibody detection in humans was 4.5%, and was notably higher among people over 60, which may be attributable to prior smallpox vaccination. In contrast, poxvirus seropositivity was only 1.1% in rodents; the positive rodents were B. indica and R. tanezumi. Notably, we found a significant association between presence of pox antibody among cohort members and exposure to bats (RR 8.07, 95% CI 1.46, 44.67; p<0.05). Seroprevalence to LCMV was much lower (human, 0.8%; rodent, 0.4%) than has been observed from Thailand, where overall LCMV seroprevalence among rodents in one study was 13.3%, mostly in Bandicota savilei (35.7%) and R. norvegicus (31.5%) (Nitatpattana et al. 2000).
Ljungan virus is a rodent-borne parechovirus and suspected human pathogen (Klitz and Niklasson 2013), and although relatively high seroprevalence in humans has been documented from northern Europe (Jääskeläinen et al. 2013), to date there have been no studies of Ljungan virus in Asia. None of the Vietnam rats were seroreactive to Ljungan virus; the lack of Ljungan virus seropositives among the Vietnamese rats may not be surprising, because Ljungan virus in Europe has previously been associated primarily with bank or field voles (members of the Arvicolinae) rather than Rattus spp. However, as many as 18.8% (46/245) of human cohort members tested positive to Ljungan virus antibodies. The human Ljungan seroreactivity did not appear to be linked to any particular animal exposures. The age distribution of seropositives declined with age, as has been observed from surveys of human parechoviruses (HPeV) in Europe (Benschop et al. 2008). This pattern of declining immunity with age may indicate human-to-human transmission via fecal–oral transmission, as is typical for the known HPeVs , rather than exposures to a zoonotic reservoir. It should be noted, however, that current knowledge of Ljungan immunology suggests that the virus does not cross-react with HPeV 1, 2, or 4–6 (Jääskeläinen et al. 2013). Our results provide intriguing suggestions of frequent human exposures to an as-yet-unidentified parechovirus.
In summary, our study is the first to document the common rodent species traded in wet markets and sold for human consumption in Vietnam (primarily R. argentiventer and B. indica). Our results confirm low-to-moderate levels of endemic circulation of hantaviruses in the region and suggest possible transmission of as yet unidentified rodent flavivirus and human transmission of a Ljungan-like parechovirus. Further explorations of pathogen diversity in rodent reservoirs using larger sample sizes and more sensitive molecular screening techniques (e.g., alternative degenerate primers of genus-specific studies, or metagenomic and deep sequencing approaches) will no doubt uncover substantial additional viral diversity. Given the scale and scope of rodent trading in the Mekong, and indications that rodent consumption may also be increasing in northern Vietnam, establishing the epidemiological relevance of human exposures to rat viruses is an important public health objective.
Footnotes
Acknowledgments
The authors would like to thank staff from the Sub-Department of Animal health of Dong Thap for providing logistic assistance in carrying out the study. Work was partly funded by the Vietnam Initiative on Zoonotic Infections (VIZIONS), part of the Wellcome Trust Major Overseas Programme (United Kingdom), and the Li-Ka-Shing Foundation, Sigrid Jusélius Foundation, and Academy of Finland.
Author Disclosure Statement
No competing financial interests exist.