
Abstract
Children with a cleft in the soft palate have difficulties with speech, swallowing, and sucking. These patients are unable to separate the nasal from the oral cavity leading to air loss during speech. Although surgical repair ameliorates soft palate function by joining the clefted muscles of the soft palate, optimal function is often not achieved. The regeneration of muscles in the soft palate after surgery is hampered because of (1) their low intrinsic regenerative capacity, (2) the muscle properties related to clefting, and (3) the development of fibrosis. Adjuvant strategies based on tissue engineering may improve the outcome after surgery by approaching these specific issues. Therefore, this review will discuss myogenesis in the noncleft and cleft palate, the characteristics of soft palate muscles, and the process of muscle regeneration. Finally, novel therapeutic strategies based on tissue engineering to improve soft palate function after surgical repair are presented.
Introduction
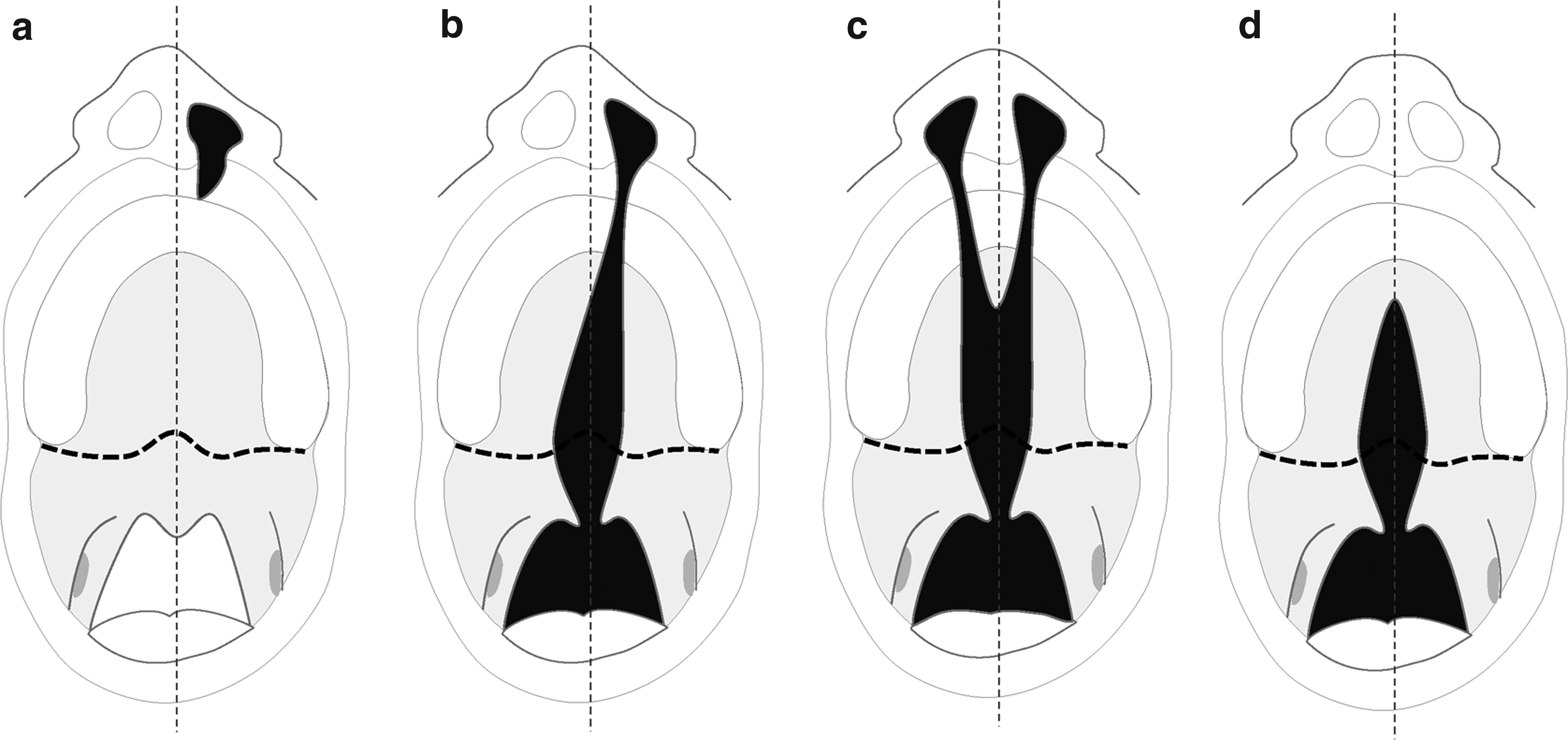
Cleft lip and/or palate.
Children with a cleft in the soft palate have difficulties with speech, swallowing, and sucking. These patients are unable to separate the nasal from the oral cavity; a phenomenon known as velopharyngeal dysfunction. Surgical repair is required to close the defect and to reconstruct the muscle sling of the m. levator veli palatini (LVP), the major muscle of the soft palate 4 (Fig. 2). It ensures optimal function and normal speech development.5,6 However, about 10% to 30% of the treated patients remain unable to achieve adequate velopharyngeal function.7,8 This results in hypernasal speech that often requires additional surgical corrections. Diverse factors are attributed to suboptimal repair such as age at the time of the surgery, skills and experience of the surgeon, type and extension of the cleft, fibrosis, damage of the motor and proprioceptive innervation, and abnormal fiber type distribution.7,9–12
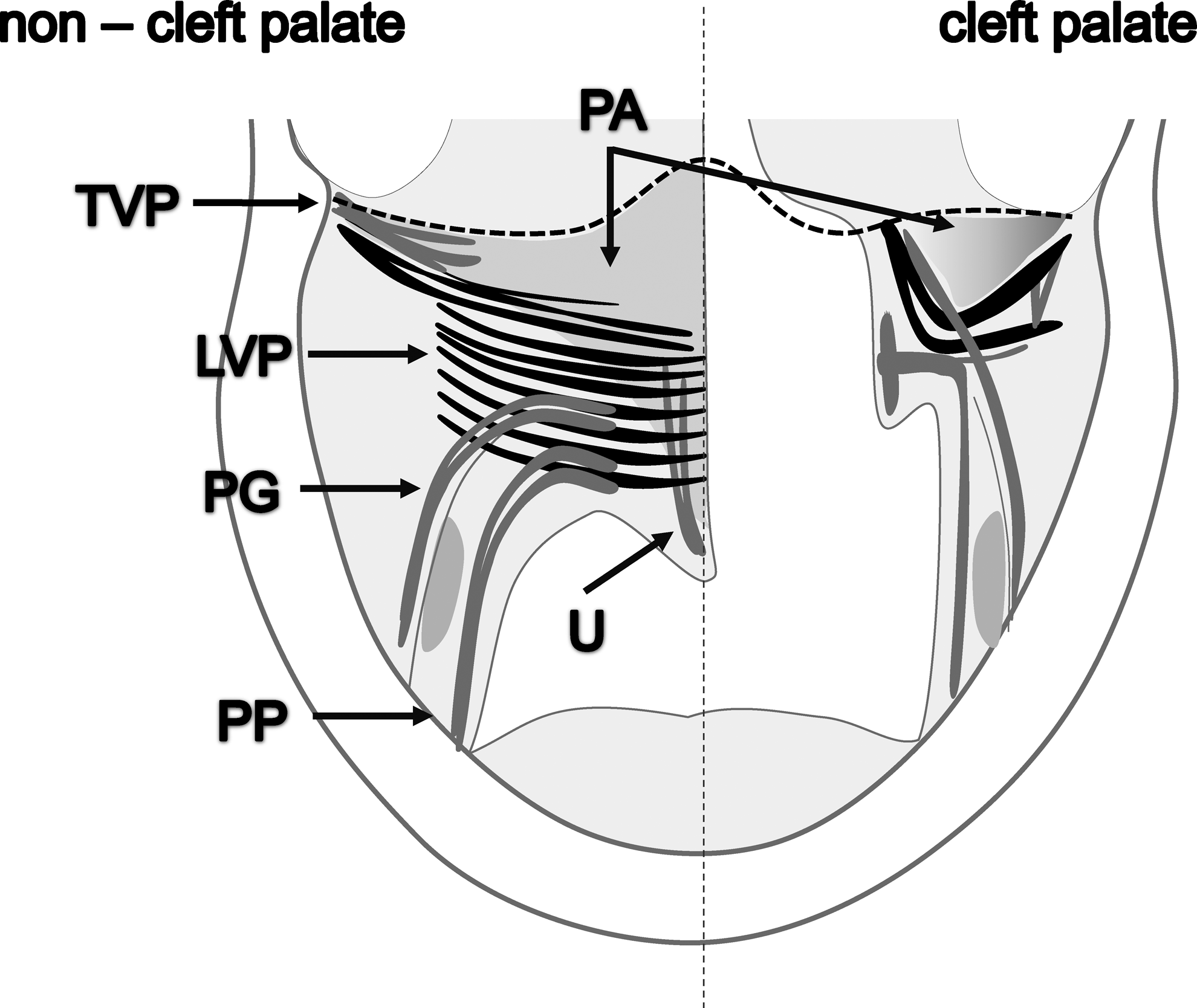
Muscles of the soft palate. m. tensor veli palatini (TVP), m. levator veli palatini (LVP), the major muscle of the soft palate (black), m. palatopharyngeus (PP), m. palatoglossus (PG), m. uvulae (U), and palatal aponeurosis (PA). The dashed line indicates the posterior border of the hard palate. Notice the abnormal insertion of the soft palate muscles into the posterior border of the hard palate at the cleft side. In this area disorganized myofibers are present. Cleft palate muscles are limited to isometric contractions and often underdeveloped.
This review will highlight the myogenesis and characteristics of the soft palate muscles in the noncleft and cleft palate, and the process of muscle regeneration. We postulate that muscle healing and function after soft palate repair can be improved by adjuvant strategies from the field of regenerative medicine. How these novel adjuvant strategies based on tissue engineering could improve muscle healing and prevent velopharyngeal dysfunction after surgical repair will be discussed.
Soft Palate Myogenesis
The muscles of the soft palate and those controlling jaw movement, facial expression, and pharyngeal (throat) and laryngeal (voice box) function are known as branchiomeric muscles.13,14 The formation of the soft palate muscles starts already early in embryonic development and yields five pairs of muscles: (1) the tensor veli palatini (TVP), (2) the LVP, (3) the palatopharyngeus (PP), (4) the palatoglossus (PG), and 5) the uvulae (U) (Fig. 2). Mesenchymal condensations of the individual soft palate muscles emerge sequentially, starting from the sixth week of gestation in humans, with the TVP appearing first and the U last. Their full morphological development is completed after the 17th week of intrauterine life. The hard palate is already fused at this stage. The masticatory muscles and the TVP develop at the same time, and are followed by the morphogenesis of the LVP, PP, PG, and U muscles, and by the palatal aponeurosis (PA).15,16 The development of the soft palate muscles in cleft patients is delayed compared with noncleft individuals. 17
Soft Palate Muscles in the Normal and Cleft Palate
The soft palate is formed by an interweaving of muscles from the skull base (TVP and LVP), tongue (PG), and pharynx (PP; Fig. 2). All muscles extend from nearby bony structures and are inserted into the PA located in the center of the soft palate (Fig. 2). Hence, all muscles have only one skeletal insertion, with the exception of the U, which is a completely intrinsic muscle of the soft palate without bony attachment. In the cleft palate, the muscles are attached to the posterior border of the hard palate (Fig. 2). The abnormal insertions of the muscles, in particular the LVP, in cleft palate patients prevent normal functioning. These muscles have two instead of one skeletal attachments limiting them to isometric contractions. 18 As a result, the cleft muscles pull the two halves of the soft palate in a superior and lateral direction, causing further widening of the cleft. 19 In addition, the LVP may become severely atrophic because of reduced function, and often has only half of the thickness found in healthy newborns. The LVP myofibers are also disorganized next to the cleft margin.16,17
Both slow and fast fibers are present in the soft palate muscles. Slow fibers are highly resistant to fatigue, with a low activation threshold, whereas fast fibers are more fatiguable, with a higher activation threshold. In noncleft individuals, the LVP contains predominantly slow fibers,20,21 whereas in cleft individuals it contains a higher proportion of fast fibers and a reduced capillary supply compared with normal LVP muscles.22–24 A fetal myosin heavy chain isoform is also present in a small number of fibers in all palatal muscles. In comparison with limb muscles, the individual and intramuscular variability in fiber diameter of the LVP is much larger. 21 The higher number of fast fibers in cleft patients may cause the increased fatigability of the LVP during speech, 23 which contributes to velopharyngeal dysfunction. 25 Further, fast fibers are more prone to contraction-induced injury.24,26,27
Muscle Regeneration in the Soft Palate
In general, muscles possess a strong ability to regenerate after injury. Satellite cells (SatCs) 28 are the primary muscle stem cells, and are responsible for postnatal muscle growth, maintenance, and repair. After injury, adjacent SatCs become activated and migrate to the site of injury, proliferate, differentiate, and fuse to form new myofibers or repair damaged ones.29,30 In addition, signaling molecules from infiltrating macrophages, injured myofibers, and the disrupted extracellular matrix participate in the regeneration process.29–31 The formation of scar tissue may prevent proper muscle regeneration.32–34 The majority of studies on muscle regeneration have been performed in limb muscles, while studies on head muscles are scarce. Muscles from the limbs and the head differ in several aspects. Skeletal muscles from the trunk and limbs are derived from the somites 35 during embryonic development, while most head muscles including those of the soft palate are derived from the branchial arches.36–38 The masseter muscle, a branchiomeric muscle, seems to contain less SatCs than limb muscle. 39 Freeze or crush injuries in the masseter muscle also regenerate much slower than similar injuries in limb muscles. 40 Moreover, much more fibrous connective tissue is formed in the damaged area. Proliferating SatCs from limb and head muscle also express a distinct profile of transcription factors. 39 In vitro, SatC-derived myoblasts from masseter proliferate more but differentiate later than those from limb muscle. Remarkably, both SatC populations are similarly able to regenerate limb muscle injuries after transplantation. 39 These observations support the crucial role of the SatC niche in muscle regeneration.
As mentioned, fast fibers are predominantly found in cleft palate muscles compared with normal soft palate muscles.22–24 In general, the percentage of SatCs in fast muscle fibers is significantly lower than in slow muscle fibers.41,42 This may further decrease the SatC number in cleft palate muscles. In addition, in vitro studies show that SatCs from fast muscle fibers proliferate less than those from slow muscle fibers. 39 In rodent models for muscle atrophy induced by disuse, SatCs decrease in number and display reduced function.43–45 Further, impaired macrophage recruitment to the site of injury and impaired macrophage function has been recently demonstrated in this muscle atrophy model. 46 A similar situation may exist in cleft palate muscles that are limited to isometric contractions and are therefore also atrophied. The accompanying poor capillary supply may further impair the regeneration process and promote fibrosis.18,19
Disorganized muscle fibers are found at the attachment of cleft palate muscles to the posterior border of the palatal bone.16,17 This possibly prevents further postnatal myofiber maturation by normal function. 47
In summary, the lower regenerative capacity of branchiomeric muscles in general, and the specific properties of cleft palate muscles may compromise muscle regeneration following surgical repair in cleft patients.
Soft Palate Muscles in Animal Models
Several mammalian species have been used for the study of the normal and clefted palate. The most frequently used animal models are goats, mice, and rats.48–53 Up to now, tissue engineering studies in the soft palate have not been performed. Therefore, the selection of an appropriate animal model for the study of the regeneration of the soft palate muscles is highly important. The use of an existing congenital cleft palate model in the goat51,54–56 is not feasible due to the high costs that limits group size. Cleft palate models in knockout mice are generally limited to embryological studies due to high mortality after birth, and lack of reproducibility of the induced clefts.57–60 However, the widespread use of rodents for fundamental and preclinical research ensures the availability of extensive tools and markers. In addition, their low cost and ease of handling make them the most suitable model for the development of soft palate muscle engineering techniques.
Similar to humans, the soft palate muscles in rodents possess a mixture of slow and fast fibers, 61 and play an important role in swallowing and respiration.61,62 In addition, the soft palate muscles in cleft animals also run parallel to the margins of the cleft, and disorganized myofibers are present. 60 In contrast to humans, 17 the development of the palatal muscles in cleft palate mice is not delayed. 63
The advantage of the rat over the mouse is its larger body size favoring surgical manipulation of the soft palate. This makes the rat the most suitable animal for experimental research on muscle regeneration of the soft palate.
Tissue Engineering to Improve the Outcome of Surgery
As mentioned in the previous sections, several biological factors have been attributed to functional impairment of the soft palate after surgical correction (Fig. 3): (1) Low intrinsic regenerative capacity of soft palate muscles. As shown, branchiomeric muscles including those of the soft palate, have a low number of SatCs with a low rate of differentiation compared with limb muscles, which may contribute to their poor regenerative capacity. (2) Muscle properties related to clefting. Clefted soft palate muscles contain a lower number of myofibers and are highly disorganized close to the cleft margin. This impedes the function of the reconstructed muscle. (3) The development of fibrosis. The main harmful effect of surgery is the development of muscle fibrosis, which impairs the regeneration of muscle fibers. Strategies based on tissue engineering may ameliorate the functional outcome of surgical repair by approaching these three main issues.
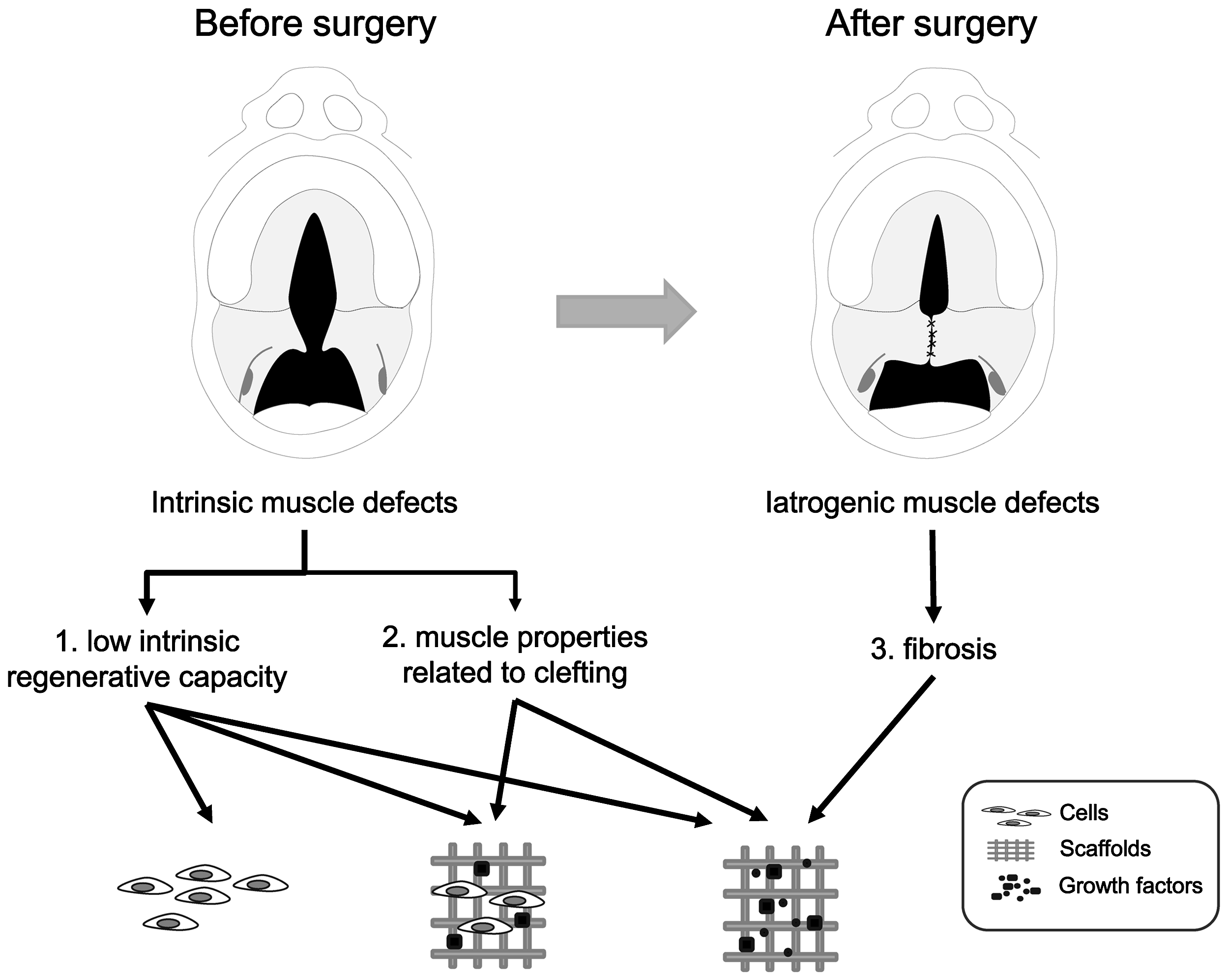
Muscle problems related to clefting. Before surgery: (1) Low intrinsic regenerative capacity: less number of satellite cells with a low rate of differentiation, (2) Muscle properties related to clefting: clefted soft palate muscles contain lesser number of myofibers and are highly disorganized. After surgery: (3) Fibrosis. Adjuvant strategies based on tissue engineering using (stem) cells, growth factors, and scaffolds may improve the outcome after surgery.
Another factor that may influence the regeneration of palatal muscles is age. In general, the number of SatCs in growing postnatal muscle is about 30%–35% of all nuclei.64–66 In adulthood, only 1–4% of all nuclei belong to the SatC population. 67 Surgical repair of the soft palate generally takes place in early childhood (6–36 months of age). After birth, the soft palate muscles are still immature. A large number of SatCs are present to fulfil the demands of growth. 68 In contrast, newborn SatCs seem to engraft less efficiently than their adult counterparts. 69 This should be taken into account when developing tissue engineering strategies for soft palate muscles.
Improving the intrinsic regenerative properties of the soft palate muscles
Isolated SatCs may be used for cell-based therapy to improve the regenerative capacity of the soft palate muscles. It has been demonstrated that transplanted SatCs are able to form new myofibers and to self–renew. 70 This approach has already been used for the treatment of a variety of diseases such as muscular dystrophy 71 and myocardial infarction.72–74 Unfortunately, the success of transplantation of SatCs and myoblasts has only been limited. After isolation, SatCs seem to lose their regenerative capacity in culture. In addition, their survival and migration to the site of injury after injection is limited.75,76
Mesenchymal and embryonic stem cells can also differentiate into skeletal muscle.77–79 However, only a small fraction of cultured mesenchymal stem cells undergo fusion into myofibers 80 suggesting that these cells may not be an appropriate cell type for therapeutic use. 81 Embryonic stem cells have a large potential for cell-based therapies for a wide range of diseases.82,83 These cells may offer additional advantages such as their larger replicative potential, and the possibility to perform gene targeting. 84 However, embryonic stem cell-derived myogenic populations with possible therapeutic value have not been found up to now.
Alternatively, other nonmuscle stem cells such as mesoangioblasts and pericytes also have myogenic potential.85,86 Both mesoangioblasts and pericytes can be delivered systemically because of their ability to cross the endothelium. This approach was successful for cell therapy in muscular dystrophy.86,87 Nevertheless, the exact ability of nonmuscle stem cells for muscle regeneration and their possible use in cell therapy remain to be elucidated.88–91
As an alternative approach, the recruitment, proliferation, and differentiation of SatCs already present in the soft palate muscle may be stimulated by specific growth factors. As mentioned, muscle growth and regeneration is controlled by locally-produced growth factors, which stimulate SatCs such as insulin-like growth factor 1 (IGF-1) and fibroblast growth factor 2 (FGF-2). IGF-1 favors muscle regeneration by increasing the rate of SatC proliferation and the formation of myotubes.92–96 Similarly, FGF-2 stimulates muscle regeneration in vitro and in vivo by promoting the proliferation of myoblasts after injury.97–100 Moreover, it facilitates the recruitment and proliferation of SatCs 101 and promotes angiogenesis.102,103 However, when growth factors are injected in vivo, they rapidly lose their biological activity due to diffusion and/or enzymatic inactivation. 104 Because of this, growth factors and attachment motifs have been incorporated into biodegradable scaffolds for controlled release. 105 With microspheres prepared from materials such as poly-lactic-co-glycolic acid, the release of such factors can be controlled.106–108 In addition, fibrin and gelatin hydrogels can give a sustained release of (growth) factors.109–112 These new techniques may improve the delivery of factors to the site of injury and stimulate muscle regeneration.
Although cell-based therapy seems to be promising for the treatment of genetic muscle defects, it is probably not a feasible option for cleft palate because it is a completely different type of condition. For example, in degenerative diseases like Duchenne muscular dystrophy, multiple cycles of muscle degeneration and regeneration deplete the SatC population. 113 In contrast, in cleft palate, muscle SatCs are preserved. In summary, growth factors and cytokines in a suitable delivery system may stimulate proliferation and differentiation of resident SatCs in the soft palate muscles after surgical repair.
Correcting cleft-associated muscle abnormalities
Muscle is a highly organized structure consisting of long, parallel multinucleated myotubes to generate sufficient force for contraction. 114 Therefore, the random myofiber orientation in cleft palate muscles impedes normal function after surgical repair. The reduced function limits the switch from a fast to a slow myofiber type.12,24 This may explain the increased muscle fatigability observed in patients with velopharyngeal dysfunction. 23
Proper alignment of regenerating myotubes may be achieved with scaffolds of specific design and surface topography. 115 Myoblasts are known to respond to the surrounding topography, 116 which leads to directional cell growth. 115 Hence, different techniques have been described to control cellular alignment on micro- and nanostructured surfaces produced either by chemical or topographic patterning. These include electrospinning, 117 photolithography, 118 and electron beam lithography. 119
Aligned nanofiber meshes can be fabricated from biocompatible polymers such as poly-lactide or poly-glycolide-co lactide via electrospinning. 119 Muscle cells cultured in these meshes show alignment and migration and a contractile phenotype.120,121 However, up to now the possibilities to control the mechanical properties of these polymer meshes is limited. 120 In addition, slowly degrading polymers persist in the regenerated tissue for a long time. Since the repair of the soft palate is generally performed in children, later growth of the soft palate must be accommodated. Therefore, the use of rapidly degrading polymers seems to be more suitable. Recently, it was reported that growth factors can be printed onto sub-micron polystyrene fibers.122,123 A combination of a suitable scaffold with growth factors and cytokines (e.g., stromal cell-derived factor–1α) might favor migration, 124 differentiation of resident SatCs, and induce myofiber alignment.
Particularly in patients with wider clefts, the amount of soft palate tissue is limited, which causes tension at the junction of the hard and soft palate after surgical repair. It has been proposed that scarring affects the length of the soft palate in anteroposterior direction, 125 which depends on the specific technique used for closure. 126 Alternatively, scarring may contribute to a narrowing of the residual cleft after two-stage palatoplasty.126–128 The site most likely to fistulize is at the junction of the hard and soft palate. 129 Fistulae usually have been associated with the type of cleft, technique and timing of repair, and the experience of the surgeon. 130 Substitutes as decellularized dermal allograft have been successfully used to close wide defects while preventing fistulae formation. 131 The use of scaffolds in the soft palate muscles allows a defined architecture to guide cell growth and development.
Preventing secondary effects of surgery: fibrosis
The deposition of excess ECM during regeneration may lead to fibrosis and impaired muscle function. A key factor in fibrosis is transforming growth factor-beta 1 (TGF-β1).132,133 It stimulates the synthesis of collagen and other extracellular matrix components and promotes myofibroblast formation. 134 Myostatin, another member of the TGF-β family, inhibits muscle regeneration by the inhibition of SatC and myoblast proliferation.135–138 In this way, TGF-β1 and myostatin synergistically amplify the fibrotic process after injury. In contrast, myostatin knockout mice develop less fibrosis and display improved skeletal muscle regeneration compared with wild-type mice. 139
The inhibition of TGF-β1 and myostatin may prevent fibrous scar formation and improve muscle healing after injury. 139 Decorin is a member of the small leucine-rich proteoglycan family, and it reduces both TGF-β1 and myostatin activity.139–142 It thereby enhances the proliferation and differentiation of myogenic cells, 143 and prevents fibrosis. 144 In addition, decorin upregulates the expression of a variety of myogenic markers including Myf5, Myf6, MyoD, and myogenin in vitro. 145 Hepatocyte growth factor increases decorin production by fibroblasts. 146 Furthermore, small signaling molecules such as nitric oxide down regulate TGF-β1 activity. 147 However, the importance of nitric oxide in muscle regeneration must be further elucidated. In summary, the use of decorin and other factors that inhibit TGF-β1 and myostatin may prevent scar formation after surgical correction of the soft cleft palate. A disadvantage might be that anti-fibrotic therapy can diminish narrowing of the residual cleft after two-stage palatoplasty. Therefore, the long-term effects of novel therapies based on tissue engineering in such a complex anatomical environment is hard to predict. Further research should elucidate the final outcome of these new therapeutic approaches.
Conclusions
This review discusses the development and characteristics of the soft palate muscles in the noncleft and cleft palate and the process of muscle regeneration. Finally, tissue engineering strategies are proposed to improve muscle regeneration after the closure of the soft palate. The regeneration of muscles in the soft palate after surgery is hampered because of (1) their low intrinsic regenerative capacity, (2) muscle properties related to clefting, and (3) the development of fibrosis. We propose the following strategies based on tissue engineering, to improve the functional outcome after surgical repair.
To improve muscle regeneration (problem 1), growth factors and cytokines might be applied to the surgical wounds by incorporation into scaffolds or microspheres. The growth factors should enhance SatC recruitment from the adjacent muscle tissue and their proliferation and differentiation into new myofibers. This will enhance the intrinsic regeneration capacity of the soft palate muscles, and thereby increase the functional outcome of surgery in terms of number and maturation of myofibers.
Suitable scaffolds may also support the alignment of myofibers and compensate the limited amount of tissue in clefted muscles (problem 2). These scaffolds should offer an architecture that guides cell growth and myofiber formation. In terms of function, this approach will increase muscle volume and force generation by the soft palate muscles.
To prevent fibrosis (problem 3), TGF-β1 and myostatin activity can be inhibited by factors such as decorin. This will reduce the formation of fibrotic tissue and stimulate proliferation of myoblasts. The function of the soft palate muscles after repair will thus improve by allowing myofiber formation and alignment (approaches 1 and 2).
Wrapping up, scaffolds that give a controlled release of growth factors, guide the alignment of new myofibers, and prevent fibrosis might improve muscle regeneration after surgical repair of the cleft soft palate. Nevertheless, further research is required in order to develop these functionalized scaffolds.
Footnotes
Disclosure Statement
No competing financial interests exist.