
Abstract
Stem cell therapy provides a viable alternative treatment for degenerated or damaged tissue. Stem cells have been used either alone or in conjunction with an artificial scaffold. The latter provides a structural advantage by enabling the cells to thrive in three-dimensional (3D) settings, closely resembling the natural in vivo environments. Previously, we disclosed the development of a 3D scaffold made from cotton, which was conjugated with arginyl-glycyl-aspartic acid (RGD), to facilitate the growth and proliferation of mesenchymal stem cells (MSCs). This scaffold allowed the MSCs to adhere and proliferate without compromising their viability or their stem cell markers. A comprehensive analysis investigation of the molecular changes occurring in MSCs adhering to the cotton fibers will contribute to the advancement of therapy. The objective of this study is to analyze the molecular processes occurring in the growth of MSCs on a cotton-RGD conjugated-based scaffold by examining their gene expression profiles. To achieve this, we conducted an experiment where MSCs were seeded with and without the scaffold for a duration of 48 h. Subsequently, cells were collected for RNA extraction, cDNA synthesis, and whole-transcriptomic analysis performed on both populations. Our analysis revealed several upregulated and downregulated differently expressed genes in the MSCs adhering to the scaffold compared with the control cells. Through gene ontology analysis, we were able to identify enriched biological processes, molecular functions, pathways, and protein–protein interactions in these differentially expressed genes. Our data suggest that the scaffold may have the potential to enhance osteogenesis in the MSCs. Furthermore, our results indicate that the scaffold does not induce oxidative stress, inflammation, or aging in the MSCs. These findings provide valuable insights for the application of MSCs in tissue engineering and regenerative medicine.
Impact Statement
At present, there is a scarcity of research studies that examine the influence of a scaffold on cell transcriptomics. Our findings indicate that the RGD-cotton-conjugated scaffold yields favorable outcomes in terms of cell aging, oxidative stress, inflammation, and osteogenesis expressed factors. Cultivating cells within a three-dimensional structure will prove highly advantageous for scientists and clinicians who are striving to find treatments for degenerated tissues, particularly in areas where there is a lack of suitable options. The creation of this scaffold has the potential to pave the way for the production of clinical-scale material for transplantation in vivo.
Introduction
Stem cells have emerged as a promising treatment option for various ailments in the field of regenerative medicine. This innovative approach holds immense potential, particularly for degenerative diseases such as cartilage degeneration, where the regenerative abilities of cartilage cells are limited. 1 Mesenchymal stem cells (MSCs) represent a great therapeutic option for various diseases due to their regenerative potential. They are multipotent stem cells that differentiate into osteocytes, chondrocytes, adipocytes, and most cell types. 2 MSCs can be found in various tissues such as bone marrow, adipose tissue, dental pulp, cord blood, and placenta and are widely used in clinical trials, 3 either alone or in conjunction with a scaffold.
The implementation of a three-dimensional (3D) scaffold is of utmost importance in the development of tissue engineering due to the considerable advantages it offers over the monolayer culture system. 4 The scaffold’s 3D architecture offers a remarkable advantage by enabling cells to grow in a manner that closely mimics the in vivo environment. This allows them to form their extracellular matrix (ECM) and regenerate new tissue while maintaining their natural shape. In the pursuit of creating scaffolds for cell growth, both natural and synthetic polymers have been extensively investigated.4,5 Natural and synthetic polymers have been used for the development of scaffolds for cell growth. 6
Natural polymers such as chondroitin sulfate, hydrogels such as collagen and gelatin, hyaluronic acid, alginate, agarose, fibrin, polysaccharides, and sulfated-polysaccharide have been used, alone or in combination, to develop scaffolds for stem cells for tissue regeneration.6–10 They have been used for the growth and differentiation of stem cells into chondrocytes and other cell types. 11
Cellulose-based scaffolds have already been established to foster the growth and differentiation of MSCs into cartilage tissue. 12 They are composed of glucose and form a linear homosaccharide. Cotton fibers, which are made of 94% to 96% of cellulose, 13 have also been used to grow cancer stem cells. 14
Silk fibroin exhibits favorable mechanical properties, enhanced in vivo stability, and minimal immunogenicity. 6 Nevertheless, the cost of silk fibers is relatively higher, necessitating their combination with another material such as chondroitin sulfate. 6 On the contrary, cotton fibers present an appealing option for constructing a scaffold to facilitate stem cell growth. Cotton fibers possess low cytotoxicity, appropriate mechanical properties for scaffold development with diverse structures, and gradual degradation. 15 Moreover, cotton fibers are readily accessible, extensively utilized, and cost-effective in comparison with silk fibers. Most natural polymers possess the significant benefit of being both biocompatible and biodegradable Nevertheless, they undergo rapid degradation, which limits the time frame available for tissue regeneration and may trigger an immune response upon implantation.6,7,16
In addition, various synthetic polymers such as hydroxyapatite containing calcium, polyester, polylactic acid (PLA), poly (lactic-coglycolic acid)/poly (L-lactic acid), and triblock copolymer have undergone experimentation as scaffolds to facilitate the growth and differentiation of stem cells.4,17–19 Bioactive glass and calcium phosphates have also been used in bone tissue engineering. 20 A synthetic polymer polyethersulfone-based scaffold was recently developed for the growth of MSCs and osteogenic differentiation. 21 The dental implants’ surface, which underwent dual acid-etched (TEST) treatment, demonstrated the ability to enhance the adhesion and growth of dental pulp MSCs. In addition, it was found to improve their osteogenesis and angiogenesis, as reported by Diomede et al., in 2020. 22 Synthetic polymers possess the benefit of being mechanically, physically, and chemically compatible for the development of scaffolds. 16 They are simple to produce, can be manufactured in large quantities, and can be adjusted to suit various applications.6,21,23 However, the major drawback of synthetic polymers is the release of acids such as lactic acid, toxic to cell function causing inflammation. 4
To improve the attachment of cells to the scaffold, modifications are made to the scaffold surfaces by incorporating receptor adhesion motif peptides obtained from proteins. Among these peptides, the arginyl–glycyl–aspartic acid (RGD) sequence derived from fibronectin is the most commonly utilized.7,10 This peptide is ubiquitously expressed in most ECM proteins.7,10 It allows integrin αvβ3 expressing cells to bind to them. 24 Scaffolds that have been modified with RGD have been utilized for various cell types such as fibroblasts, endothelial cells, and stem cells. 10 Previous investigations have shown that the RGD-modified scaffold surface greatly enhances the expression of cell adhesion markers in comparison with the linear surface. 24
Nonetheless, there have been few investigations into the gene expression profile of cells that attach to the scaffold. Huang et al., for instance, observed an elevation in the expression of chondrogenic markers (collagen II/collagen I ratio), aggrecan, and Sox9 in the cells that adhere to the cellulose scaffold. 12 Furthermore, the upregulation of collagen II and aggrecan in MSCs was observed upon the introduction of sulfated cellulose scaffolds. The utilization of next-generation sequencing allows for the analysis of gene profiles in cells, which can be further explored in the field of tissue engineering. It is worth noting that our previous research demonstrated that the RGD-conjugated cotton 3D scaffold not only supports the healthy growth and proliferation of cells but also maintains the integrity of stem cell markers. 25
Hence, it is crucial to analyze alterations in the gene profile and any deviations that may impact the biological functionalities of the MSCs for more effectively planned clinical implementations. In the present investigation, a comprehensive transcriptomic analysis was conducted to compare MSCs that adhered to the 3D RGD-conjugated cotton scaffold with those that were seeded without a scaffold and cultivated in a 2-dimensional (2D) environment.
Materials and Methods
Material
Commercially available bone marrow MSCs (Stem Cell Technologies, Cambridge, Massachusetts, United States) were cultured in 10% fetal bovine serum qualified for MSC growth, and Dulbecco’s modified Eagle’s medium (DMEM) with low-glucose containing glutamine and penicillin–streptomycin (10,000 U/mL) (Life Technologies, Carlsbad, California, United States). For whole-transcriptome sequencing, RNA from cells adhering to the cotton fibers and control cells was isolated using an RNeasy extraction kit (Qiagen, Hilden, Germany). Complementary DNA was synthesized with the high-capacity cDNA reverse transcription kit (Applied Biosystems, Carlsbad, United States). DNA sequencing was run with Hiseq2000 Sequencer (Illumina, San Diego, California, United States).
Scaffold development
To covalently conjugate cotton fibers to RGD moieties, 300 mg (9.10−4 mol) of RGD peptide was dissolved in anhydrous dimethylformamide (DMF), followed by a dropwise addition of anhydrous chlorotrimethylsilane (9.10−3 mol), and then, the mixture was left to stir for 20 min. Next, dry cotton (10 g) that had been suspended in anhydrous DMF was added to the previously prepared solution and left to stir overnight at room temperature. The resulting mixture was then filtered and washed 10 times with distilled water to remove unbound RGD and all traces of the reagents previously used. The functionalized cotton was then dried in a desiccator for three days in a vacuum. 25
Cell culture
MSCs were revived and cultured in complete DMEM+ glutamine (with 10% serum and 1% penicillin–streptomycin). When cells reached 80% to 90% confluency, they were passaged as previously described. 25 Cells were then seeded at 0.5 × 106 with RGD-cotton-conjugated cotton fibers in 3 wells of 6-well plates and left for 48 h in an incubator at 37° and 5% CO2. The control wells contained cells only seeded at the same concentration without the scaffold. At 48 h, cells were detached from the cotton fibers by washing the fibers with medium. Cells were then collected in a 15 mL tube. Control cells were detached with trypsin and collected in a falcon tube. Both tubes were centrifuged at 1500 rpm for 5 min, the medium was removed, and cell pellets were collected for transcriptomic analysis.
Preparation of cDNA library and sequencing
Total RNA was isolated with the RNeasy kit using a cell pellet from each condition. RNA concentration and purity (A260/A280 > 1.8) were measured with Cubit. One microgram of RNA was used for whole transcriptomic analysis using Illumina Hiseq2000 Sequencer.
As previously described, for cDNA library, single-stranded cDNA was synthesized using 1 µg of RNA. Double-stranded cDNA was synthesized by primer extension using Ex Taq polymerase. Double-stranded cDNA was fragmented to 300–500 bp sizes by sonication to generate libraries for sequencing. Fragments were purified with AMPure beads (Agencourt, United States). Sequencing libraries were prepared from sheared cDNA using the TruSeq Paired-End Cluster Kit v2.0 and 200 cycle TruSeq SBS HS v2 kit generating 100 base pair reads. An electropherogram was run to check the size of the library and quality.
Gene Ontology and pathway analysis
The expression profile of 13675 genes was quality-checked using a box plot and principal component analysis (PCA) plot in both conditions. Differential expression of gene (DEG) analysis was done by NOIseq R package with no biological replicates. 26 We used the default (q = 0.8) as well as the most stringent q = 0.9 threshold. The DEG was visualized by a bar plot and mean-difference (MD) plot. Gene set enrichment analysis was done using R’s cluster profiler and enrichr packages with Kyoto Encyclopedia of Genes and Genomes (KEGG), Gene Ontology (GO) term enrichments, and protein–protein interaction (PPI).27,28 We analyzed gene set enrichment for up- and downregulated genes separately to understand the molecular mechanism of changes happening after the introduction of cotton.
Results
Cells adhered to the cotton fibers and formed aggregates 48 h after seeding (Fig. 1A and Fig. 1B). Cells kept their round natural shape in the 3D environment. Cells seeded in a classical monolayer adhered to the plastic dish (Fig. 1C) had a fibroblast-like phenotype in the 2D environment.

MSCs were seeded at 5 × 105 in 3 wells of a 6-well plate and left for 48 h.
Data quality
We first run a quality check on the data, to verify gene expression counts and experimental bias between samples. Experimental bias could be due to different techniques applied to the samples, different equipment, different reagents, and different handling processes. Our analysis showed that samples have been treated and analyzed under similar conditions. The expression values’ box plot shows that conditions are comparable and could be analyzed further (Fig. 2A).

Data quality check:
In the PCA (Fig. 2B), MSCs treated with a cotton scaffold formed a distinct cluster from the control cells. This shows that both conditions are distinct and could be separated based on gene expression values.
Differently expressed genes
We identified 67 DEGs, 31 of which are upregulated, while 36 genes are downregulated significantly in cotton condition based on log2 fold change (p < 0.05) and q = 0.8 (Fig. 3 and Fig. 4). Figure 3A is a dot plot showing gene expression in the cells adhering to the scaffold. The red dots show a significant difference compared with the control sample. The genes that are not significantly different from the control are represented in black (Fig. 3A). The DEGs are represented in the bar chart (Fig. 3B), in terms of their expression value in the cotton condition.

Differently expressed genes (DEG).
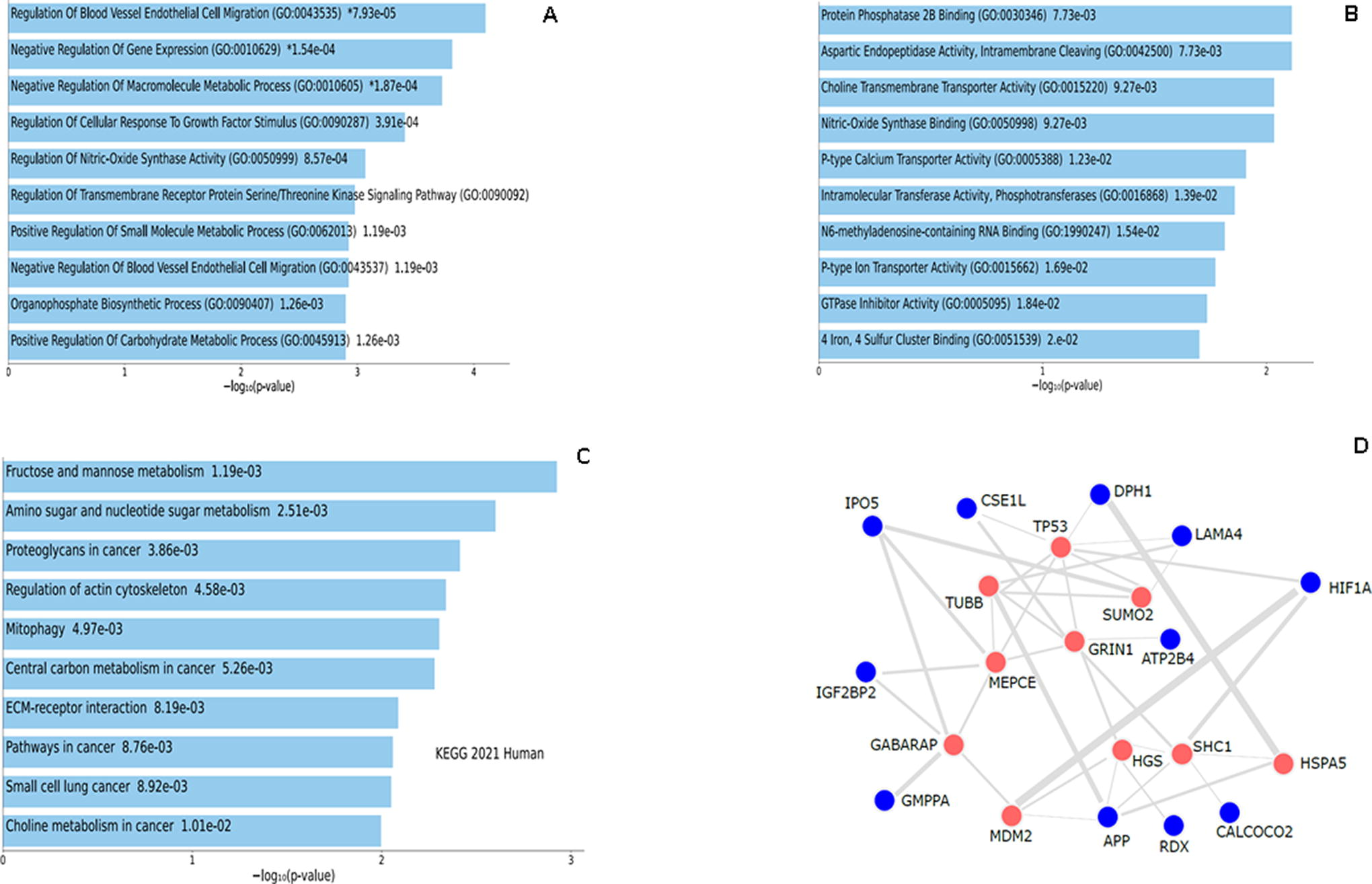
Gene enrichment analysis of the upregulated DEGs. GO term enrichment analysis showing the top 10 results from enrichment analysis of upregulated genes in
In the group of upregulated genes, we observed genes involved in cell adhesion: APP, ITGA3, and LAMA4,29–31 a gene involved in cell proliferation, angiogenesis, and cell differentiation, the fibroblast growth factor-2 (FGF2). 32 Gene with antiapoptotic roles (TXNDC5) and genes involved in cell cycle and growth (TSG101, ZNF207) were also increased.
On the contrary, the list of downregulated genes included mainly the cell cycle and growth genes (USP16, NUSAP1, ECT2). Furthermore, genes involved in cell proliferation (TRMT1, 33 MAP2K1, and MAP4K4), 34 cell differentiation (SUN1, ECT2, MAP2K1), neurogenesis (ECT2, SMARCE1), cell motility, migration, and angiogenesis (SPAG9), 35 and apoptosis (RABEP1, SRPRB, 36 MAP4K4), 34 were also significantly downregulated. The full list of DEGs is provided in Supplementary Table S1A for the upregulated genes and Supplementary Table S1B for the downregulated genes.
Functional enrichment analysis
The biological classification of the identified genes was further analyzed by the gene enrichment analysis.
Gene enrichment analysis: upregulated genes
GO term enrichment analysis was used to investigate the biological process and molecular functions of the upregulated genes. The bar charts in (Fig. 4A and Fig. 4B) show the biological process and molecular function, the number of genes associated with each category, and the statistical significance.
Our data revealed that upregulated genes are principally enriched in the regulation of gene expression, followed by the ECM organization and extracellular structure organization. Their molecular functions are mainly related to protein heterodimerization activity, fibroblast growth factor receptor, and choline transmembrane transporter activity. The top 10 most significant biological processes and molecular functions are represented in Figure 4A and 4B. The top 20 of these processes together with the cellular localization are shown in Supplementary Fig. S1. The top 20 corresponding most significantly increased (p < 0.05) GO term molecular functions are shown in Table 1. The top 20 biological processes for the upregulated genes are shown in Supplementary Table S2.
Representation of the Top 20 Gene Enrichment Based on GO Molecular Function of Upregulated Genes
The table represents the results of the GO molecular function of upregulated genes in cells with cotton.
Upregulated genes: KEGG pathway enrichment analysis
Pathway analysis showed that the upregulated genes were enriched in pathways in cancer, followed by proteoglycans in cancer, regulation of actin cytoskeleton, and PI3K-AKt signaling pathway. The upregulated genes were also enriched in other pathways such as the ECM-receptor interaction and focal adhesion (Fig. 4C). The list of the top 20 enriched pathways is provided in the Supplementary Figure S1D. Table 2 represents the top 20 gene enrichment terms based on “KEGG Pathways” showing the genes involved in each pathway and their significance.
Representation of the Top 20 Gene Enrichment Terms Based on “KEGG Pathways” in Upregulated Genes
The table shows gene enrichment terms based on “KEGG Pathways” in upregulated genes in cells with cotton.
PPI network construct for the upregulated DEG
The PPI network provided the molecular organization of the upregulated DEG. The upregulated DEGs are connected to a number of hub genes, involved in cell proliferation and differentiation (TP53, MDM2, MAPK, beta-tubulin, GABARAP),37–42 and genes involved in antiaging, cell protection, and anti-inflammatory roles (SUMO2, HGS, Grin1, HSPAS).43–47 DEGs were also connected to genes involved in cancer (SHC1, MEPCE) (Fig. 4D).48,49 The top 10 gene enrichment terms based on PPI hub genes are represented in Supplementary Table S3 showing the genes connected to each hub gene and the significance.
GO enrichment analysis: downregulated genes
GO enrichment analysis of the downregulated genes was also categorized into three functional groups (biological processes, molecular functions, and cellular localization). The results showed that downregulated genes are enriched in various biological processes mainly in the chromatin disassembly, nucleosome disassembly, protein-DNA complex disassembly, regulation of stress-activated mitogen-activated protein kinase (MAPK) cascade, positive regulation of transcription, and DNA-template (Fig. 5A). Their molecular functions are mainly associated with MAPK scaffold activity, nucleosomal DNA binding, nuclear receptor coactivator activator, ubiquitin-binding, creatine kinase activity, procollagen-proline 4 dioxygenase activity, JUNK kinase binding, procollagen proline dioxygenase activity, nuclear receptor binding, and aldehyde dehydrogenase (NAD+) activity (Fig. 5B). The bar chart of the full list of the top 20 biological processes, molecular functions, and cellular localization for the downregulated DEG is provided in Supplementary Figure S2. Table 3 shows the top 20 gene enrichment terms based on GO molecular function of the downregulated DEG. The top 20 biological processes in the downregulated DEG are shown in Supplementary Table S4.
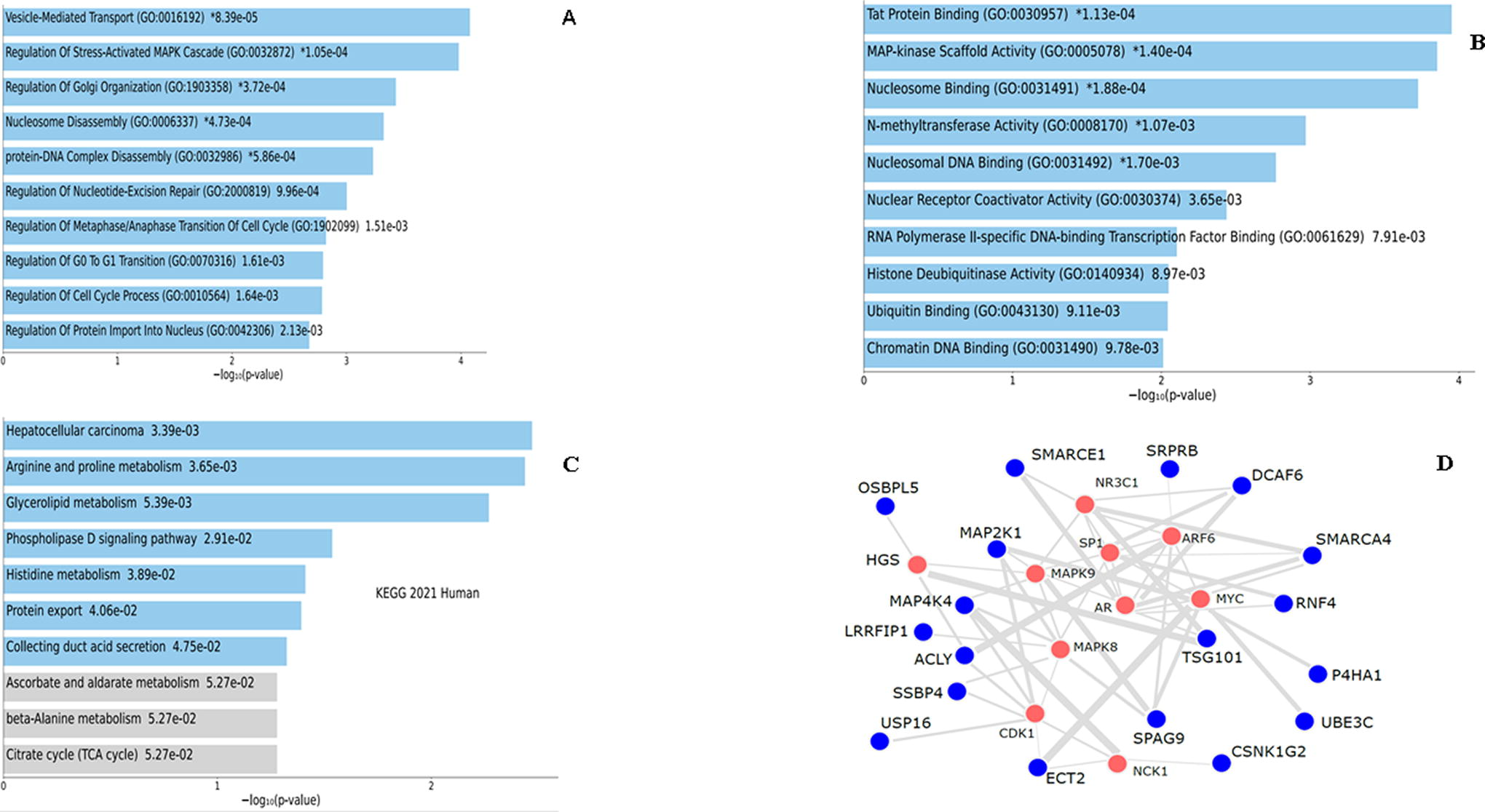
Gene enrichment analysis of the downregulated DEGs. GO term enrichment analysis showing the top 10 results from enrichment analysis of upregulated genes in
Representation of Top 20 Gene Enrichment Terms Based on “GO Molecular Function” in Downregulated Genes
The table shows gene enrichment terms based on “GO molecular function” in downregulated genes in cells with cotton.
KEGG pathway analysis showed that the downregulated genes were mainly enriched in metabolic pathways, arginine and proline metabolism, protein export, tryptophan metabolism, and pyruvate metabolism (Fig. 5C). The bar chart for the top 20 pathway analysis of the downregulated DEG is provided in the Supplementary Data (Fig. 6D). Table 4 represents the top 20 KEGG pathways in the downregulated DEG.

Differently expressed genes (DEGs).
Representation of Top 20 Gene Enrichment Terms Based on “KEGG Pathways” in the Downregulated Genes
The table shows gene enrichment terms based on “KEGG Pathways” in the downregulated genes in cells with cotton.
PPI network construct for the downregulated DEG
The downregulated DEGs were connected to several hub genes involved in cell proliferation, cell differentiation, and inflammatory response (NR3C1, MAPK8, NCK1),50–52 cell survival and apoptosis (SP1, HSPAS),43,53 cancer cell proliferation (AR, CDK1, SHC1), and stem cell proliferation and phenotype (cMYC).48,54–56 Similarly to the upregulated DEGs, the downregulated were also interacting with the hepatocyte growth factor (HGS) (Fig. 5D). Supplementary Table S5 shows the top 10 hub genes with the upregulated DEG connections showing the genes connected to each hub gene and the significance.
Analysis with a more stringent threshold
The threshold was increased to 0.9 to ensure maximum significant differences in the DEG. The MD plot shows the up- and downregulated genes in the cotton-seeded cells based on the log2 fold change. The stringent threshold revealed 18 upregulated genes and 15 downregulated genes (Fig. 6B). The list of the whole genes is provided in Supplementary Table S6 and Venn diagram (Fig. 7), which shows the common DEG between the default threshold (0.8) and the stringent threshold (0.9).
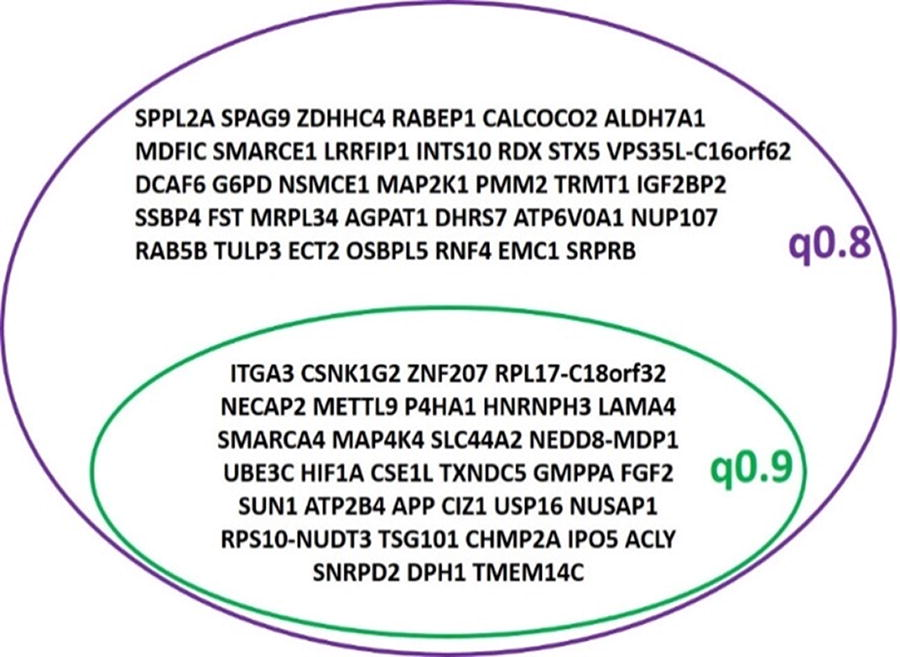
Venn diagram representing the DEGs with both thresholds applied. The DEGs that are consistent with a very stringent threshold of 0.9 are represented.
Genes involved in cell adhesion such as APP, ITGA3, and LAMA4 were significantly upregulated in the MSCs seeded on the cotton fibers. FGF2 and the genes with an antiapoptotic role (TXNDC5) and cell cycle and growth role (TSG101) were also significantly upregulated.
On the contrary, USP16 and NUSAP1 genes involved in cell cycle and growth were downregulated. Genes involved in cell proliferation, cell motility, cell migration, angiogenesis, and apoptosis (MAP4K4) 34 and genes involved in cell differentiation (SUN1) were significantly downregulated.
Discussion
Our study aimed at characterizing the gene expression profile of MSCs adhering to the 3D cotton-based scaffold. MSCs are heterogeneous cell populations exhibiting donor-to-donor, organ, and tissue differences, 57 and therefore, their characterization is of paramount importance for future clinical applications. Whole-genome sequencing helps us to understand how the 3D cotton-based scaffold can affect MSC gene expression, which has great implications in cell therapy.
Our previous data using gene and transcript-wise expressions for each population and calculating with custom Perl scripts, from the CLC Genomics workbench, identified 511 DEGs in the cotton condition with the analysis done without any filtration or reads per kilobase of transcript per million (RPKM) filtration or threshold. 25 In the present study, we used the R tool for differently expressed analysis with stringent threshold and gene enrichment analysis. Two thresholds were applied Q = 0.8 and Q = 0.9, which gave more significant results.
After normalizing the data, we identified 67 DEGs, 31 upregulated genes and 36 downregulated genes in the MSCs seeded with cotton. Adhesion markers were significantly increased in the MSCs seeded with cotton. APP, ITGA3, and LAMA4 were consistently upregulated in a more stringent threshold (0.9), which might reflect the effect of the RGD-cotton-conjugated fibers on the cells. As previously stated, RGD oligopeptides enhance cell adhesion to a scaffold.7,10,58 They bind to the VLA-5 integrin receptor expressed on the surface of the stem cells. 25
Analysis of DEG showed the upregulation of the TSG101 gene and downregulation of USP16 and NUSAP1 in the cotton condition, all of which are involved in regulating cell cycle and growth. Antiapoptotic gene (TXNDC5) and MAP4K4 were upregulated and downregulated, respectively. The cell differentiation marker FGF2 was upregulated, whereas SUN1 was downregulated in the cotton condition. These data indicated that there might be a balance in the expression of genes of the same functions keeping the cell homeostasis in terms of cell proliferation, apoptosis, and differentiation. The cells are capable of balancing between processes to achieve homeostasis. Regulation of gene expression is an important mechanism for all living cells, resulting from the interaction between specific proteins and specific DNA motifs to regulate gene expression. 59 There could be a balance between the upregulated and the downregulated genes with similar functions.
Our results showed increased expression of the ITGA together with an increase in the ECM-receptor, focal adhesion, and PI3K-Akt signaling pathways. Integrin alpha (ITGA) has a key role in ECM-receptor interaction and focal adhesion pathways in MSCs. 60 Our data are in correlation with previous reports indicating that RGD mediated increased expression of the adhesion markers in the cells. 24
It was previously shown that the overexpression of FGF2 in MSCs improved their survival. 61 We have previously shown that the cotton-based scaffold stimulates cell growth and maintains cell viability. 25 An increase in FGF2 expression in MSCs adhering to the cotton fibers could play a role in their viability and proliferation. The increase in FGF2 expression corresponds with the increase in the molecular function of fibroblast growth factor receptor binding in the MSCs adhering to the cotton fibers.
The MSCs adhering to the RGD-conjugated cotton fibers were shown to have enhanced expression levels of cell adhesion, cell differentiation, and ECM-related markers, which indicates that the stiffness of the cotton fibers may induce all these changes in the MSCs. It was previously suggested that the stiffness of the MSC microenvironment could induce the expression of adhesion cytoskeleton markers and ECM-related cytoskeleton markers, which in return would affect cell differentiation. 62 The increased biological process of the ECM organization in cells adhering to the cotton fibers indicates their enhanced adhesion potency to the fibers, as previously reported by Zeiger et al. (2012). 63
Our data revealed an increase in the molecular function of the protein phosphatase 2B binding, and an upregulation of the ECM pathway and the mammalian target of the rapamycin (mTOR) pathway. These data might suggest that MSCs adhering to cotton fiber are more likely to undergo osteogenic differentiation. It was previously reported that protein phosphatase 2B binding induces osteogenesis and inhibits adipogenesis in MSCs.64,65 Both ECM and the rapamycin (mTOR) signaling in addition to Jun N-terminal kinase were reported to induce osteogenesis in MSCs.66–68 The expression level of the Jun kinase was downregulated in the MSCs adhering to the cotton fibers. This might also suggest that cells have more potential to undergo osteogenesis. IGF2BP2 was recently reported to regulate osteogenesis in MSCs.69,70 The upregulation of the IGF2BP2 could indicate that MSCs are more likely to differentiate into osteocytes. Altogether, our data point toward the important role that the 3D cotton-based scaffold might play in the osteogenesis of MSCs.
It has been reported that MSCs are associated with a wide spectrum of signaling pathways involved in cancer induction and in cancer inhibition. 71 It is not clear whether the increase in the pathways in cancer in the MSCs adhering to cotton fibers reflects the cancer-promoting activity of these cells or not. Further analysis is warranted to investigate the cancer-promoting/inhibiting role of MSCs adhering to the cotton.
ECM’s role is to maintain the structure, the function of cells and tissues, cell proliferation and migration, and a role in wound healing via interaction with integrins.72,73 It was recently shown that the ECM-receptor interaction pathway leads to the reconstruction of the ECM in transplanted MSCs. 72 Proteoglycans are also involved in ECM organization. 74 MSCs adhering to cotton fibers had their ECM-receptor interaction and proteoglycan pathways upregulated indicating the stimulatory effect of cotton fibers on matrix formation and organization in the extracellular environment. The upregulation of the ECM pathway would protect the MSC structure and functions and regulate cell proliferation and migration. The focal adhesion PI3K-Akt-mTOR-signaling pathway has also been shown to affect ECM formation. It induces cell-ECM contact and cell–matrix adhesions. 75 Our findings indicated that the RGD-conjugated cotton fibers might have induced cell-ECM formation through upregulation of the focal adhesion PI3K-Akt-mTOR-signaling pathway. The results showed the increase in the ECM-receptor pathway, focal adhesion, and PI3K-AKt signaling would play an important role in the induction of cell-ECM formation protecting cell structure and functions and inducing cell proliferation.
Our result demonstrated the association between cotton fiber-adherent MSCs and increased glutathione metabolism. Lan et al., (2022) indicated that injected bone marrow MSCs would enhance stroke recovery through an increase in glutathione metabolism. The latter protects the cell from oxidative stress. 76 These data suggested that upregulation of the glutathione metabolism pathway would protect the MSCs from damage related to oxidative stress. The senescence protective effect of cotton fiber on MSCs is mediated through upregulation of the actin cytoskeleton pathway and a decrease in the metabolic pathways. The inhibition of the actin cytoskeleton and the metabolic dysfunction are believed to induce senescence in MSCs and in the MSCs of aged mice.77,78
The increase in the choline transmembrane transporter activity indicates that MSCs adhering to cotton fibers have anti-inflammatory roles. It was recently demonstrated that the MSCs might have anti-inflammatory roles through the cholinergic anti-inflammatory pathway and mediated via the choline transmembrane transporter. 79
The biological process and molecular functions of chromatin disassembly were shown to be downregulated in the cells adhering to the cotton fibers. The processes of chromatin disassembly and reassembly occur during DNA repair to maintain genomic integrity. 80 Both chromatin disassembly and reassembly are essential events for living cells but work oppositely. While the former induces activation of transcription, the latter inhibits transcription. 80 The upregulation of the biological process of gene regulation together with the downregulation of chromatin disassembly might work in a balanced manner to maintain gene integrity in the MSCs adhering to the cotton fibers.
Mitogen-activated protein kinases (MAPKs) are involved in a wide range of cellular activities, including the regulation of cell proliferation, differentiation, and inflammation These cellular activities are mediated by MAPK-specific scaffold proteins. 81 For instance, inflammatory cytokines activate MAPKs, which in return respond to the environment changes by stimulation of one of the MAPK groups such as c-Jun N-terminal kinase. 82 Downregulation of the JUN kinase in the MSCs adhering to cotton fibers correlates with a decrease in the MAPK biological processes. This finding may suggest that MSCs adhering to cotton fibers have no inflammatory effect in their environment and would indicate that MSCs cultured with cotton have great beneficial implications on cell transplantation.
The level of expression of the hub genes was not significantly different between the two conditions. However, the level of gene expression might not reflect the protein expression levels. PPI showed that the DEGs were interacting with genes involved in stem cell potency (TP53, Myc), osteogenesis via MDM2-regulated MAPK signaling pathway,37,38,40,56 and antiaging through the activation of SUMO2 and HGS.44–46 As described earlier, GO term analysis and pathway analysis suggest that MSCs might be protected from aging. Hub genes involved in adipogenesis (NR3C1) and chondrogenesis (SP1) might suggest that cells have to some extent the potential to differentiate into chondrocytes and adipocytes.50,83 This reflects the multipotentiality of the MSCs to differentiate into three lineages osteocytes, chondrocytes, and adipocytes.
Taken together, our data suggest that the adhesion of the MSCs to the RGD-conjugated cotton fiber scaffold promotes healthy MSCs by inhibiting oxidative stress and preventing cellular aging. Furthermore, the scaffold would exercise its protective function by enhancing the ECM formation, stimulating cell proliferation, and osteogenesis differentiation. Future studies focusing on the differentiation of the MSCs into the three different lineages, especially on the osteogenesis potential of the cells, will yield considerable benefits. The findings would have important implications for the evaluation of the molecular and cellular changes that might occur in MSCs grown with a scaffold intended for clinical applications.
Footnotes
Acknowledgment
The authors confirm that the article has not been previously published and that it is not under consideration elsewhere.
Availability of Data and Materials
All data generated and analyzed during this study are included in the main article and in the supplementary information. The data sets are available from the corresponding author.
Authors’ Contributions
H.A. generated the bioinformatics data, S.A. interpreted and analyzed the data, S.A., T.N., R.A., H.A., and F.M. wrote the discussion and proofread the article. The rest of the sections were drafted by S.A. The first draft of the article was written by S.A. All the authors read and approved the final article.
Disclosure Statement
The authors declare that they have no conflict of interest. The authors have no relevant financial or nonfinancial interests to disclose.
Funding Information
The work was funded by the King Abdullah International Medical Research Center (KAIMRC) (IRBC/1590).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.