
Abstract
Phenotype instability and premature hypertrophy prevent the use of human mesenchymal stromal cells (MSCs) for cartilage regeneration. Aim of this study was to investigate whether intermittent supplementation of parathyroid hormone-related protein (PTHrP), as opposed to constant treatment, can beneficially influence MSC chondrogenesis and to explore molecular mechanisms below catabolic and anabolic responses. Human MSCs subjected to chondrogenic induction in high-density culture received PTHrP(1–34), forskolin, dbcAMP, or PTHrP(7–34) either constantly or via 6-h pulses (three times weekly), before proteoglycan, collagen type II, and X deposition; gene expression; and alkaline phosphatase (ALP) activity were assessed. While constant application of PTHrP(1–34) suppressed chondrogenesis of MSCs, pulsed application significantly increased collagen type 2 (COL2A1) gene expression and the collagen type II, proteoglycan, and DNA content of pellets after 6 weeks. Collagen type 10 (COL10A1) gene expression was little affected but Indian hedgehog (IHH) expression and ALP activity were significantly downregulated by pulsed PTHrP. A faster response to PTHrP exposure was recorded for ALP activity over COL2A1 regulation, suggesting that signal duration is critical for catabolic versus anabolic reactions. Stimulation of cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA) signaling by forskolin reproduced major effects of both treatment modes, whereas application of PTHrP(7–34) capable of protein kinase C (PKC) signaling was ineffective. Pulsed PTHrP exposure of MSCs stimulated chondrogenesis and reduced endochondral differentiation apparently uncoupling chondrogenic matrix deposition from hypertrophic marker expression. cAMP/PKA was the major signaling pathway triggering the opposing effects of both treatment modes. Intermittent application of PTHrP represents an important novel means to improve chondrogenesis of MSCs and may be considered as a supporting clinical-treatment mode for MSC-based cartilage defect regeneration.
Introduction
M
Although common protocols for chondrogenic in vitro differentiation of MSCs evoke the expression of key molecules characteristic for articular cartilage, the natural course toward a stable articular chondrocyte phenotype is not recapitulated, as a process reminiscent of endochondral bone formation is triggered [7]. The hypertrophic phenotype of chondrocytes is mainly characterized by a strong increase in cell volume and the onset of expression of Indian hedgehog (IHH), collagen type X, and alkaline phosphatase (ALP), which is closely associated with matrix mineralization.
In the growth plate, the extent and velocity of hypertrophic differentiation are tightly controlled, most prominently by the parathyroid hormone-related protein (PTHrP)/IHH feedback loop [10,11]. The N-terminal region of PTHrP shares a high sequential and structural homology with parathyroid hormone (PTH) and signals via a common receptor, the G protein-coupled PTH/PTHrP receptor (PTH1R). Activation of the receptor triggers several different intracellular signaling pathways, including Gαs/adenylate cyclase/cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA) and Gαq/11/phospholipase Cβ/protein kinase C (PKC) [12]. More recently, signal transduction via ERK1/2 and β-arrestin has been reported as well [13]. The principal receptor-binding domain of PTHrP lies within amino acids (aa) 15–34, but the first 2 amino acids of PTH/PTHrP are essential for the activation of the cAMP/PKA-dependent pathway. The C-terminus of PTH/PTHrP(1–34), where PTH and PTHrP share structural rather than sequential homology, contains a domain necessary for PKC activation within aa 28–34. N-terminally truncated peptide analogs such as PTHrP(7–34) can bind to the receptor but do not elicit a cAMP/PKA response [14]. As PTHrP(7–34) can cell-type-dependently activate PKC [15,16], it may represent a useful tool to antagonize PTHrP(1–34)-related cAMP/PKA signaling while simultaneously maintaining PKC signaling.
There is evidence that PTHrP is operative also in postnatal articular cartilage [17] and might therefore be involved in the maintenance of the stable articular phenotype. Capable to modulate chondrocyte maturation and hypertrophy, PTHrP is, thus, a highly interesting candidate molecule to improve chondrogenesis of MSCs. However, constant application of PTHrP in chondrogenic medium suppressed chondrogenesis instead of selectively inhibiting hypertrophic features only [6,18]. Interestingly, PTH can exert opposing effects on the osteogenic differentiation of osteoblasts [19,20], rat MSCs [21], and on bone mass dependent on a constant versus intermittent application regime. Whereas constant application acted catabolically on bone, an intermittent application (eg, once daily) provided anabolic effects, resulting in a net increase in bone mass [22,23]. Similar effects were reported for PTHrP [24 –26] and first clinical trials are conducted to evaluate the benefit of intermittent PTHrP administration for the treatment of osteoporosis [27].
Although chondrocytes and osteoblasts share common progenitors, comparably little is known about a potential effect of intermittent application of PTH/PTHrP on chondrocytes in vitro and on cartilage tissue in vivo. Few studies reported on an improved osteochondral repair [28,29], stimulation of chondrogenesis in bone healing during callus formation [30], and inhibition of osteoarthritis progression in an in vitro and in vivo azacytidine-induced osteoarthritis model [31]. In contrast, an inhibition of cartilage formation was observed in a microfracture study after intermittent treatment with PTH [32].
How intermittent application of PTHrP affects in vitro chondrogenesis of MSCs remained so far unexplored. We here hypothesized that, opposite to constant application, intermittent application of PTHrP can exert beneficial effects on MSC chondrogenesis and investigated whether this may include stimulation of extracellular matrix deposition and suppression of hypertrophic chondrocyte development. Beyond, we intended to identify the signaling pathways relevant for constant versus intermittent PTHrP effects on human MSCs. Bone-marrow-derived MSCs were subjected to 6 weeks of chondrogenesis, receiving either constant or intermittent PTHrP(1–34) or chondrogenic medium alone. Markers for chondrogenesis and hypertrophy were quantitatively evaluated on the mRNA and protein levels and the contribution of different signaling pathways was explored by treatment with forskolin, dbcAMP, and PTHrP(7–34).
Materials and Methods
Isolation and expansion of MSCs
MSCs were isolated from fresh bone marrow aspirates obtained from 24 patients undergoing total-hip replacement by fractionation on a Ficoll-Paque Plus density gradient (GE Healthcare Europe), and the low-density mononuclear cell fraction was washed and seeded in expansion medium [33] consisting of high-glucose Dulbecco's modified Eagle's medium (DMEM), 12.5% fetal calf serum (FCS), 2 mM
Written consent was obtained from all tissue donors and the study received approval from the local ethics committee.
Chondrogenic induction of MSCs
Pellets consisting of 5×105 cells were exposed for 6 weeks to chondrogenic induction medium (high-glucose DMEM with 0.1 μM dexamethasone, 0.17 mM ascorbic acid-2 phosphate, 5 μg/mL insulin, 5 μg/mL transferrin, 5 ng/mL selenous acid, 1 mM sodium pyruvate, 0.35 mM proline, 1.25 mg/mL bovine serum albumin, and 1% penicillin/streptomycin) supplemented with 10 ng/mL recombinant human TGF-β1 (Pepro-Tech). Medium was changed three times a week. Where indicated, 2.5 nM of PTHrP(1–34), 2.5 nM or 1 μM of [Asn10, Leu11, D-Trp12] PTHrP(7–34) NH2 [PTHrP(7–34); all from Bachem], 10 μM of forskolin (Calbiochem/Merck Millipore), or 1 mM of N6,2′-O-dibutyryladenosine-3′:5′-cyclic monophosphate sodium salt (dbcAMP; Sigma-Aldrich Chemie) was supplemented.
For intermittent application, PTHrP(1–34), PTHrP(7–34), or forskolin was added to the culture medium to yield a final concentration of 2.5 nM or 10 μM, respectively, 6 h prior to every medium change starting on day 7 of the pellet culture. Culture supernatants were then replaced with fresh medium containing no PTHrP or forskolin. Since forskolin was dissolved in dimethyl sulfoxide (DMSO), respective DMSO controls were prepared for the pulsed application of forskolin. DMSO treatment alone yielded no significant effect in all experiments allowing to omit these controls in subsequent experiments.
RNA isolation and quantitative real-time PCR
Total RNA was isolated from two to six pooled, homogenized pellets per guanidinium isothiocyanate/phenol extraction (peqGOLD Trifast; Peqlab). Polyadenylated mRNA was purified from total RNA with oligo(dT)-coupled magnetic beads (Dynabeads; Invitrogen/Life Technologies). The mRNA was then subjected to first-strand cDNA synthesis using reverse transcriptase (Omniscript1; Qiagen) and oligo(dT) primers. The expression level of individual genes was analyzed by RT-PCR using the MX3000P QPCR System (Stratagene/Agilent Technologies). The following forward (F) and reverse (R) primer pairs were used for amplification: β-actin-F, 5′-CTCTTCCAGCCTTCCTT CCT-3′; β-actin-R, 5′-GATCCACACGGAGTACTTG-3′; Col2A1-F, 5′-TGGCCTGAGACAGCATGAC-3′; Col2A1-R, 5′-AGTGTTGGGAGCCAGATTGT-3′; Col10A1-F, 5′-CCC TTTTTGCTGCTAGTATCC-3′; Col10A1-R, 5′-CTGTTGT CCAGGTTTTCCTGGCAC-3′; IHH-F, 5′-CGACCGCAAT AAGTATGGAC-3′; IHH-R, 5′-GGTGAGCGGGTGTGA GTG-3′; PTH1R-F, 5′-GGTGAGGTGGTGGCTGT-3′; PTH1R-R, 5′-AGCATGAAGGACAGGAAC-3′; and MMP13-F, 5′-CTGGAGATATGATGATACTAAC-3′; MMP13-R, 5′-CACGCATAGTCATATAGATACT-3′. Gene expression levels were calculated as percentage of β-actin expression using the ΔCt method.
Measurement of ALP activity
Supernatants conditioned for 2 days were incubated with substrate solution (10 mg/mL of p-nitrophenyl phosphate in 0.1 M glycine, 1 mM MgCl2, and 1 mM ZnCl2, pH 9.6). After 150 min, the ALP activity was measured spectrophotometrically at 405/490 nm and was referred to a standard curve made from p-nitrophenol (Sigma-Aldrich Chemie).
Collagen extraction, quantification of collagen type II, and western blotting
Collagens were extracted as described elsewhere [5]. Briefly, pellets were homogenized, digested in pepsin buffer (0.5 M acetic acid, 0.2 M NaCl, and 2.5 mg/mL of pepsin), and precipitated with 4.5 M NaCl. Extracted collagens were resuspended in 400 μL of precipitation buffer (0.4 M NaCl and 0.1 M Tris Base, pH 7.4) and 1.2 mL of 100% ethanol and incubated for 4 h at −20°C. Precipitated collagens were resolved in lysis buffer (1% Triton X-100, 150 mM NaCl, and 50 mM Tris, pH 8.0). The collagen type II content of each lysate was determined by a native type II collagen detection ELISA (Chondrex).
For western blotting, amounts of lysate corresponding to 30 ng collagen type II were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and blotted onto a nitrocellulose membrane (Amersham Biosciences/GE Healthcare Europe). The membrane was cut horizontally and the lower part was probed with mouse anti-type X collagen antibody (1:500; Quartett) whereas the upper part was probed with mouse anti-type II collagen antibody (1:1,000; MP Biomedicals). Peroxidase-coupled goat anti-mouse antibody (1:5,000; Jackson ImmunoResearch Laboratories) was utilized as secondary antibody. Bands were visualized by enhanced chemiluminescence (Roche Diagnostics) and signals were quantified by densitometric integrity measurements using Bio1D v. 15.01.
Measurement of cAMP activity
Pellets at day 25 of chondrogenic differentiation were treated with indicated concentrations of forskolin, PTHrP(1–34), and PTHrP(7–34) in chondrogenic medium for 1 h. Culture supernatants were collected and the cAMP activity was determined by ELISA (R&D Systems).
Quantification of proteoglycan content
Pellets were lysed in guanidine-hydrochloride (GuHCl) buffer (50 mM Tris and 4 M GuHCl, pH 8.5) and unsoluble components were removed by centrifugation. One hundred microliters of each supernatant was mixed with 50 μL of SAT buffer (0.3% H2SO4 and 0.75% Triton X-100) and then supplemented with 750 μL of Alcian blue solution (0.3% Alcian blue, 0.16% H2SO4, 0.04 M GuHCl, and 0.35% Triton X-100). Precipitates were pelleted by centrifugation, resuspended in DMSO buffer (40% DMSO and 0.05 M MgCl2), and centrifuged at 16,000 g for 15 min at room temperature. The pellets were then solubilized in Gu-Prop buffer (4 M GuHCl, 33% 2-propanol, and 0.25% Triton X-100) and the proteoglycan content was determined by measurement of the absorbance at 570 nm.
After initial results from four donors, it became desirable to additionally determine the DNA content from the same sample. For this purpose we switched to an alternative proteoglycan quantification protocol (10/14 donors), in the course of which the pellets were digested with 100 μg/mL proteinase K (Roche Diagnostics), dissolved in Tris-HCl buffer (0.05 M Tris and 1 mM CaCl2, pH 8) overnight at 60°C. After digestion, 30 μL of each sample was mixed with 200 μL of a 1,9-dimethyl-methylene blue (DMMB) dye solution (40 mM glycine, 40 mM NaCl, and 48 μM DMMB) and absorbance was measured immediately at 530 nm.
Quantification of DNA content
The DNA content of pellets was determined using the Quanti-iT Pico-Green dsDNA kit (Invitrogen). For this assay, 20 μL of the Proteinase K-digested pellet samples prepared for the proteoglycan quantification was mixed with 80 μL TE buffer (200 mM Tris HCl and 20 mM EDTA) and PicoGreen solution. DNA content was determined by fluorescence measurement at 485/535 nm.
Histology
Pellets were fixed in 4% paraformaldehyde as described previously [34]. Sections (5 μm) were stained with Safranin O (Fluka) and counterstained with Fast Green (Chroma) to analyze the proteoglycan content. Immunohistologic staining of collagen type II was performed as described elsewhere [35]. Briefly, sections were incubated with a monoclonal mouse anti-human type II collagen antibody (1:1,000; MP Biomedicals) and reactivity was detected using biotinylated goat anti-mouse secondary antibody (1:500; Dianova), streptavidin-ALP (Dako), and fast red (Sigma-Aldrich Chemie).
Statistics
The mean and SD values were calculated for all variables. Differences between two groups were analyzed with the nonparametric Mann–Whitney U test or with Wilcoxon test for paired samples. Correlations were analyzed by determination of the Pearson's coefficient (r 2). P≤0.05 was considered significant. Data analysis was performed using SPSS 20.0.
Results
Intermittent PTHrP supplementation stimulates in vitro chondrogenesis
In confirmation of previous results [6], constant PTHrP treatment of MSCs throughout in vitro chondrogenesis resulted in cell pellets with virtually no proteoglycan and collagen type II deposition at day 42 (Fig. 1). When MSCs received intermittent PTHrP(1–34) treatment for 6 h three times a week, however, a collagen type II- and proteoglycan-rich matrix was deposited like in the non-PTHrP-treated control group according to histology (Fig. 1A, C). Quantitative analysis of the proteoglycan and collagen type II content of pellets (Fig. 2) revealed a significantly higher proteoglycan (mean 1.54-fold, P<0.01) and collagen type II (mean 2.94-fold, P<0.05) deposition in the PTHrP-pulsed group compared with controls. DNA levels were slightly, but significantly, elevated (P<0.05; Fig. 2C), suggesting enhanced survival or proliferation of cells during chondrogenesis under intermittent PTHrP treatment. Since the absolute proteoglycan/DNA content (n=10) was significantly higher (1.2-fold, P<0.01) after pulsed PTHrP treatment, not only higher cell numbers but also more proteoglycan deposition per cell contributed to superior matrix deposition (data not shown). Thus, as opposed to constant application, intermittent PTHrP treatment three times a week stimulated in vitro chondrogenesis of MSCs compared with the standard protocol.
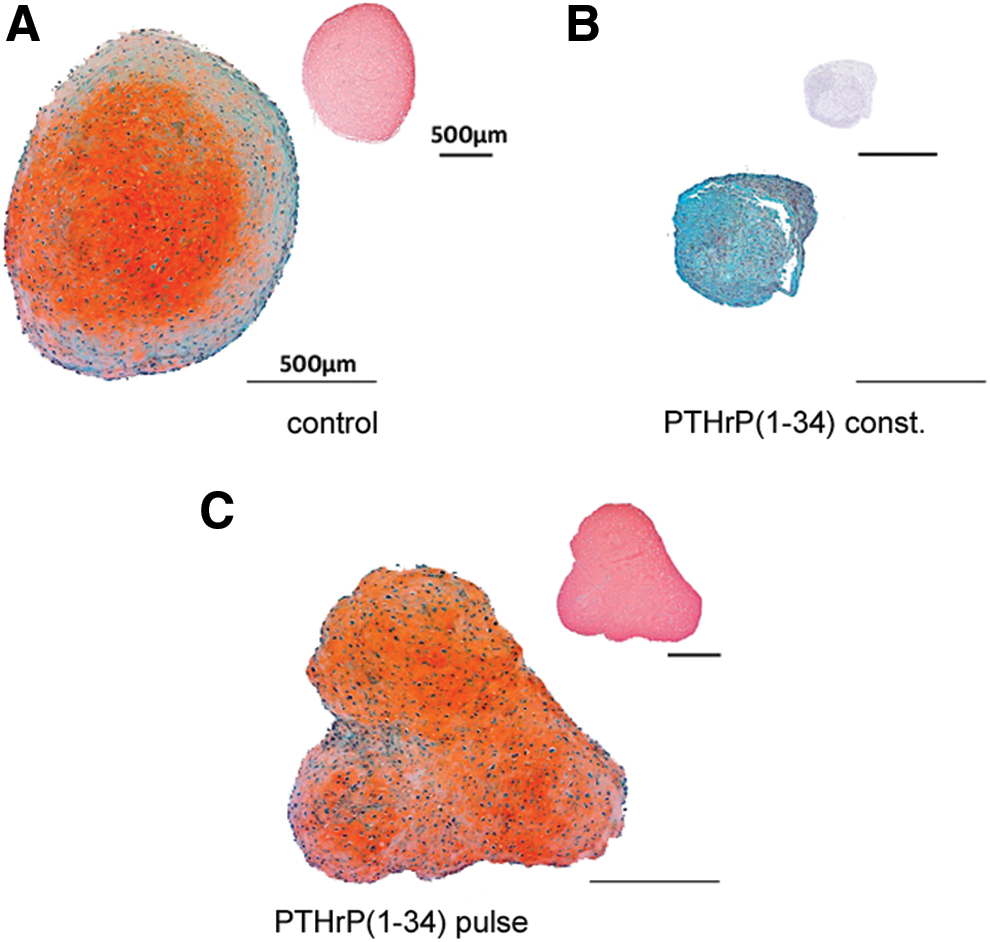
Histology of cartilaginous matrix deposition. Mesenchymal stromal cell (MSC) pellets were treated with chondrogenic medium alone
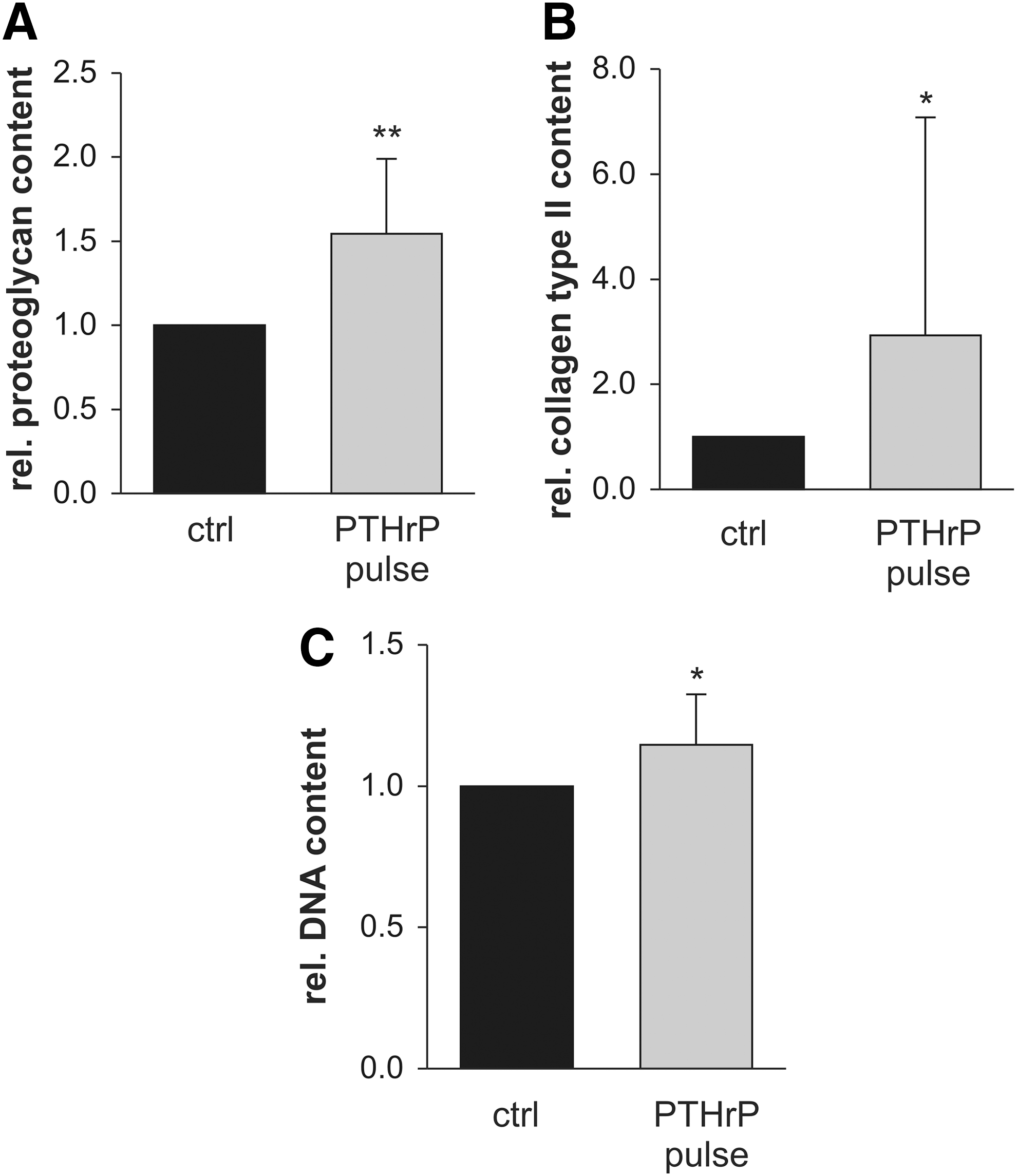
Biochemical analysis of MSC pellets after chondrogenesis. Pellets received chondrogenic medium alone (ctrl) or were additionally stimulated for 6 h with 2.5 nM PTHrP(1–34) before each medium change [parathyroid hormone-related protein (PTHrP) pulse; three times a week]. On day 42, pellets were harvested and the proteoglycan
Intermittent PTHrP supplementation reduces hypertrophy
To investigate whether the anabolic effect of intermittently supplemented PTHrP affected expression of chondrogenic and hypertrophic differentiation markers, gene expression of collagen type 2 (COL2A1), collagen type 10 (COL10A1), IHH, PTHR1 , and matrix metalloproteinase (MMP13) was quantified by quantitative real-time PCR. In line with collagen type II protein deposition (Fig. 2B), COL2A1 mRNA levels at day 42 were significantly higher in the PTHrP-pulsed group (2.1-fold; P<0.05, Fig. 3A). The COL10A1 mRNA levels were, however, rather similar (Fig. 3A) and no significant difference was obtained for the COL10A1/COL2A1 ratio (Fig. 3B). Time course analysis (Fig. 3C) demonstrated that COL2A1 expression was stimulated especially in the late phase of MSC chondrogenesis.
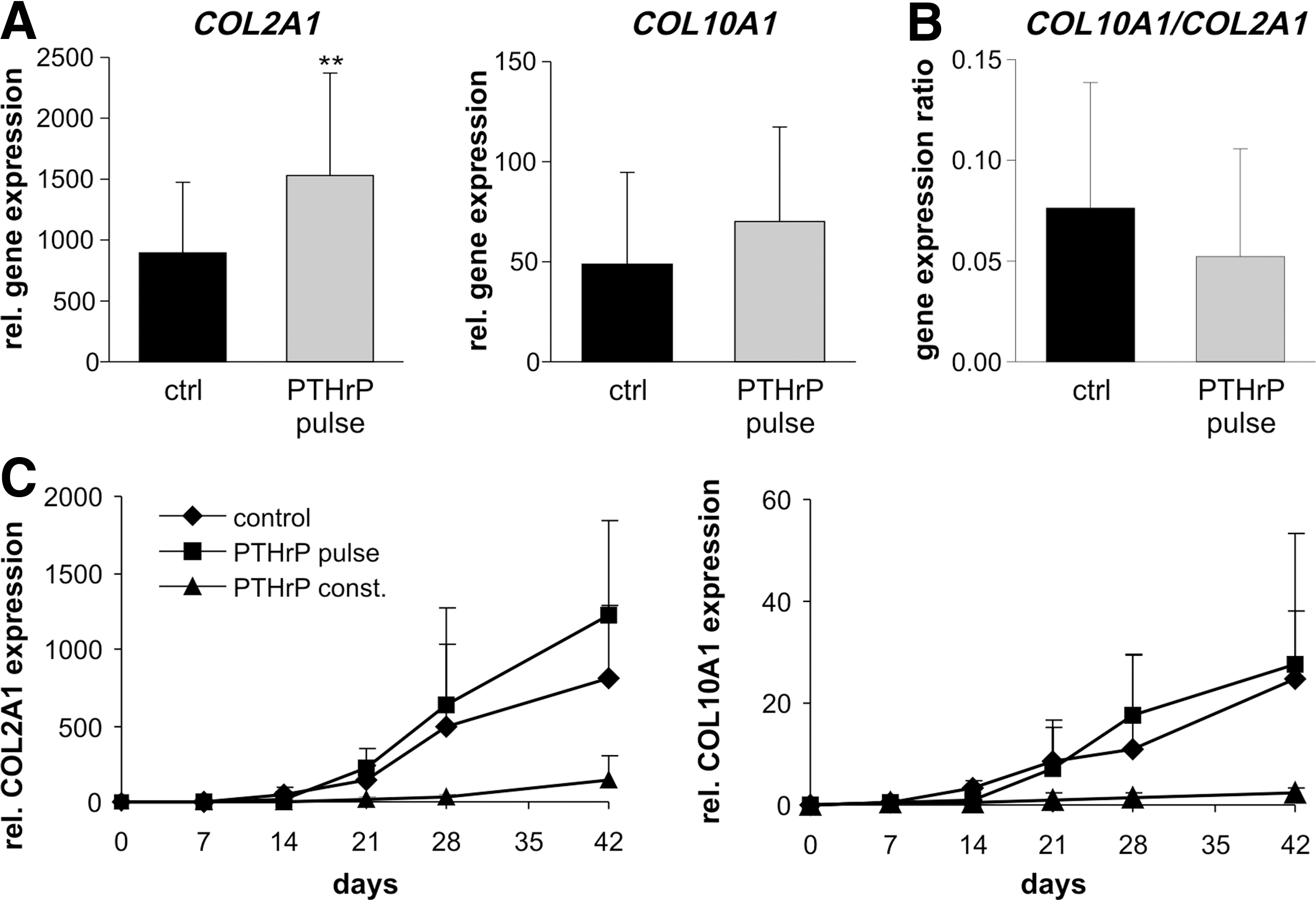
Gene expression analysis of collagen type 2 (COL2A1) and collagen type 10 (COL10A1). MSC pellets were treated with chondrogenic medium alone (ctrl) or chondrogenic medium additionally supplemented with 2.5 nM PTHrP(1–34) either constantly (PTHrP const.) or intermittently (PTHrP pulse). RNA was extracted from MSC pellets at the indicated time points and mRNA levels of COL2A1 and COL10A1 were determined using quantitative real-time PCR and standardized to β-actin levels.
Samples with better chondrogenesis show higher collagen type II content and usually contain also more collagen type X. To normalize collagen type X deposition to the degree of chondrogenesis of pellets, collagen type II was used as reference protein in western blot analysis. Densitometric analysis of protein bands from the same pellet reflected successful adjustment to collagen type II content and revealed no coherent tendency with respect to an increase or decrease of collagen type X deposition at termination of culture (Fig. 4A, B). As a result, neither the COL10A1/COL2A1 mRNA ratio (Fig. 3B) nor the collagen type X/collagen type II protein ratio (Fig. 4C) was different between the PHTrP pulse and the control groups, indicating no specific effect of pulsed PTHrP on the hypertrophic marker collagen type X.
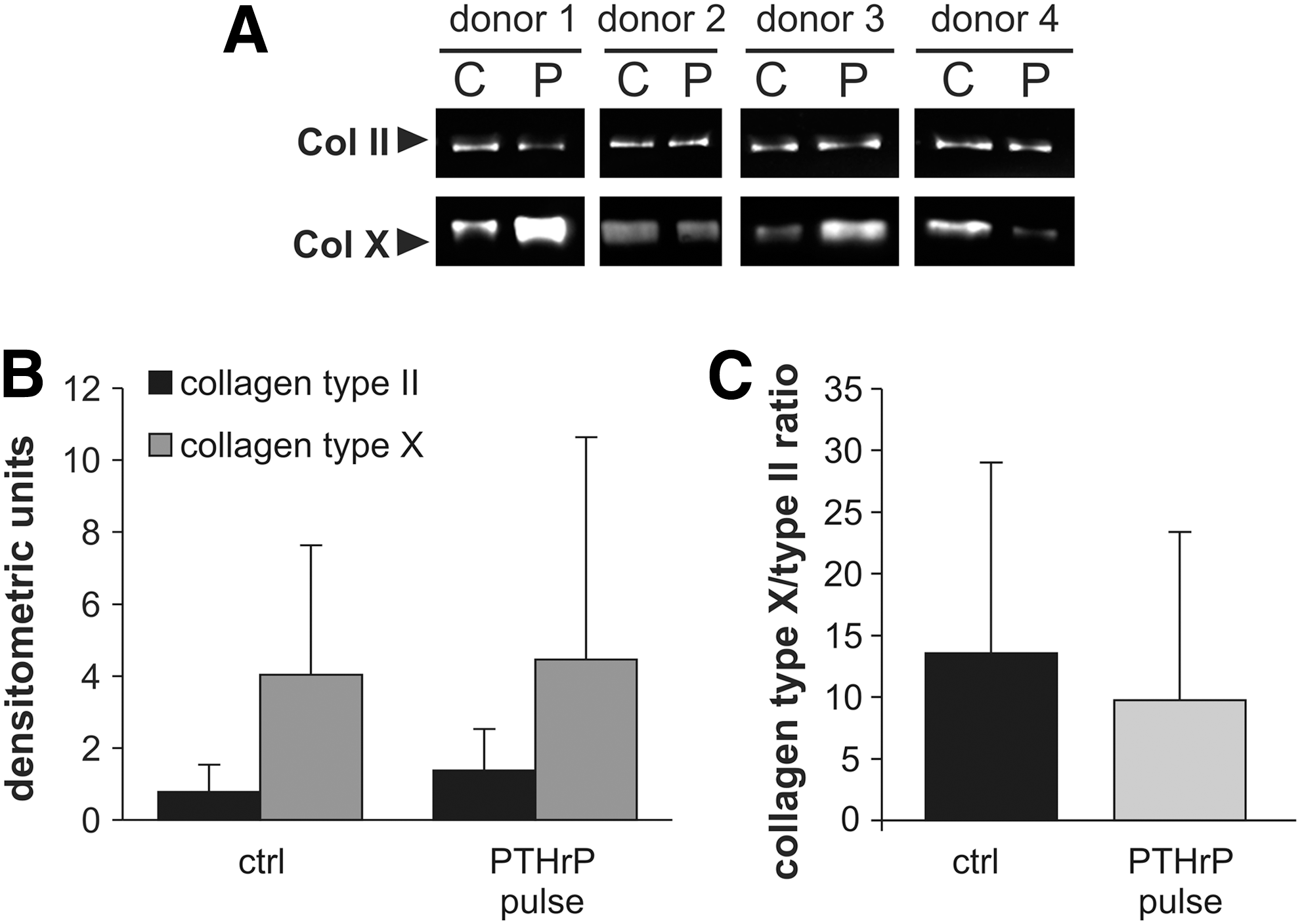
Western blot analysis of the collagen type X/collagen type II expression ratio of pellets from the untreated positive control group receiving transforming growth factor-beta (TGF-β) but no PTHrP (C) compared with pellets that additionally received intermittent PTHrP(1–34) (P) during chondrogenesis.
Pulsed application of PTHrP did not alter PTH1R mRNA expression levels versus the positive control, indicating no differential ligand sensitivity in response to intermittent PTHrP treatment. However, samples with higher PTH1R expression levels showed a higher stimulation of COL2A1 expression compared with samples with lower PTH1R expression levels. Pearson's coefficient of PTH1R and ΔCOL2A1 (COL2A1 expression pulse group−COL2A1 expression control group) demonstrated a significant correlation (r 2=0.902, P<0.01, n=16 samples from four experiments), suggesting that a correlation between ligand sensitivity and the extent of COL2A1 stimulation may exist (Fig. 5B). At day 42, IHH mRNA levels were, however, significantly reduced under pulsed PTHrP treatment (Fig. 5C) (decrease to 70%, P<0.05) in line with an inhibitory effect of PTHrP on IHH expression leading to reduced hypertrophy. The mRNA levels of the terminal differentiation marker MMP13 were not altered (Fig. 5D). Importantly, the enzyme activity of ALP in culture supernatants was significantly reduced in the pulsed PTHrP group from day 14 on, reaching 67% of control ALP levels at day 42 (P<0.01, Fig. 5E). Altogether this demonstrated that intermittent PTHrP supplementation of MSCs during chondrogenesis enhanced COL2A1 expression and concomitantly reduced hypertrophy according to IHH levels and ALP enzyme activity.
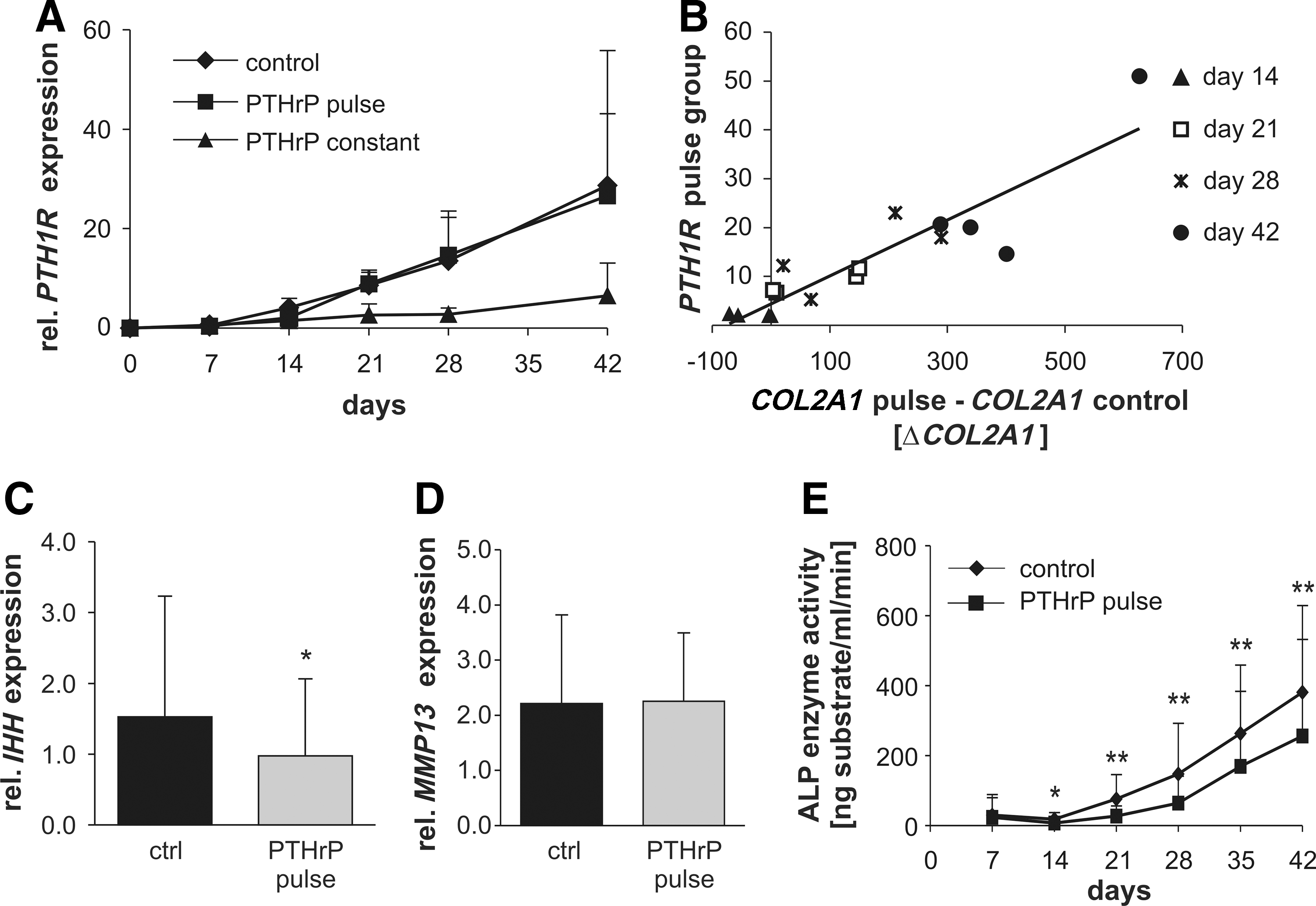
Expression of PTH/PTHrP receptor (PTH1R), Indian hedgehog (IHH), MMP13, and alkaline phosphatase (ALP) enzyme activity. Pellets received chondrogenic medium alone (ctrl) or were additionally stimulated with 2.5 nM PTHrP(1–34) either constantly (PTHrP const.) or intermittently (PTHrP pulse).
Constant PTHrP(1–34) inhibits chondrogenesis via cAMP/PKA signaling
Since constant and intermittent application of PTHrP exerted opposing effects on MSC chondrogenesis, we sought to clarify whether these might be mediated via divergent signaling pathways. Forskolin is an activator of adenylate cyclase, whereas the N-terminally truncated PTHrP(7–34) fails to elicit a cAMP response. Detection of cAMP levels confirmed a stimulation of cAMP levels by 10 μM forskolin in MSCs at day 25 of chondrogenesis and a lack of cAMP signaling by PTHrP(7–34) but not PTHrP(1–34) treatment (Fig. 6A). In line, constant application of 2.5 nM PTHrP(1–34) or 10 μM forskolin blocked chondrogenic differentiation, according to strongly decreased proteoglycan and collagen type II staining in histology, while 2.5 nM PTHrP(7–34) displayed no effect (Fig. 6B). To investigate how long it takes until constant PTHrP exposure displays its negative effect on COL2A1 and COL10A1 expression and on ALP enzyme activity and to decide whether small-molecule-induced activation of the cAMP/PKA pathway parallels timing of PTHrP(1–34) effects, chondrogenic pellets were treated for 2 or 4 days with PTHrP(1–34), forskolin, and the cAMP-analogon dbcAMP, respectively. Stimulation of the cAMP/PKA pathway with forskolin and dbcAMP from day 21 on reached a significant decline of COL2A1 and COL10A1 mRNA levels at 96 h but not 48 h (Fig. 6C) and paralleled the effects of PTHrP(1–34). This indicated that not hours but rather several days of PTHrP exposure are needed before negative effects on COL2A1 expression become significant. Inhibition of ALP activity, however, was already significant after 48 h in all three treatment groups. This demonstrated that constant presence of PTHrP(1–34) during chondrogenesis influenced MSCs predominantly via activation of the cAMP/PKA pathway and may affect ALP enzyme activity earlier than COL2A1 expression.
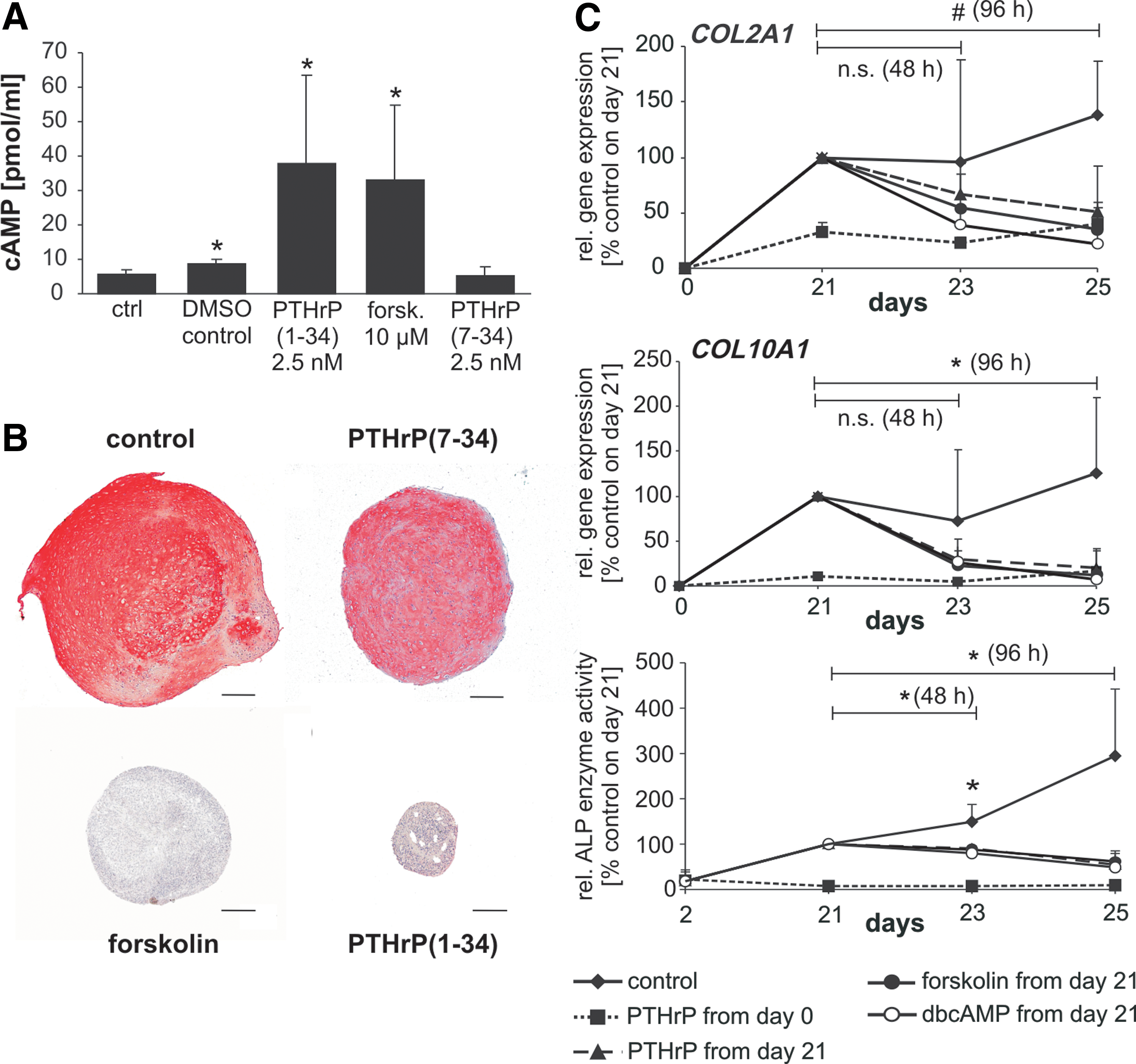
Effects of agonists of PTHrP signaling on MSC chondrogenesis.
Intermittent PTHrP(1–34) affects proteoglycan and DNA via cAMP/PKA signaling
To test whether the effects of the intermittent PTHrP treatment may be explained by activation of the cAMP/PKA or the PKC pathway, MSCs were intermittently treated with either 2.5 nM PTHrP(1–34), 2.5 nM PTHrP(7–34), or 10 μM forskolin. Intermittent forskolin, but not PTHrP(7–34), mimicked the positive effects of pulsed PTHrP(1–34) on proteoglycan (Fig. 7A) and DNA content of day-42 pellets (Fig. 7B). The significant suppression of ALP activity by pulsed PTHrP(1–34) to 61% at day 42 (P<0.05, Fig. 7C) was, however, not fully imitated by intermittent forskolin treatment, which induced only a trend to lower ALP values (74%, P=0.063). Pulsed PTHrP(7–34) could neither enhance proteoglycan and DNA content nor influence ALP activity (Fig. 7). In conclusion, positive effects of intermittent PTHrP(1–34) on MSC chondrogenesis were imitated by cAMP/PKA (forskolin) but not PKC [PTHrP(7–34)] signaling while ALP downregulation was not fully reproduced by either pathway.
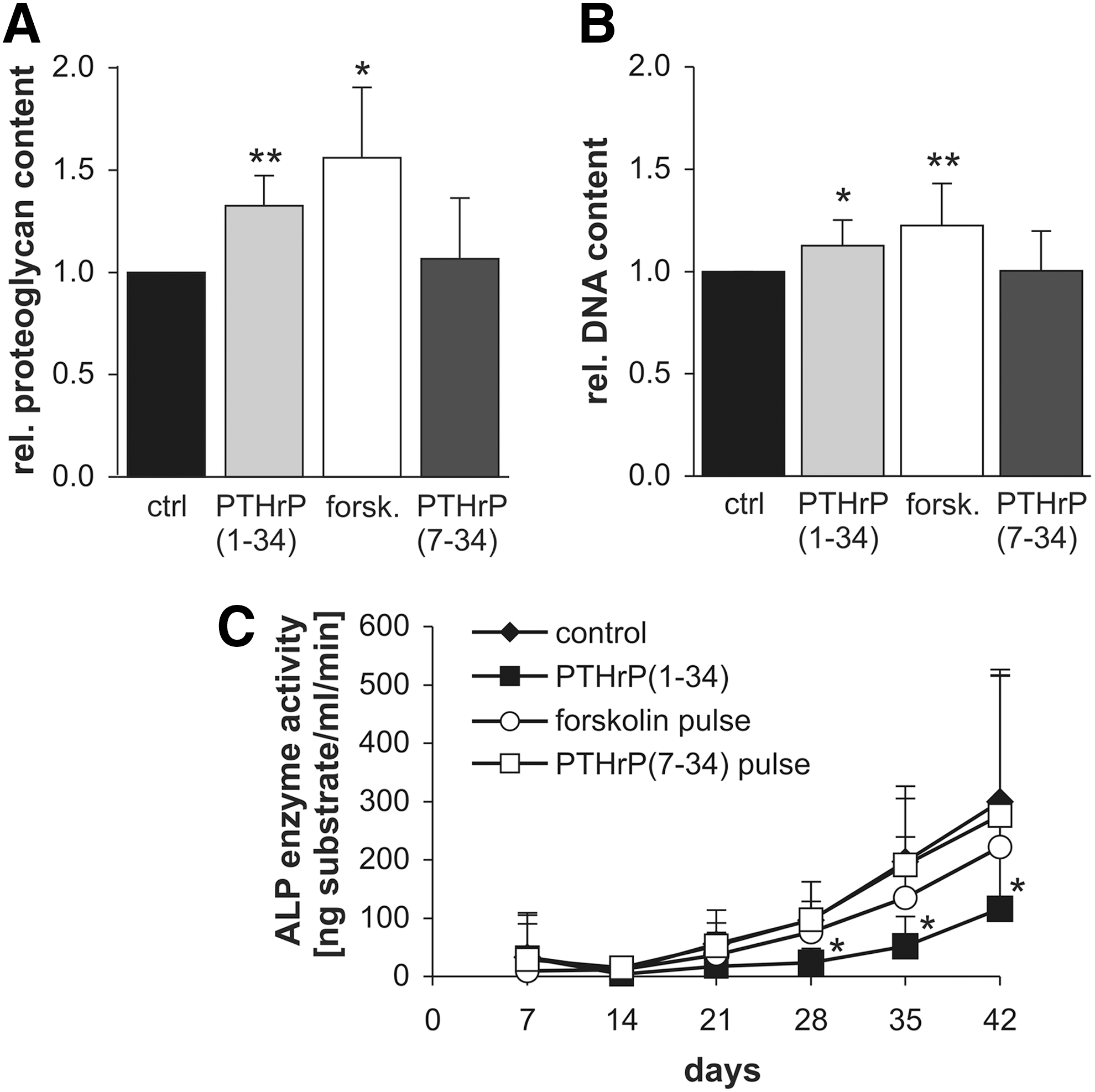
Biochemical analysis after intermittent forskolin or PTHrP(7–34) treatment. Pellets received chondrogenic medium alone (ctrl) or were additionally stimulated for 6 h with either 2.5 nM PTHrP(1–34), 2.5 nM PTHrP(7–34), or 10 μM forskolin (forsk) before each medium change, respectively.
Discussion
Under common in vitro differentiation protocols, chondrogenesis of MSCs leads to undesired phenotype instability and premature hypertrophy of the developing chondrocytes. Due to its documented antihypertrophic properties, PTHrP is an interesting candidate molecule to improve chondrogenesis of MSCs; however, constant treatment of MSCs with PTHrP during chondrogenesis, unfortunately, resulted in a suppression of differentiation rather than a selective inhibition of hypertrophic features of chondrocyte development [6]. In view of a well-documented switch from a catabolic to an anabolic response in bone when PTH/PTHrP is administered intermittently instead of constantly [22], we here exposed MSCs to a pulsed instead of constant PTHrP regimen during chondrogenesis, in order to evoke beneficial effects on MSC chondrogenesis. This indeed allowed us to guide MSCs to chondrogenic differentiation under significant elevation of the DNA, proteoglycan, and collagen type II content of pellets and reduced signs of hypertrophy.
A significantly increased proteoglycan/DNA ratio argued in favor of the envisaged anabolic stimulation of the cells. Further, higher cell numbers per pellet suggested either a reduced loss of cells by apoptosis, which is abundant in MSC pellets during early chondrogenesis [36], and indeed PTHrP action has been linked to the prevention of apoptosis before [37]. Alternatively, pulsed PTHrP may have increased proliferation during pellet culture [33] in line with a documented influence on cell proliferation via cyclin D1 [38] and the cyclin kinase inhibitor p57 [39]. In the context of bone formation, the anabolic action of PTH was attributed to enhanced osteoblast or preosteoblast survival or differentiation and not to cell proliferation [40,41], but to unravel the exact mechanism operational in our anabolic chondrogenic setting, highly sensitive quantitative assays applicable to 3D culture are required.
Higher COL2A1 mRNA levels versus controls were observed especially during the last 2 weeks of chondrogenesis when PTHrP receptor levels are high, indicating an anabolic effect mainly on the more mature cells. This rules out that PTHrP just delayed the progression of chondrocyte development due to its presence for about 10% of the total time interval. Thus, enhanced proteoglycan deposition and COL2A1 expression per cell is not just a result of keeping the cells longer in a matrix-producing stage, but it is a true anabolic action based on stimulation of gene expression via PTHrP. Several reports suggested that the anabolic action of PTH involves, at least in part, the action of insulin-like growth factor 1 (IGF-1) [19] and studies are ongoing to evaluate whether regulation of IGF-1 may also be important for stimulation of chondrogenesis.
Interestingly, we see no uniform tendency for COL10A1 mRNA levels and collagen type X deposition in pellets. Yoshida et al. monitored the time course of IHH, patched 1 (PTCH1), osteopontin (OPN), and COL10A1 mRNA inhibition by PTH(1–84) in chondrocytes in vitro [42] and found that IHH, PTCH1, and OPN mRNA expression was reduced rapidly within 4–6 h after PTH treatment, whereas COL10A1 was downregulated only after 24 h. Others similarly reported a peak influence of PTHrP on collagen type X expression in growth plate chondrocytes after 24 h [43]. Pulsed PTHrP treatment for 6 h was obviously sufficient to effectively stimulate COL2A1 expression but too short to exert a profound effect on COL10A1 expression. After longer PTHrP exposure at 48 h (day 23), mean mRNA levels for COL2A1, COL10A1 (Fig. 6C), and IHH (not shown) were reduced and a significant downregulation was reached after 96 h. We here selected a PTHrP pulse duration of 6 h in accordance with a previous study, where 6 h—but not 1, 3, or 24 h—of pulsed PTH treatment exerted anabolic effects on osteoblastic monolayer cultures [19]. Further, 6 h but not constant treatment of rat MSCs with PTH in monolayer cultures reduced ALP activity [21]. In view of the clearly time-dependent effects of PTHrP on MSCs during chondrogenesis, it is next important to clarify whether a time window of PTHrP exposure may exist, in which not only ALP and IHH but also COL10A1 can be downregulated while COL2A1 is still stimulated. However, as MMP13, PTH1R, and COL10A1 expression was not significantly affected by intermittent treatment with PTHrP compared with the positive control, this clearly indicates that hypertrophic maturation is further linked to other signaling pathways apart from PTHrP signaling, such as the transforming growth factor-beta (TGF-β) superfamily pathways [44].
Interestingly, ALP activity in pellets is often more intense around cells located in the periphery of the pellet in areas of less proteoglycan deposition possibly due to higher oxygen and nutrient exposure favoring endochondral differentiation [9]. These cells often do not look like chondrocytes but have a more fibroblastic or osteoblast-like appearance and may contribute a large part of the ALP secreted into culture supernatants. The distance of PTHrP diffusion is an important parameter in growth plate development and the rate of diffusion of PTHrP may strongly depend on the presence and composition of the cartilaginous matrix. Thus, it is tempting to speculate that in our model peripheral cells experience higher concentration of PTHrP or longer PTHrP exposure than chondrocytes in the pellet center. If the antihypertrophic action of PTHrP may guide more cells in the periphery toward a chondrogenic instead of endochondral development, then this could contribute to more matrix deposition per pellet and reduced ALP activity in supernatants. According to Figure 1C, some pellets exposed to pulsed PTHrP indeed seemed to be devoid of a noncartilaginous peripheral cell layer; however, this was not a consistent observation throughout. A spatially restricted or stronger inhibition of the peripheral cells by pulsed PTHrP could also allow more nutrients and oxygen to pass into the pellet center, stimulating improved cartilage matrix deposition by an indirect effect even not requiring PTHrP signaling in the chondrocytes themselves. However, the importance of PTHR1 signaling duration is also seen in osteoblast monolayers, where 1 h and continuous PTH stimulation decreased ALP activity of rat osteoblasts, while 3- and 24-h stimulation had no effect and 6 h stimulated ALP activity [19]. Further, we reproduced most of the effects of pulsed PTHrP by the small-molecule inhibitor forskolin having different diffusion kinetics, suggesting that 3D effects are only of minor importance for an explanation of anabolic versus catabolic PTHrP effects in MSC chondrogenesis.
Overall, we encountered high donor variabilities leading to high standard deviations and the need to provide a high sample number to achieve statistical significancy throughout all experiments. A high donor variability regarding chondrogenic progression of MSCs in pellet culture and in parameters of chondrogenic matrix deposition is well established in the field. This striking interdonor variability and even intradonor variation between individual pellet samples was a main topic of one of our recent studies [45]. We demonstrated that the proliferation capacity of MSCs at initiation of chondrogenic culture strongly correlated with speed of chondrogenic progression during culture and with outcome. Obviously high anabolic conditions favor reduced apoptosis and faster start of chondrogenesis since less cells may die and more cells can enter matrix deposition under such conditions. Further, cells underwent a transient proliferation stage essential for a successful chondrogenesis starting earlier in “fitter” cultures compared with others [33]. We here examined all pellets at fixed time points after switch to induction conditions, not taking into account the individual timing of the sample population. Therefore the cells may be in different maturation stages at the time of investigation, yielding different expression levels of chondrogenic and hypertrophic markers. In line with this, the standard deviation usually increases with time in culture and typically is highest at the late time points.
PTHrP signals via numerous divergent pathways [46,47], but most of its impact on chondrocyte hypertrophy [48,49] as well as the PTH/PTHrP-mediated anabolic effects on bone mass or osteoblast differentiation [50,51] has been linked to the cAMP/PKA-dependent pathway. Little was so far known about the molecular mechanisms of PTHrP action on MSCs during in vitro chondrogenesis. In line with the previous results we show here for the first time that stimulation of cAMP/PKA signaling by forskolin was able to fully reproduce both the negative effects of constant PTHrP application and the positive anabolic effects of intermittent PTHrP exposure on chondrogenic extracellular matrix deposition. PTHrP(7–34) capable of PKC but not of PKA activation [15,16,52] instead was ineffective under both application regimes. This demonstrated that PTHrP exerted its catabolic and anabolic influence on MSC chondrogenesis mainly via the cAMP/PKA pathway under both modes of administration, underlining that signal duration is the decisive variable. Differential duration of PKA signals may feed back on PTH1R expression levels, ligand sensitivity of cells, and crosstalk with other downstream signaling pathways, leading to receptor desensitization or altered availability of costimulatory molecules and repressors to regulate gene transcription. In line with the first two explanations, PTH1R is not upregulated during chondrogenesis of MSCs under constant PTHrP exposure [6] and samples with high PTH1R levels responded with a higher stimulation of COL2A1 expression compared with samples with lower levels as evident from the strong correlation of receptor levels with this anabolic response (Fig. 5B).
Besides its ability to mimick PTHrP effects on MSC anabolism, forskolin only showed a nonsignificant tendency to reduce ALP activity. It is possible that higher concentrations of forskolin would have induced more pronounced effects, but cAMP generation of forskolin at the chosen concentration of 10 μM nearly equaled that of 2.5 nM PTHrP(1–34) and therefore represented a good approximation to the used PTHrP concentration. Ishizuya et al. proposed a cooperative contribution of both PKA and PKC activation on the anabolic stimulation of osteoblasts, where only a simultaneous activation of both pathways could mimic anabolic intermittent PTH effects [19]. We therefore also tested a combined administration of pulsed PTHrP(7–34) and forskolin, which had, however, no additional effect to forskolin in our model (data not shown). Thus, the cAMP/PKA pathway seems to be dominant for the prochondrogenic and antihypertrophic effects of intermittent PTHrP in MSC chondrogenesis, whereas involvement of the PKC-dependent pathway seems less important. Additional studies on the noncanonical pathways of PTH1R signaling are, however, needed to fully clarify this issue.
In conclusion, intermittent treatment of MSCs during chondrogenesis represents a novel and simple means to augment cartilaginous matrix deposition together with a simultaneous suppression of undesired hypertrophic features. In spite of opposing effects of constant versus intermittent PTHrP application on chondrogenic differentiation markers, both treatment modes influenced the MSCs mainly via the cAMP/PKA-dependent pathway, suggesting that PKC signaling is less important. Overall our data are encouraging to further optimize signal duration and pulse frequency, in relation to agonist concentration, to fully deploy the potential of pulsed PTHrP application on MSC chondrogenesis, and in order to try to group COL10A1 regulation together with the other hypertrophic markers of reduced expression. In this context, ectopic transplantation of treated pellets in mice will be important to verify that the beneficial results of pulsed PTHrP treatment are not just a transient effect.
In view of this prochondrogenic and antihypertrophic effect seen in vitro and first promising in vivo results [28], it is now attractive to consider PTHrP injections, as used for osteoporosis prevention, as a means to support MSC-based cartilage regeneration strategies in a clinical setting.
Footnotes
Acknowledgments
This work was supported by funds of the German Research Foundation (DFG Ri707/7-1) and the Orthopaedic University Hospital Heidelberg. The authors thank Simone Gantz for statistical advice, Birgit Frey for assistance in histology, and Melanie Weisser and Rosalie Bock for technical support.
Author Disclosure Statement
No competing financial interests exist.