
Abstract
There is surprising confusion surrounding the concept of biological totipotency, both within the scientific community and in society at large. Increasingly, ethical objections to scientific research have both practical and political implications. Ethical controversy surrounding an area of research can have a chilling effect on investors and industry, which in turn slows the development of novel medical therapies. In this context, clarifying precisely what is meant by “totipotency” and how it is experimentally determined will both avoid unnecessary controversy and potentially reduce inappropriate barriers to research. Here, the concept of totipotency is discussed, and the confusions surrounding this term in the scientific and nonscientific literature are considered. A new term, “plenipotent,” is proposed to resolve this confusion. The requirement for specific, oocyte-derived cytoplasm as a component of totipotency is outlined. Finally, the implications of twinning for our understanding of totipotency are discussed.
Highlights
• Inaccurate use of the term “totipotent” by scientists creates unnecessary ethical controversy.
• Public concern over producing embryos by reprogramming reflects confusion over totipotency.
• Twinning by blastocyst splitting does not provide scientific evidence for totipotency.
What Is Totipotency?
T
The difference between these two definitions is not trivial. Producing a mature organism requires the ability to both generate all the cells of the body and to organize them in a specific temporal and spatial sequence, that is, to undergo a coordinated process of development. Totipotency in this strict sense is demonstrated by the ability of an isolated cell to produce a fertile, adult individual. Consequently, a cell that is totipotent is also a one-cell embryo; that is, a cell that is capable of generating a globally coordinated developmental sequence.
While stem cells, tumors, and embryos have many molecular features in common, embryos are clearly organisms [5
–8]. Embryos develop in a predictable manner toward a species-specific adult form (human embryos do not mature into mice, monkeys, or tumors). Embryos repair injury. They adapt to changing environmental conditions. Most importantly, they show coordinated interactions between parts (molecules, cells, tissues, structures, and organs) that promote the survival, health, and continued development of the organism as a whole; that is, interactions that are characteristic of “an individual constituted to carry on the activities of life by means of organs separate in function but mutually dependent: a living being,” (
The ability to produce an orchestrated developmental sequence should not be misconstrued as some kind of mystical element that is merely attributed to an embryo. The fact that the embryo undergoes a self-directed process of maturation is entirely a matter of empirical observation; embryos construct themselves. Scientists tend to view this developmental capability as a series of cellular/molecular events. Others may view human development in more spiritual or poetic terms. But neither of these views alters the scientific facts; embryos manifest a unique molecular composition and pattern of behavior that is characteristic of an organism (ie, “a living being”) that has not been observed in tumors or other human cells.
Regardless how individuals or societies ultimately weigh the value of the embryo relative to the value of scientific research, it is important to appreciate that in all cases, the ethical consideration given to human embryos does not reflect the status they will achieve at some point in the future (ie, what they will mature into). If this was the case, then there could be no possible objection to embryo-destructive research since, by definition, adult status is never attained in such situations. Rather, ethical consideration is given to human embryos based on the status they already possess; that is, their unique and fully operative ability to function as a human organism. Therefore, ethical controversy regarding totipotent human cells only concerns cells that are totipotent in the strict, organismal sense; that is, a cell that is a human embryo.
The term totipotent describes the properties of an individual cell (not a group of cells) with the two meanings of this term roughly corresponding to the progressive restriction in potential cells exhibit during normal development. At the expanded blastocyst stage (Fig. 1A), cells in specific regions of the embryo are restricted to produce only a subset of the cells observed at more-mature stages (Table 1). Restrictions in potency occur gradually as development proceeds, with the ability to independently initiate a developmental sequence (ie, totipotency in the first sense) being lost relatively early during development, and individual cells of the late morula and early blastocyst producing a wider range of derivatives than cells at the expanded blastocyst stage (Fig. 1B).
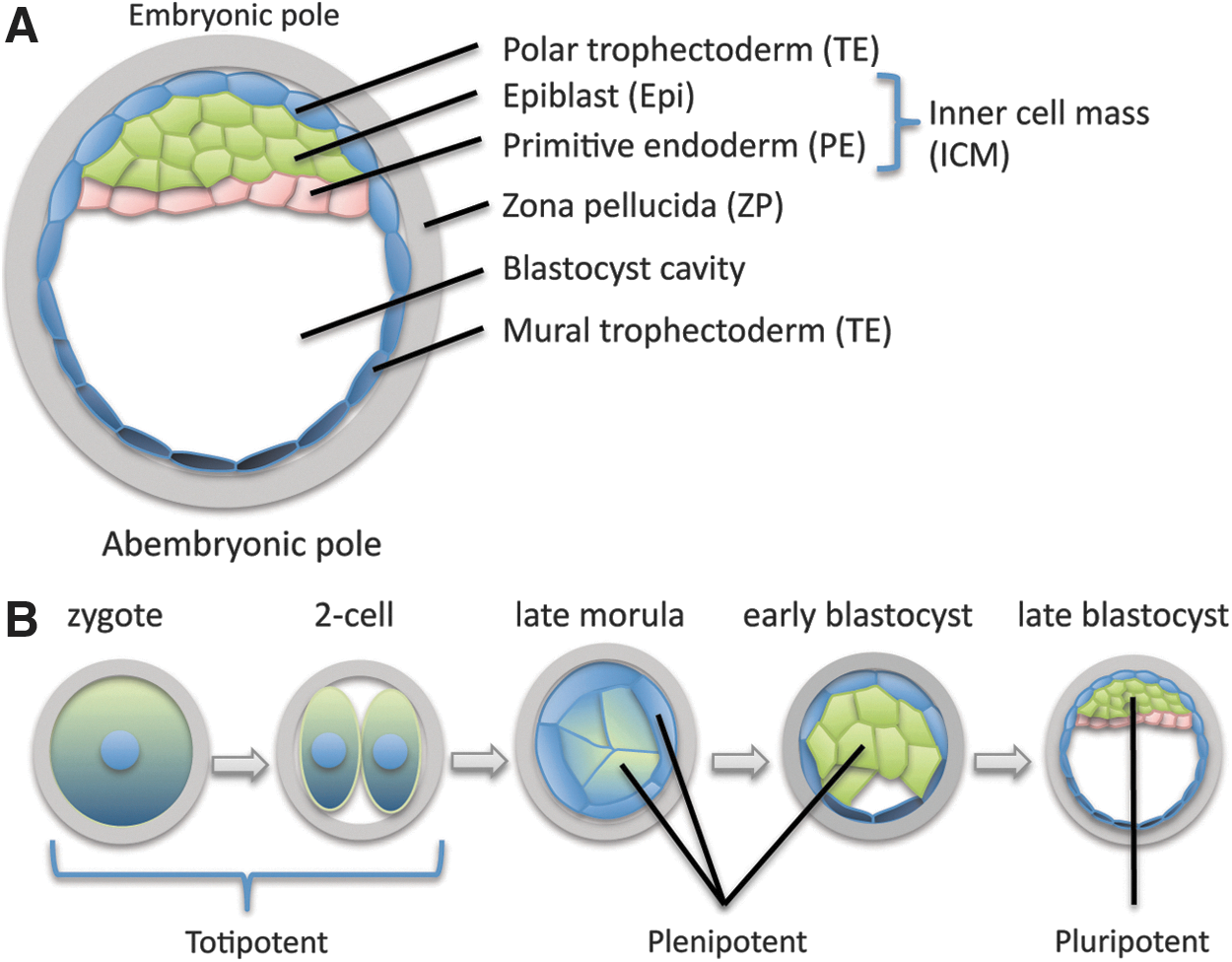
Cell types and potency at different developmental stages.
In humans, PE is also referred to as hypoblast.
Epi, epiblast; ICM, inner cell mass; PE, primitive endoderm; TE, trophectoderm.
The confusion between the two senses of totipotency has led some to propose new terminology, with one author suggesting the term “totipotent” be reserved for organisms, while stem cells and tumors that produce all cell types but do not organize them into a coherent body plan would be referred to as “omnipotent” [9]. Yet the strong connotation of this word outside the field of science compromises its utility. “Plenipotent” (from the Latin plenus, or “full”) would be preferable for individual cells that are totipotent in the cellular or weak sense, reserving “pluripotent” (from the Latin pluris, or “more”) for cells that make only a subset of the normal derivatives of the embryo (Fig. 1B and Table 2). These terms would better reflect the progressive restrictions in potency that occur during normal development (Fig. 1B), and avoid the unfortunate application of the term “totipotent” to cells that are not embryos.
Cell types that have met some or all of the experimental tests listed are given as examples, with cases in which there is some debate over the evidence being given in italics. The tests are listed in rough order of stringency, with the first test being the only definitive proof for the specified level of potency. Teratoma classification follows the designation of Cunningham et al. [193]. Note: These terms apply only to individual cells, not to groups of cells.
Testing the developmental potency of human cells is limited by ethical constraints. hESCs, iPSCs, and embryonal carcinomas are plenipotent based primarily on the last two tests and on limited data from chimeric mice.
Primordial germ cells are plenipotent based primarily on yolk-sac tumor/teratoma formation.
Some mESCs and iPSCs are able to form PE-derived cells under some culture conditions and can make limited contributes to both TE- and PE-derived structures in chimeric mice, suggesting that they may be plenipotent.
hESCs, human embryonic stem cells; iPSCs, induced pluripotent stem cells; mESCs, mouse embryonic stem cells.
In many cases, there are significant limitations in the data available to classify the developmental capabilities of cell types, particularly for human cells. Moreover, not all studies agree on the potency of the same cell type. Consequently, as is the case for all classifications, those suggested in Table 2 can only be taken as provisional, and must be interpreted relative to a specific body of work. Yet individual studies can be evaluated based on the evidence they present. For example, a recent study of embryonic stem cells (ESCs) [10] suggests neither human nor mouse cells produce trophectoderm (TE) derivatives, a finding that would classify both cell types as pluripotent. In contrast, other studies indicate that human ESCs (hESCs) are able to produce both trophoblast and syncytiotrophoblast, a finding that would classify hESCs as plenipotent [11,12]. Similarly, several recent studies indicate that subpopulations of mouse ESCs (mESCs) are able to differentiate as TE derivatives [4,13,14], which would classify this subset as plenipotent. In contrast, other studies of mESCs either indicate that TE derivatives can only be induced by artificial manipulation of specific genes [15] or, conversely, that the great majority of mESCs are able to produce TE derivatives under specific culture conditions [16,17], further illustrating that the potency of specific types of stem cells has not been conclusively resolved in the literature. Nonetheless, relating the degrees of potency observed in cell lines to the progressive restrictions in potency observed in development (Fig. 1B and Table 1) provides a more accurate classification and avoids the confusing misapplication of the term “totipotent.”
Currently, most scientists refer to ESCs from both mice and humans as “pluripotent,” without attempting to distinguish between the progressive restrictions in potency that are seen in embryos (Fig. 1B). Consequently, in the current analysis, the term totipotent (without quotes) will be used in the organismal or strict sense, and pluripotent will be used as it is currently used in the literature, that is, to refer to cells that produce, but do not organize, all or most of the cell types found in a mature individual (ie, to refer to cells that are both “plenipotent” and “pluripotent,” as defined in Table 2).
Confounding Usage of the Term “Totipotent” in the Scientific Literature
Although many scientific articles use “totipotent” in a consistent manner [18 –21], a surprising number use the term less rigorously, conflating the two meanings and/or accepting inadequate evidence as proof of strict totipotency. These misapplications fall into four general classes: (1) equating participation in development with the ability to independently generate a developmental sequence, (2) equating the ability of groups of cells to collectively generate a developmental sequence with totipotency of individual cells, (3) equating the expression of early embryonic markers with totipotency, and (4) taking a partial or superficial resemblance to an embryo as evidence for totipotency.
Participation in development is not totipotency
The most common misapplication of the term totipotent in both the scientific and nonscientific literatures is to equate the ability of a cell to participate in an embryonic process with the ability to generate all of the structures of the body independently (Fig. 2). For example, the observation that stem cells will generate all or most of the tissues of the postnatal body after injection into an early stage embryo [22,23] or in a tetraploid complementation assay [24] is often misinterpreted as evidence that stem cells are totipotent [25 –28]. Indeed, the generation of live mice from tetraploid complementation using reprogrammed pluripotent cells [29] has been cited as evidence that “somatic cells alone have the potential to become an adult human being” [30]. Some go so far as to conclude, “a reprogrammed human cell is not fundamentally different from a nuclear-transfer or natural fertilization zygote in its ability to become a fetus” [31].
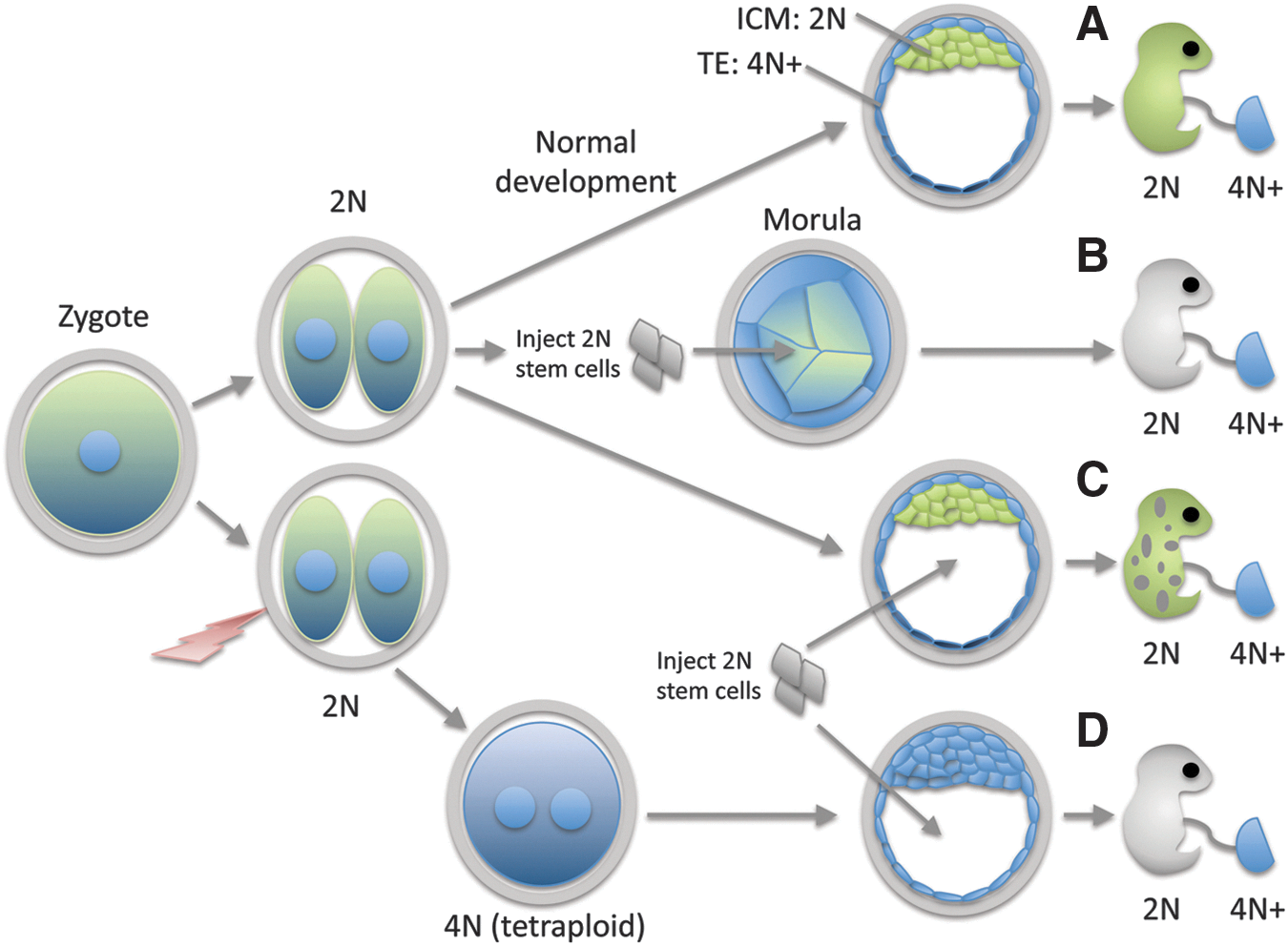
Contribution of diploid and tetraploid cells to development.
Yet, this assertion ignores the fact that in such experimental procedures, stem cells are merely participating in an embryonic process, not originating it. Clearly an unmanipulated embryo will produce a fully formed individual all on its own, without any assistance from stem cells (Fig. 2A). The incorporation of injected cells into a preexisting embryo merely reflects the epigenetic state of pluripotent cells that biases them to contribute to the inner cell mass (ICM) lineage at both morula (Fig. 2B) and blastocyst stages (Fig. 2C).
Similarly, tetraploid embryos are still embryos, although with a significant defect that in most cases proves fatal. However, fully tetraploid mice frequently complete embryogenesis, and occasionally survive to live birth (reviewed in Eakin and Behringer [32]). Similarly, there are reports of live-born, tetraploid humans [33,34], although mosaicism has not been strictly ruled out in most cases. These results demonstrate that the capacity to generate a full developmental program (ie, to progress through the complete sequence of developmental events, despite abnormalities) persists in tetraploid embryos, and that injected diploid stem cells are merely “complementing” the developmental defect arising from the tetraploid state. In these assays (Fig. 2D), the injected stem cells produce “most” (but not all) [32] of the cells in the postnatal body. Importantly, in these experiments, stem cells respond to developmental signals and participate in developmental processes but they do not originate these processes on their own (ie, they are not totipotent), or they would also be capable of generating a full developmental sequence independently, without the “assistance” of either a morula-stage or tetraploid embryo.
Some of the confusion regarding the usage of the term totipotent in this context reflects the arbitrary distinction between structures viewed as “embryonic” (ie, those derived from the ICM and persisting in the postnatal body) and those typically referred to as “extraembryonic” (ie, tissues and organs that have primarily gestational functions; see Table 1). This distinction has a long history in human medical embryology, with the term “extraembryonic” being in common use by the early 1890s [35]. Medically, it is reasonable to distinguish between transient structures that function only during gestation and structures that have functions after birth, since only the latter are the subject of medical intervention (although with the rapidly expanding use of exutero intrapartum therapy, or EXIT, to treat fetal anomalies [36], this distinction may eventually become less medially relevant.).
Yet the term “extraembryonic” does not mean these structures are “nonembryonic.” Structures such as the amnion and placenta are clearly part of the embryo, not part of the mother or of some other entity that coexists with the embryo. Like all embryonic structures, placenta and membranes are (1) generated by the embryo, (2) physically continuous with the embryo, (3) genetically identical to the embryo, and (4) critically required for the function and survival of the embryo as a whole (ie, vital organs of the embryo itself). Consequently, “extraembryonic” structures are best seen as transient structures of the embryo and fetus; that is, organs or tissues that function only during prenatal life.
Many structures have similarly transient functions, including the lungs (functioning only during postnatal life), the thymus (functioning in a greatly diminished capacity at adult stages, compared with its function in the fetus), and the uterus (functioning as a reproductive organ only between puberty and menopause). Due to the purely gestational functions of extraembryonic structures, they are physically discarded at birth, but this is not fundamentally different from the atrophy of the ductus arteriosis (a transient vascular structure that has essential functions only prenatally) in the neonate, or the loss of primary teeth in childhood.
While injected stem cells make minor contributions to placenta and membranes in tetraploid complementation assays (indicating that they are capable of differentiating into these tissues to a limited extent), the failure of stem cells to produce the embryo entirely on their own (including all of the “extraembryonic” organs) indicates that stem cells are not totipotent in either sense, but rather are merely pluripotent, producing only part of the whole embryo in the context of an embryonic process that they do not independently initiate.
Collectively generating a developmental sequence is not totipotency
Scientific authors occasionally equate the ability of groups of cells to collectively generate a full developmental sequence with totipotency of individual cells within the group (Fig. 3). For example, based on the failure to generate live-born mice from aggregates of cells derived from 32-cell-stage embryos [37], the authors assert that blastomere “totipotency” is lost after the fifth round of cell division. Similar results were obtained and similar conclusions were drawn from the observation that “outer” cells of human blastocysts form blastocyst-like structures that initiate expression of ICM-associated genes following reaggregation [38]. Finally, several authors [27,39 –42] have asserted that blastomeres isolated from morula-stage embryos are “totipotent,” because when they are reaggregated with tetraploid cells from a freshly dissociated embryo, either live-born animals or what appear to be normal blastocysts can be produced. Yet, there is no evidence in any of these studies that cells in the aggregate are “totipotent” in either sense of the term; that is, either able to produce all cells of the embryo or to organize these cells into a mature body.
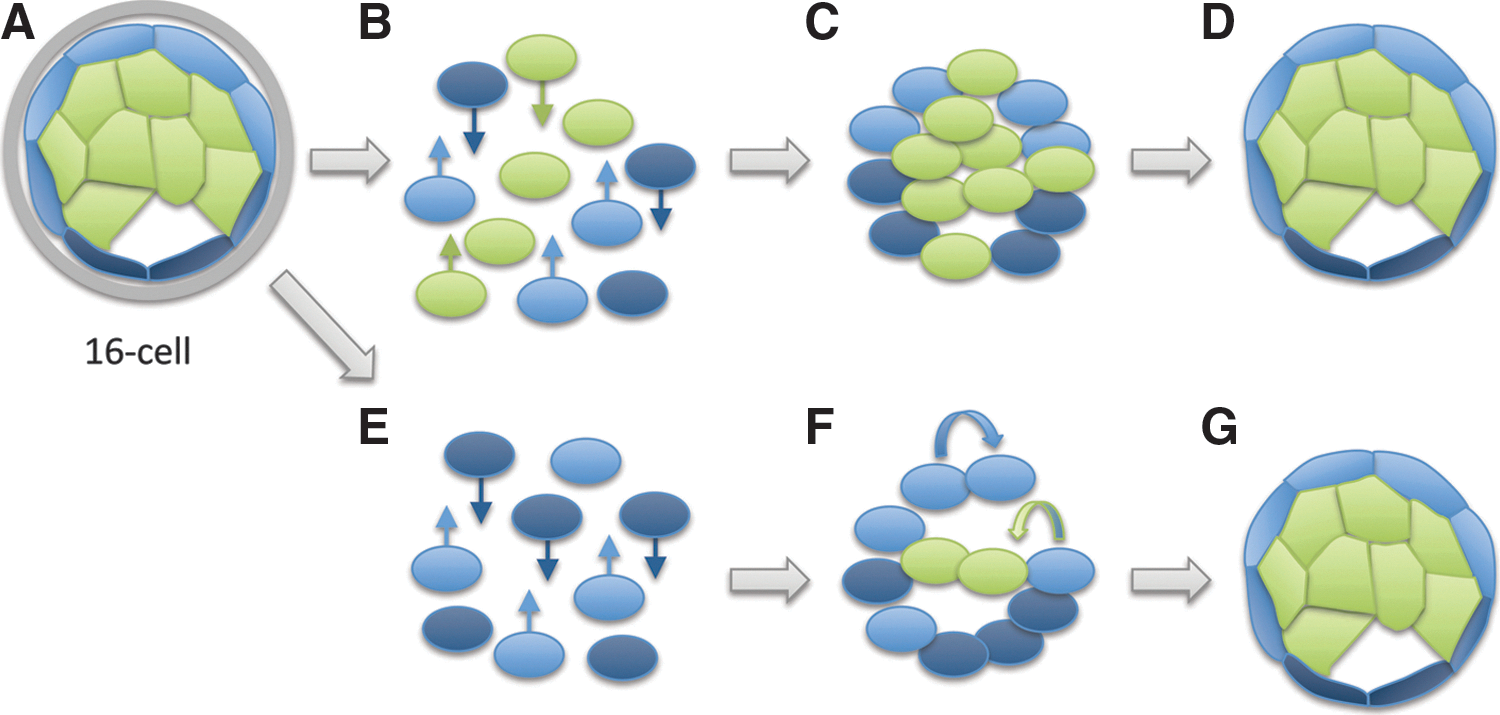
Dissociation and reaggregation of morula-stage blastomeres.
Primary cells isolated from a freshly dissociated embryos are likely to be a mixed population with different developmental capacities (Fig. 3A) and are clearly not equivalent to stem cells [43 –47]. Zygotic transcription commences during the first cell cycle in both mouse [48 –51] and human [52 –59] embryos. While some studies have seen no difference in gene expression between early blastomeres [60], there is evidence from several groups suggesting that as early as the 4–8-cell stage, blastomeres have unique molecular properties [61 –71] that translate into distinct developmental capabilities after dissociation and reaggregation [72,73]. Substantial evidence stretching back over 60 years indicates that when cells with distinct developmental states are aggregated, cell sorting plays a significant role in re-establishing the initial pattern [74,75]. In mice, cell sorting plays a significant role in the segregation of cells within normal 16-cell [76] and 32-cell embryos [77]. Therefore, the cell sorting that has been observed in some studies [37] is likely to reorganize the spatial relationships of blastomeres with limited developmental competency to reconstitute a full embryo (Fig. 3B, E).
Yet even with the most generous interpretation of cell aggregation studies (that cell sorting does not occur and blastomeres at the 16-cell stage contribute randomly to the reconstituted embryo), the ability to respecify as part of a group (ie, the ability to make “any” cell of an embryo) is distinct from totipotency (ie, the ability to autonomously make all cells of an embryo and to organize them into a coherent whole). The significant role of “community effects” in development, first described by Sir John Gurdon in 1988 [78], clearly illustrates that the behavior of cells in groups is distinct from the behavior of the individual cells comprising the group. If the concept of totipotency is not restricted to individual cells and is instead applied to groups of cells that are collectively capable of undergoing development, then all of the cells of any organism at any stage of maturation must be considered “totipotent.” Applying the term “totipotent” in this manner reduces it to a synonym for “organism” and significantly compromises its scientific utility.
An important insight from cell aggregation experiments is that there is more than one way to make an embryo. Embryos produced by either fertilization or by cloning begin as a single-cell zygote that is (by definition) totipotent. Yet, when embryos are reconstituted from collections of morula-stage blastomeres (either diploid or a mixture of diploid and tetraploid), the embryo as a whole must be considered “totipotent” in some sense (ie, it must be considered an organism), but the individual cells within the embryo are not totipotent.
To date, there is no evidence that groups of cells derived from any source other than a freshly dissociated morula-stage embryo can collectively generate a full developmental sequence on their own. Even when freshly isolated ICM [79] or TE [80,81] from blastocyst-stage embryos are transferred to the uterus or other supportive locations, they do not develop as embryos.
Groups of stem cells are also not totipotent. mES lines were first isolated in 1981 [82,83]. Yet, despite thousands of articles that describe the production of stem cell aggregates over the last three decades, there have been no reports of spontaneous generation of embryos from ESCs. Similarly, there have been no reports of generating embryos from a combination of mES and trophoblast stem cells [84]. The failure of aggregated stem cell lines derived thus far to spontaneously form embryos is likely to reflect the fact that stem cell lines have a range of properties [85 –87] that are not identical either to zygotes [88,89] or to cells that have been freshly isolated from embryos [43 –47].
While these negative findings do not strictly rule out the possibility that later-stage blastomeres or pluripotent stem cells “could” combine in some way to generate an embryo (it is logically impossible to prove something from a negative finding), they strongly suggest that collective reconstitution of an embryo from cells of a reaggregated morula-stage embryo (Fig. 3) reflects a unique balance of developmental properties within those freshly dissociated cells—a balance that is not present in cultured stem cell lines or in cells derived from later-stage embryos.
Expression of molecular markers found in early embryos is not totipotency
Scientific authors occasionally conflate expression of molecular markers characteristic of specific embryonic stages with the corresponding developmental capacities of cells at those stages. For example, several studies have determined that most or all mESCs cycle in and out of a unique transcriptional state that shares some elements in common with blastomeres at the two-cell (2C) stage [14,90 –92]. Cells in this 2C-like state lack pluripotency markers normally associated with the ICM, as do human zygotes and embryos at the two-cell stage [88]. The “2C-like” cells also show expanded developmental potential when injected into embryos. The authors present no evidence that isolated 2C-like cells are capable of generating embryos on their own and are careful not to pronounce their 2C-like cells totipotent in the organismal sense, but the repeated observation that the 2C stage correlates with a period in which “blastomeres are totipotent” [14] is strongly suggestive of this unwarranted conclusion and illustrates the confusing usage of the term “totipotent” in the scientific literature. Similarly, the ability of stem cells [3,4] or carcinomas [1,2] to express a molecular marker of “extraembryonic” cell lineages in addition to markers associated with ICM has been taken as evidence of totipotency, despite there being no indication that these cells can initiate a developmental sequence on their own.
Behaving like part of an embryo or merely looking like an embryo is not totipotency
Finally, some scientific authors suggest that the ability of stem cells to replicate limited aspects of normal embryonic development is evidence for totipotency, or something very close to it. For example, a number of studies have shown that ESC aggregates treated with specific signaling molecules exhibit some of the molecular cascades and cell behaviors observed during normal gastrulation [93 –97]. While these studies do not equate stem cells with embryos, several authors conclude that stem cell aggregates are surprisingly “embryo like.” For example, one article states that aggregates “resemble normal embryonic development much closer than previously thought,” exhibiting “an unexpected degree of self-organization” [97]. Yet stem cell aggregates do not resemble organisms so much as they resemble tumors, which can also show a surprising degree of self-organization, producing well-formed teeth [98] and, even in one case, a remarkably normal eye [99].
Similarly, ESC aggregates [100,101] and tumors [102] occasionally generate cystic structures that have some visual similarity to a blastocyst-stage embryo, which has led some authors to ask whether ESCs and tumors might also be embryos [9,103]. Yet many cell types with clearly restricted potency will form such cystic structures, including liver, heart, and cartilage [104]; neurons [105]; fibroblasts [106]; kidney [107]; and umbilical cells [108], indicating that the mere formation of such structures is not sufficient evidence for totipotency.
Confusions Regarding Totipotency in the Nonscientific Literature: Reprogramming “Too Far”
Since the advent of cellular reprogramming [109 –111], the concern that embryos might be inadvertently generated during reprogramming [112] has frequently been raised, with authors asking, for example, “how to ensure that dedifferentiation goes only so far and no further?” [113]. The possibility of producing embryos by reprogramming has been raised by some scientific authors as well [31,103].
The belief that reprogramming could “accidentally” bring a cell into a totipotent state seems to stem from the stubbornly compelling notion that unipotent, pluripotent, and totipotent represent points along a continuum [114] and that reprogramming progressively rewinds development backward toward the beginning. In this view, if reprogramming goes one step beyond pluripotency, then a totipotent zygote will accidentally be produced. Some authors have speculated that this could be done intentionally, generating an “induced totipotent stem” cell, [31] despite the obvious illogic of this assertion; totipotent zygotes are manifestly not capable of self-renewal, and are therefore not “stem cells.”
Reprogramming does not merely “rewind” development
As plausible as this concern may seem, it is not possible for reprogramming to accidently produce a totipotent cell for at least two important reasons. First, reprogramming does not simply “reverse” development, like rewinding an old audiocassette tape reverses the recording; that is, reprogramming does not move a cell backward along the developmental pathway that initially produced its mature state. Instead, reprogramming is more of a single leap from one cellular state to another [115,116]. For example, reprogramming a skin cell into a heart cell [117,118] undoubtedly involves intermediate steps between these two states, but it does not rewind development in any rational sense, since cardiac cells are not part of the developmental history of skin cells.
Pluripotency is as much of a specific cellular state as any other. To reprogram an adult cell to pluripotency, factors are introduced that result in a highly specific cascade of gene expression that is not seen in a totipotent zygote [88]. For example, in human embryos, the transcription factors Nanog and Pou5f1 (also called Oct3/4) are expressed in the ICM and are not found in the zygotes [119 –121]. Consequently, a cell that expresses such factors cannot be identical to a zygote [122].
The state of the cytoplasm
The second important reason a totipotent zygote cannot be accidently produced during reprogramming involves the nature of totipotency—specifically the reasons pluripotent and totipotent are radically different cellular states, even though they occur close together in developmental time. Totipotency is not merely a state of the cell's nucleus; it also requires a very specific type of cellular cytoplasm that is a critical component of totipotency.
At this time, the only known totipotent cytoplasm is produced by an oocyte and contributed to the embryo at fertilization. The fact that oocytes produce the cytoplasmic factors that are required for an embryo to be totipotent is the reason oocytes are used for cloning. During cloning, oocyte components can, over the first several days of development, induce an adult nucleus to assume a state similar to that of a normal embryo (ie, a state that is capable of driving a complete developmental sequence), mimicking the processes that naturally occur after sperm-egg fusion [123]. If a competent nuclear state is achieved, then the oocyte-specific components in the cytoplasm work with the reprogrammed nucleus to produce an embryonic pattern of development, despite the significant differences observed between cloned embryos and embryos derived from fertilization [124 –127].
Oocytes are highly structured cells that are uniquely produced by the complex process of oogenesis, which involves a characteristic sequence of gene activation [128] that is distinct from the pattern observed in the maternal pronucleus after fertilization or following zygotic gene activation [58,59]. Oogenesis also requires information from other cells in the ovary [129 –131]. Moreover, recent work clearly documents multiple oocyte-derived components that are essential for mammalian embryonic development (reviewed in Li et al. [132] and Matzuk and Burns [133]). For example, oocyte-expressed Ooep [134], PADi6 [135], Nlrp5 [136], Ecat1 [137], and Tle6 [138] are required for the normal function of a critical subcortical cytoplasmic complex, with loss of any of these genes resulting in embryo lethality at the two-cell stage. Maternal expression of Kdm1B [139], Dmap1 [140], Dppa3 [141], and several others is required for correct DNA methylation and maintenance of genomic imprinting, with maternal gene deletions resulting in death at early embryonic stages. Finally, maternally supplied Brg1 [142] and Brwd1 [143] are required for zygotic gene activation, with loss of these genes resulting in arrest at the two-cell stage.
Simple reprogramming of a somatic nucleus does not establish the cytoplasmic components required for totipotency and these factors are not produced during normal embryonic development; that is, the transcriptomes of oocytes and early embryos are clearly distinct [58,59]. Without the required cytoplasmic components contributed by oocyte, there can be no zygote—regardless of the state of the nucleus [19].
In other words, even if the nucleus of a somatic cell is fully reprogrammed to be identical in every respect to that of a zygote (a state that is distinct from a pluripotent stem cell and one that would require different reprogramming factors), it would still not be totipotent, because it lacks the nongenetic factors (proteins, RNA, miRNA, and specific macromolecular complexes) that are critical components of totipotency. Conversely, if a nucleus from a bona fide zygote produced by fertilization was transferred to a differentiated cell (eg, a muscle cell), it would not continue to be a zygotic nucleus in this new location. It would be (or would rapidly become) a muscle cell nucleus that functions in the manner specified by the muscle cell cytoplasm [144].
The requirement for totipotent cytoplasm in order for a cell to actually be a zygote is precisely the reason that totipotency persists for such a short time in development. In most mammals, only the first two cells of the embryo remain totipotent, that is, able to generate a complete embryo on their own when separated [145 –147], perhaps in part because cells no longer have sufficient mass to continue development [148,149]. In rare cases, totipotency is preserved until the four-cell stage [150], with there being one reported case of a child born after transfer of a four-cell embryo in which only one of the blastomeres was judged to be alive [151]. There has also been one report in pigs of totipotency persisting until the eight-cell stage [152]. But there is no evidence for totipotent cells persisting beyond this stage. As soon as a cell has only a subset of the specialized cytoplasmic components contributed by the oocyte, it is no longer totipotent.
The cell fate restrictions observed in mice and other species from the two-cell stage onward are likely to reflect inheritance of required oocyte-derived components as well as cell–cell interactions and stochastic events [72,153 –156]. There is clear evidence from multiple laboratories that by the four- to eight-cell stage these two processes have resulted in blastomeres that have distinct molecular, functional, and developmental properties [61 –71,157,158], indicating that from the four-cell stage onward, mammalian development is mosaic to some extent.
The requirement for totipotent cytoplasm in no way denies the clearly regulative aspects of mammalian development that depend on cell–cell signaling, cell interaction, and random events. Neither does it imply that inheritance of maternal factors is the only mechanism underlying cell differentiation in mammals, or that such factors must be differentially inherited by specific blastomeres, although differential inheritance is possible mechanism underlying early lineage restrictions. Rather it suggests that the distinction between regulative and mosaic development is a false dichotomy [159]. Cell fate decisions are produced by a continuum of overlapping and redundant mechanisms [76], with oocyte-derived factors clearly playing a critical role in the developmental competency of early blastomeres [160 –164].
Is it possible to make a cloned embryo from an adult cell by reprogramming?
Reprogramming can generate pluripotent stem cells because it initiates a chain of events that brings an adult cell nucleus into a pluripotent state and this “converted” nucleus subsequently produces the non-nuclear factors that are required to actually be a pluripotent stem cell. Yet totipotent zygotes are unusual, because unlike stem cells or any other cells in the body, zygotes do not produce the highly specialized cytoplasm they require on their own. Rather they inherit this specialized cytoplasm from a very different kind of cell with a very different nuclear state, that is, from the oocyte. Once a cell nucleus has the configuration of a totipotent zygote, it does not produce the same factors produced by the oocyte.
This does not mean that it would be impossible to make an embryo by reprogramming, but it does mean that it cannot happen “accidently.” And converting an adult cell into an embryo using reprogramming (making an “induced totipotent cell”) would be difficult to accomplish, even intentionally (Fig. 4).
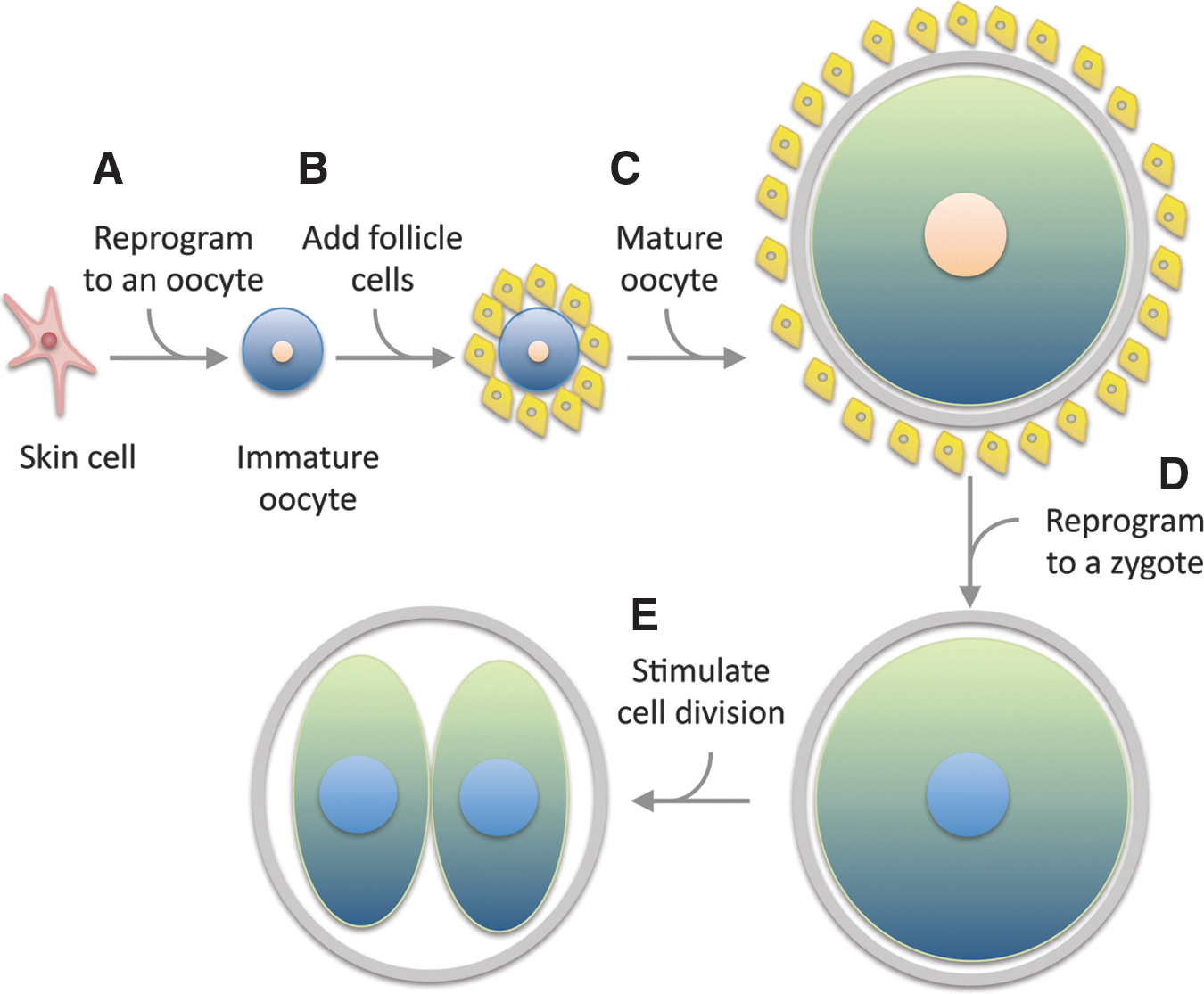
The multiple steps to reprogram a mature cell to a zygote cannot happen unintentionally.
To convert an adult cell into a zygote, it would first have to be reprogrammed to become a cell that is capable of providing the factors that are normally generated during the process of oogenesis. The simplest way of accomplishing this would be to reprogram the adult cell into an immature oocyte (a distinct state from a pluripotent stem cell that would require different reprogramming factors). The immature oocyte would then have to be provided with all of the cell interactions and ovarian factors required for it to become a mature oocyte. During this process, the normal epigenetic reprogramming and meiotic divisions that occur as part of oogenesis [123,165] would have to be suppressed in order to preserve the nucleus in a state that is capable of driving human development (this may not be technically or even logically possible.). Once this unnaturally suppressed oocyte had been made, the nucleus would again have to be reprogrammed to a zygotic state, a significant remodeling that normally reflects factors derived from both sperm and egg [21,123,166]. If all of this could be achieved, then the “secondarily reprogrammed” totipotent cell would have to be activated to begin the process of development. Then, and only then, would a cloned embryo be produced from an adult cell by reprogramming. And this could hardly happen “by accident.”
Twinning
It is sometimes asserted that because twinning can occur by splitting of a blastocyst-stage embryo (Fig. 5), this “proves” that the blastocyst contains a mixture of totipotent and pluripotent cells [167 –169]. This view raises the concern that because pluripotent stem cells are similar to cells of the ICM, stem cell cultures could also contain a mixture of pluripotent and totipotent cells.
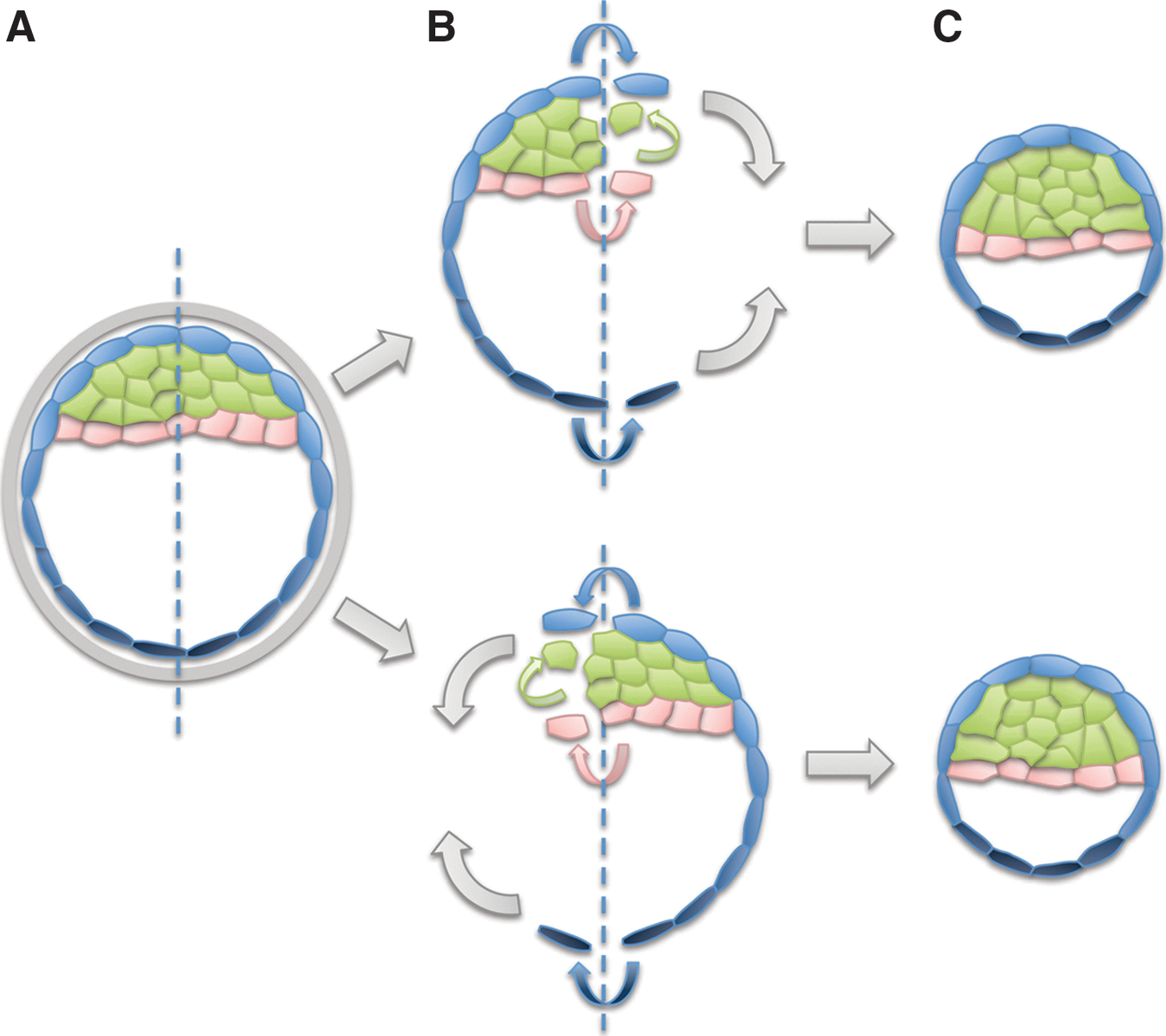
Twinning at the blastocyst stage does not require totipotent cells to be present.
Twinning can potentially occur by a number of different mechanisms (a topic that has recently been re-examined in an excellent critical review; [170]). However, there is no scientific evidence to date that twinning at the blastocyst stage involves totipotent cells, and multiple studies have concluded exactly the opposite; that is, that blastocyst cells are quite unlike totipotent zygotes on multiple parameters [43 –47,85 –87] and that blastocyst cells are completely incapable of producing a whole embryo when isolated from each other [145 –147], instead producing only stem cell lines. These observations argue strongly that twinning does not involve totipotent cells, but rather relies on some other developmental mechanism.
The simplest way twinning could occur at the blastocyst stage is by the process of “regulation,” first described by Driesch over 100 years ago [171]. Regulation does not mean that when an embryo is split, cells revert to totipotency and start the process of development all over again. It simply means that a split embryo remains an embryo. Just as an individual who loses a limb continues to be a human being, although a damaged one, a blastocyst-stage embryo that has lost half of its cells is still an organism. And one of the characteristics of organisms is that they repair injuries. Yet unlike adult human organisms, who have limited ability to regenerate lost tissue, human organisms at the embryonic stage are very good at regenerating missing parts (Fig. 5).
When an embryo is split, each half can (in some cases) become a “demi-embryo” that proceeds with development [172 –175]. Direct observation indicates that in such demi-embryos, the ratio of cells in the ICM and TE is either maintained or restored by cell proliferation [176 –178]. Lineage analysis indicates that cells of the TE or ICM largely contribute cells to their own lineage (Fig. 5B), just as they do in normal development [179 –181]. There is no evidence for totipotent cells producing most or all of the structures of the twin after embryo splitting.
Similar results are seen for even more catastrophic injuries. When the cells of the ICM are isolated from blastocyst-stage embryos, the outer cell layer [either TE or primitive endoderm (PE)] can be partially regenerated in some cases. Detailed analysis of these imperfectly restored embryos shows that the regenerated tissues are not produced by “totipotent” cells within the ICM, but rather by committed TE or PE cells that were isolated along with the ICM [182,183].
After blastocyst splitting, embryonic development is not reinitiated (Fig. 5C), but rather proceeds in synchrony with normal (un-split), sibling embryos [184]. There is no recapitulation of the events of early development. The embryo repairs itself by the same kind of process an adult human uses to repair injury; cells in each of the embryo's specialized tissues replace the specialized cells that have been lost, and once the damage has been repaired, development proceeds from the blastocyst stage forward without returning to a zygote-like state. Twinning appears to be another example of a community effect, whereby a group of cells has properties that exceed the potency of any of the individual cells comprising the group. Importantly, twinning at the blastocyst-stage provides absolutely no evidence for the presence of totipotent cells within the blastocyst. Therefore, the ability of embryos to generate twins raises no credible concerns about the potency of stem cells derived either from reprogramming or from blastocyst-stage embryos.
Conclusions
Totipotent zygotes are distinct from pluripotent stem cells or tumors because they can originate development. The ability to both produce all cell types and to organize them into a coherent body plan is the defining feature of an organism [5,6] and also the strict definition of totipotency. Misapplication of this term in the scientific literature creates artificial controversy over areas of research that are ethically unproblematic. In the interest of both scientific accuracy and public education, scientists should confine the use of the term totipotent to embryos, and apply a rigorous scientific standard to defining totipotency; the ability of an isolated cell to mature into a fully formed individual when placed in a supportive environment (for mammals, a uterus). To describe cells that produce all the derivatives of the zygote, yet are not capable of undergoing development (eg, hESCs and embryonal carcinomas), either new terminology, such as “plenipotent,” or a qualified use of “pluripotent” would be preferable to a misleading application of the term “totipotent” to cells that are not organisms.
Clearly, there is more than one way to make an embryo. Naturally conceived human embryos are produced by fusion of sperm and egg in a woman's reproductive tract, a process that can be replicated, although imperfectly [185 –187], by in vitro fertilization. Recent work has shown that human embryos capable of surviving to the blastocyst stage can be produced by cloning [188]. All three of these approaches require oocyte cytoplasm to generate a totipotent one-cell embryo.
Importantly, all other ways of “reconstituting” an embryo start with an embryo produced by one of these three methods (natural fertilization, in vitro fertilization, or cloning). Injecting pluripotent cells into an existing morula, blastocyst, or tetraploid embryo generates a chimeric animal, with injected cells contributing in varying degrees to the postnatal body (Fig. 2). Under some conditions, freshly dissociated early embryos (up to the 16-cell stage) can be reaggregated and produce live-born animals (Fig. 3). Splitting an existing embryo can result in the regeneration of two demi-embryos (Fig. 5). Yet none of these methods produce an embryo—they merely reconfigure an existing embryo or contribute additional cells to an ongoing embryonic process.
Ultimately, we may discover additional ways of constituting a full organism from cells that are not themselves totipotent or identify new ways of producing a totipotent cell. Yet in all cases (both actual and theoretical), the unique capacity of an organism to undergo development clearly distinguishes an embryo from a cell that is not totipotent. And it is the capacity of an organism to generate a complete developmental program that warrants serious ethical concern. We do not owe any particular ethical consideration to human gametes, skin cells, or stem cells. Yet, if human cells are manipulated so that they participate in the development of an existing embryo (eg, by tetraploid complementation) or are used to generate a human embryo de novo (eg, by fertilization or cloning), then the embryo merits substantial ethical consideration due to the fact that it is a human organism, that is, a human being.
Footnotes
Acknowledgments
The author thanks Drs. H.J. Yost and D. Prentice for suggestions on the article and figures.
Author Disclosure Statement
No competing financial interests exist.