
Abstract
In mammals, the sensory hair cells and auditory neurons do not spontaneously regenerate and their loss results in permanent hearing impairment. Stem cell therapy is one emerging strategy that is being investigated to overcome the loss of sensory cells after hearing loss. To successfully replace auditory neurons, stem cell-derived neurons must be electrically active, capable of organized outgrowth of processes, and of making functional connections with appropriate tissues. We have developed an in vitro assay to test these parameters using cocultures of developing cochlear explants together with neural progenitors derived from human embryonic stem cells (hESCs). We found that these neural progenitors are electrically active and extend their neurites toward the sensory hair cells in cochlear explants. Importantly, this neurite extension was found to be significantly greater when neural progenitors were predifferentiated toward a neural crest-like lineage. When grown in coculture with hair cells only (denervated cochlear explants), stem cell-derived processes were capable of locating and growing along the hair cell rows in an en passant-like manner. Many presynaptic terminals (synapsin 1-positive) were observed between hair cells and stem cell-derived processes in vitro. These results suggest that differentiated hESC-derived neural progenitors may be useful for developing therapies directed at auditory nerve replacement, including complementing emerging hair cell regeneration therapies.
Introduction
S
Several laboratories have now explored the possibility of using stem cells to replace auditory neurons, which progressively degenerate after hearing loss [3 –12]. Of these studies, two have reported the use of human stem cells for auditory neuron replacement [11,12], and this will eventually be critical for translation of this therapy into clinical practice. In developing a cell replacement therapy for hearing loss, donor stem/progenitor cells must also be electrically active and capable of organized outgrowth of processes, including forming synapses on appropriate cells/tissues in the peripheral and central auditory system [13,14]. An additional important aspect of correct targeting and innervation of the host tissue from the donor cell is that donor cells are prespecified to an appropriate progenitor cell type.
During embryonic development, auditory neurons are derived from the otic placode; the earliest morphologically visible event in the development of the inner ear [15]; this ectodermal thickening develops in close proximity to the hindbrain. The development of neurosensory progenitors from the otic placode is thought to follow the same molecular principles of brain development, including the combined interaction of diffusible signals, including fibroblast growth factors [basic fibroblast growth factor (bFGF); 3, 8, and 10), Shh, Wnts, and bone morphogenetic proteins [15 –17]. The interaction between these four factors produces a patterning gradient that induces target cells into otic placode derivatives [16]. Moreover, the primary auditory neurons of the mammalian cochlea arise from the medial ectoderm of the otic placode [18], where they express Eya1 [19,20], Six1 [20], Pax2 [21 –23], and Islet1 [23]. These neurons then undergo sequential expression of neurog1, neurod1, Brn3a, and GATA3, before delaminating from the otocyst and comprising the auditory neurons of the inner ear [24,25]. Similarly, the neural crest cells arise from an adjacent region of the dorsal neural tube and delaminate laterally toward the otic placode where they develop in close association. Moreover, auditory neural progenitors share similar properties to neural crest progenitors, particularly those fated to become sensory neurons and their progenitors share common molecular and phenotypic profiles during development [26], including some common genes required for their specification, such as Sox9 [27,28], Neurog1, NeuroD1, Brn3a [29,30], and NTrk receptors [31,32]. Thus, neural crest-like cells may also be suitable for deriving sensory neurons for auditory neuron replacement, particularly when employing neurosphere (NS) assays, which innately produces a mixed population of neural progenitor cells.
Neural crest-like cells can be differentiated from the human embryonic stem cell (hESC) NS, using combined treatment with mouse embryonic fibroblasts (MEFs) and the small ROCK/Rho inhibitor Y27632 [33]. Importantly, treatment with Y27632 was shown to result in significant delamination of neural crest-like progenitors away from the peripheral edge of the NS [33]. Given the heterogeneous nature of this differentiation assay and the close development of the neural crest and otic placode in situ, we examined how neural progenitors derived from this differentiation protocol (treated NS) behaved when cocultured with early postnatal cochlear explants [14]. Untreated NS were set up concomitantly and served as controls.
Materials and Methods
Stem cell culture, NS formation, and NS treatment
The H9 (WA-09; WiCell) and ENVY hESC lines were cultured in a 20% serum-containing medium on mitomycin-C-treated MEFs and passaged weekly by mechanical dissociation [34]. The ENVY hESC line was chosen for use in the cochlear explant cocultures, given the high levels of expression of the green fluorescent protein (GFP) in all differentiated progeny [35], thereby enabling rapid discrimination of hESCs cellular derivatives from endogenous tissues. In addition, the ENVY cell line is particularly useful for neuroscience experimentation in general, as ENVY-derived neurons express high levels of GFP in both somata and processes, enabling visualization and tracing of neural processes in vitro and in vivo.
For neural induction of H9 and ENVY hESCs, 500 ng/mL recombinant noggin (R&D Systems) was added to the hESC medium at the time of colony transfer onto MEFs, and cells were cultured for 14 days without passage. After noggin treatment, colonies were mechanically transferred to individual wells of a nonadherent 96-well plate to allow NS formation (Stage 1, Fig. 1). NS were cultured in suspension in Neurobasal media (Invitrogen) supplemented with the epidermal growth factor (EGF) and bFGF (20 ng/mL each; R&D Systems) for a further 4 days (Stage 2, Fig. 1). These steps have been described in detail by Dottori and Pera [34]. Subsequent differentiation of NS toward neural crest-like progenitors (i.e., treated NS; Fig. 1) was carried out as previously described by Hotta et al. [33], but briefly, after 4 days of NS formation (Day 18, Fig. 1), NS were plated onto a feeder layer of mitomycin-treated MEFs in Neurobasal media supplemented with bFGF and EGF (20 ng/mL of each) for 24 h. After 24 h (Day 19, Fig. 1), the medium was replaced with a fresh medium containing Y27632 (25 mM; Sigma Aldrich). Following 48 h incubation (Day 21, Fig. 1), either whole cochlear explants or denervated cochlear explants were then dissected and set-up on organotypic membranes and cocultured with either NS (untreated NS) or MEF/Y27632-treated NS (treated NS) for a further 5 or 10 days in vitro (DIV) (Stage 3, Fig. 1) as described below.
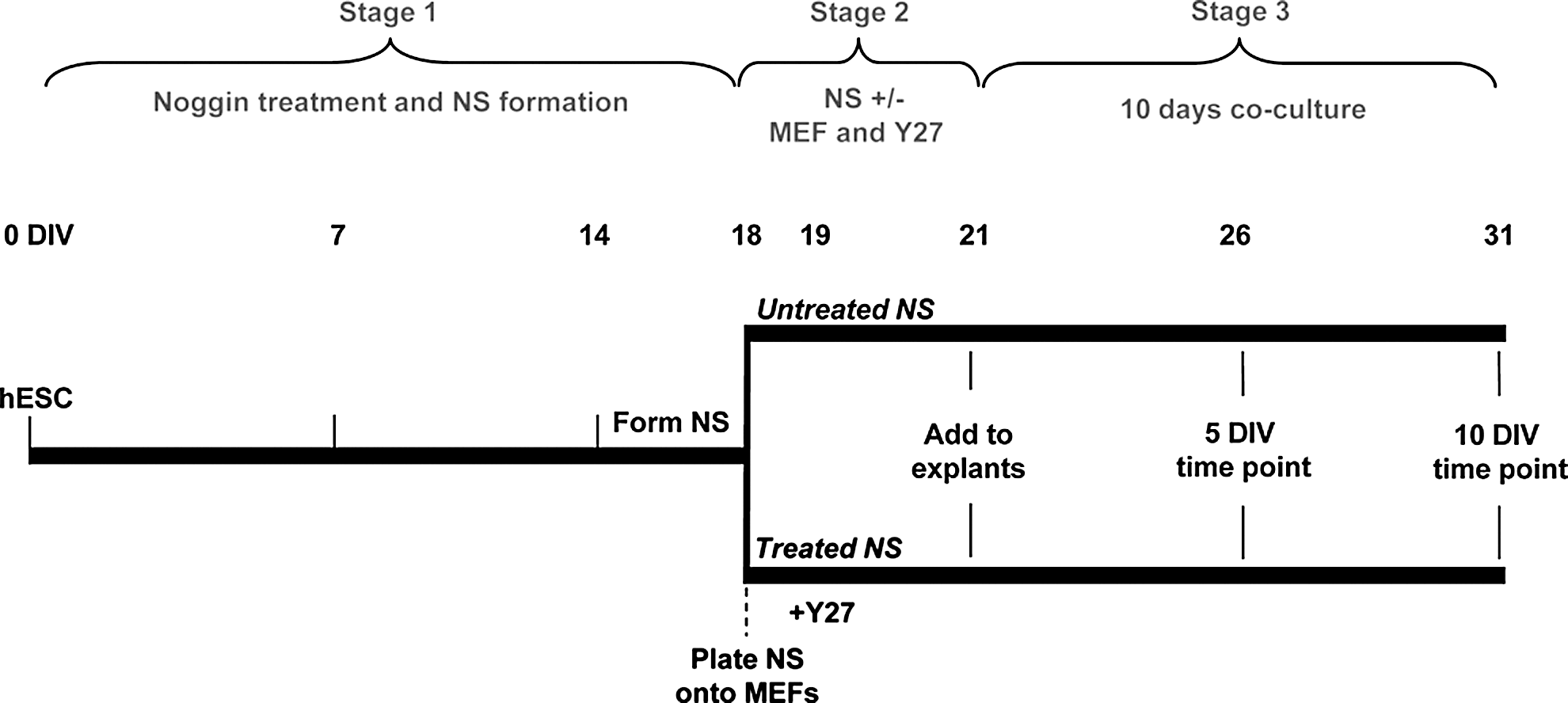
Timeline of in vitro differentiation and coculture. The timeline summarizes the 3 stages of differentiation and coculture used in the study. hESC, human embryonic stem cells; NS, neurosphere; MEF, mouse embryonic fibroblasts; Y27, ROCK/Rho kinase inhibitor Y27632; DIV, days in vitro.
In vitro electrophysiology
For in vitro electrophysiology recordings, Day 18 NS (Fig. 1) were plated onto small round glass coverslips (10 mm; Lomb Scientific) and grown for a further 10, 14, 21, 28, or 32 DIV at 37°C, 10% CO2. Coverslips were transferred to a recording chamber fitted to an AxioExaminer D1 microscope (Carl Zeiss) and equipped with a 40×water-immersion objective lens. Cells were morphologically identified using Dodt optics and a monochrome CCD camera (Spot RT SE18; Diagnostic Instruments), and superfused at 1–2 mL/min with solution of the following composition (in mM): 137 NaCl, 5 KCl, 10 HEPES, 1 MgCl2, 2 CaCl2, and 10 glucose (pH 7.35; 300–305 mOsmol/kg). Whole-cell patch-clamp recordings were made at room temperature using borosilicate microelectrodes (2–6 MΩ; 1.0 mm outer diameter; 0.58 mm internal diameter filled with an internal solution containing (in mM) 115 K-gluconate, 10 HEPES, 7 KCl, 0.05 ethylene glycol tetraacetic acid, 2 Na2ATP, 2 MgATP, and 0.5 Na2GTP (pH 7.3; 290–295 mOsmol/kg). All chemicals were purchased from Sigma-Aldrich unless otherwise indicated. Tetrodotoxin (TTX; Alomone), 4-aminopyridine (4-AP), and tetraethylammonium (TEA) were diluted daily to final concentrations in the bath perfusate and administered by superfusion via a gravity-fed system. Signals were recorded with a MultiClamp 700B amplifier (Molecular Devices), data acquisition system (Digidata 1440A; Molecular Devices), and AxoGraph X analysis software (AxoGraph Scientific). Results are presented as mean values±standard error of the mean.
Early postnatal cochlear explant dissections and cocultures
Sensory explants containing both hair cells and auditory neurons were dissected and cultured from early postnatal rat pups aged P3/P4, using techniques previously described [36,37]. All explants were grown on 0.4-μm organotypic membranes (Millipore; 2–3 explants per membrane) in Neurobasal A media (Invitrogen) containing B27 and N2 supplements (Invitrogen) in combination with 50 ng/mL of each of the human recombinant neurotrophins: the brain-derived neurotrophic factor (Millipore) and neurotrophin 3 (Millipore). Explants were maintained at 37°C, 10% CO2 for either 5 or 10 DIV as illustrated in Fig. 1 Stage 3 (26 days=5 DIV; 31 days=10 DIV), before being fixed and processed for immunocytochemistry. In an additional set of experiments, cochlear explants were denervated to dissect the hair cells from the neural tissue in the explants, using microdissection techniques previously described [38,39]. In these experiments, the hair cells alone were cocultured with Y27632-treated NS for 10 DIV to investigate the formation of pre- and postsynaptic terminals between stem cells and hair cells. These cocultures constitute our synapse-forming assay and were fixed and processed for immunocytochemistry as described below. It should be noted that hair cell numbers rapidly decline after 12 days in organotypic culture; therefore, longer term coculture investigations of this nature (similar to that performed by [40]) are not viable using the described assay. This is not limited to our laboratory, but rather a reflection on the fastidious nature of these sensory cells and the difficulty in culturing them in vitro. This is substantiated by the lack of any other published studies to date culturing cochlear hair cells for longer than 2 weeks in vitro [8,11,39,41]. All experimentation was repeated in triplicate, and all animal experimentation was conducted in accordance with the Royal Victorian Eye and Ear Hospital Animal Research and Ethics Committee Guidelines, project approval number 09/182AR: B. Nayagam.
Immunocytochemistry
Endogenous GFP expression in the hESC line, ENVY, facilitated their detection and imaging in live cultures. For fixed cell cultures, double-and triple-labeling experiments were performed throughout the study to distinguish the sensory hair cells (Myosin VIIa-positive) from the hESC-derived cells (GFP-positive) and the neurons (including both stem cell-derived neurons and endogenous auditory neurons), which were detected using antibodies against peripherin and neurofilament-M (NFM; 160 kDa) and neurofilament-H (NFH; 200 kDa). Additional antibodies were employed to investigate the presence of the cranial placode marker Pax2 and the sensory neural transcription factor Brn3a in both the early treated and untreated NS. All antibody concentrations were determined individually, and negative controls (without primary antibody) eliminated all immunoreactivity. Details of all primary antibodies used in the study, including optimal concentration and manufacturer's details are as follows: Mouse anti-GFP (1:200; Millipore MAB3580), Rabbit anti-Pax2 (1:200; Covance PRB-276P), Mouse anti-Brn3a (1:1,000; Millipore MAB 1585), Rabbit anti-synapsin 1 (1:200; Invitrogen A6442), Mouse anti-PSD95 (1:200; Millipore MAB1596), Rabbit anti-MyosinVIIa (1:100; Proteus PTS25-6790), Rabbit anti-Peripherin (1:200; Millipore AB AB1530), Chicken anti-Neurofilament 160 kDa (1:1,600; Millipore AB5735), and Chicken anti-Neurofilament 200 kDa (1:800; Millipore AB5539). All secondary antibodies were sourced from the Alexa Fluor dye series (Invitrogen) and used at the following concentrations: Goat anti-mouse IgG1 (1:500), Goat anti-rabbit IgG (1:500), Goat anti-mouse IgG2a (1:500), and Goat anti-chicken IgG (1:500) standard immunocytochemical procedures were used to fluorescently label fixed cells. Briefly, cells were blocked in a solution of 0.1% triton-X in phosphate-buffered saline (PBS) containing 2% goat serum for 1 h at room temperature before primary antibodies were added at the desired concentration; 200 μL per well for 2 h at room temperature, and then overnight at 4°C. The following day, cells were thoroughly rinsed in a blocking solution and the relevant secondary antibodies added at the concentrations described for 2 h at room temperature with gentle rotation. All cells were then rinsed 3 times for 10 min in PBS and mounted with the ProLong Gold antifade reagent containing the nuclear stain 4′6-diamindo 2-phenylindole (DAPI) (Invitrogen). For the synapse-forming assay, ProLong Gold antifade mount (Invitrogen) was used. Slides were dried at room temperature overnight and the cover glass edges sealed with nail varnish the following day.
Image analysis and quantification
Fluorescence photomicrographs of both NS and cocultures were taken using an LSM 510 META confocal scanning laser system attached to a Zeiss AxioImagerZ1 microscope and image analysis was performed using the LSM Image Browser software. To quantify the growth of treated and untreated hESC-derived processes into the explant, we counted the number of neurites that were observed growing into the tissue at a distance equal to half the radius (0.5r) of the NS present in the coculture. This enabled us to control for slight differences in NS size (and therefore number of cells in the cocultures). All measurements of neurite growth were performed blindly and data were statistically analyzed using SigmaPlot 11 software, to identify significant differences in the numbers of positively labeled cells between treatment groups. Statistically significant effects were detected using a Kruskal–Wallis one-way analysis of variance on Ranks, with the Dunn's All Pairwise Multiple Comparison Procedure to isolate the groups that differed.
Results
Both treated and untreated NS express Pax2, Brn3a, peripherin, and neurofilament proteins
We have previously described a robust method for deriving neural crest-like progenitors from the GFP-positive ENVY hESC line [35], by culturing hESC-derived NS on a feeder layer of MEFs in the presence of the small molecule Rho/ROCK inhibitor, Y27632 [33]. Neural crest-like progenitors derived using this method (i.e., treated NS) were shown to differentiate into peripheral neuronal lineages both in vitro and in vivo [33]. To further characterize the neurosensory cell phenotypes produced from this differentiation assay, we examined expression of sensory neural proteins in both treated and untreated NS after various periods of differentiation (Fig. 2). Expression of the cranial placode marker, Pax2, was detected at Day 21 of hESC differentiation (Fig. 2A, C), followed by expression of the sensory neural transcription factor Brn3a (Fig. 2E, G) and NFM (Fig. 2F, H) at Day 26. Both peripherin (Fig. 2I, K) and NFH (Fig. 2M, O) were detected at 31 DIV. Similar expression of Pax2, Brn3a, peripherin, NFM, and NFH was observed in both treated and untreated NS examined at the same time points, suggesting that sensory neural precursors are present both in treated and untreated hESC-derived NS. To date, there is no specific marker that defines auditory neuron progenitor subpopulations that gives rise to myelinated versus unmyelinated neurons. Thus, it is difficult to discern in our in vitro cultures whether the human stem cell-derived Brn3a+ progenitors potentially represent myelinated and/or unmyelinated subpopulations of auditory neurons. Both auditory neural subtypes express the markers Pax2, Brn3a, peripherin and neurofilament proteins.
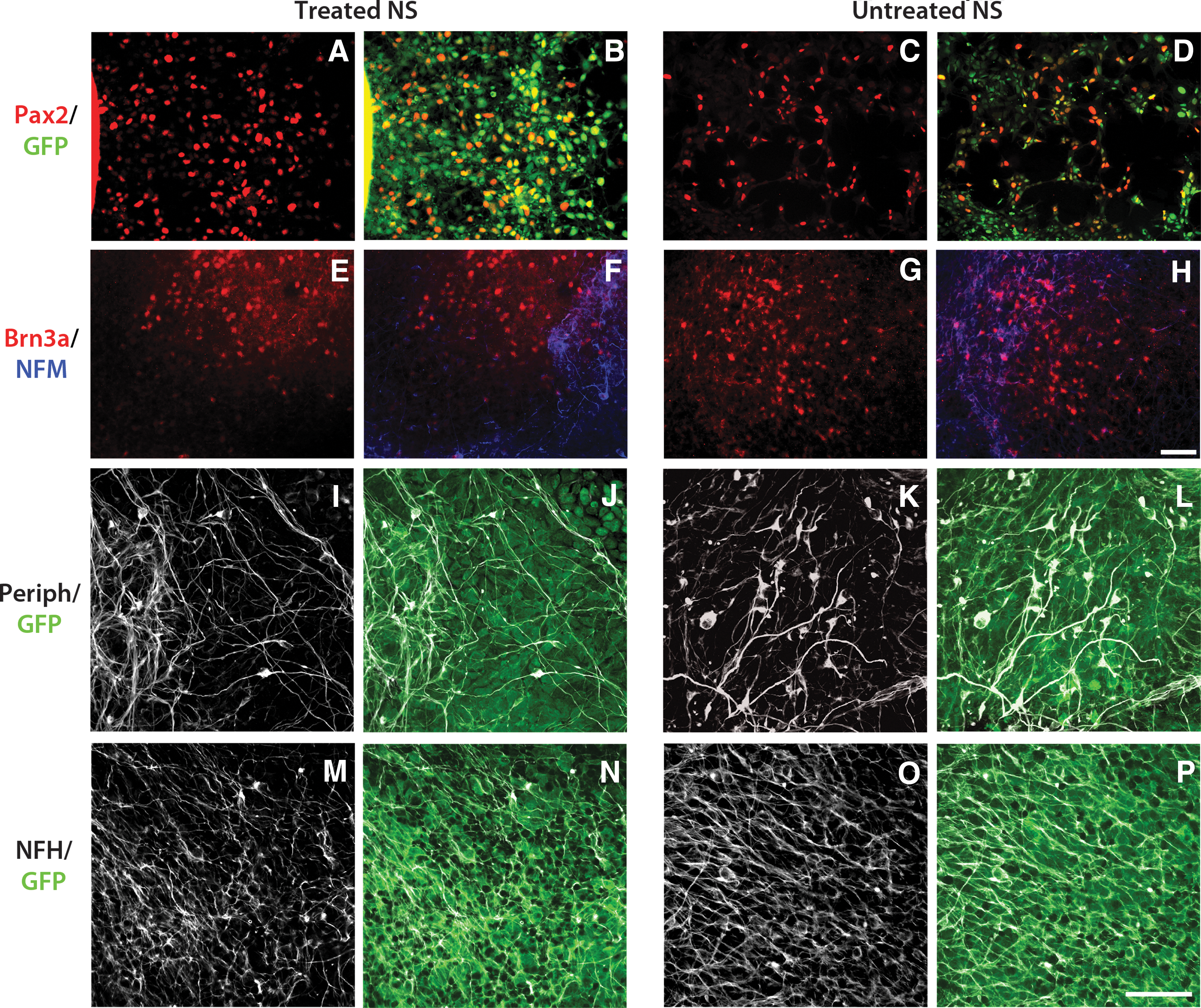
Immunocytochemical labeling of treated and untreated NS cultures. Both treated and untreated NS were immunolabeled for Pax2, [red;
Stem cell-derived neurons are electrophysiologically active
To evaluate the functional potential of neurons derived from treated NS, we examined their electrical activity using whole-cell patch clamp recordings after 31, 35, 42, and 53 DIV. Recordings were made from cells displaying a patent bipolar neuronal morphology (n=33; Fig. 3A). The mean resting membrane potential was −49.6±1.3 mV. All cells examined were capable of generating an action potential in response to membrane depolarization (+20 to +190 pA, 300 ms duration). All cells exhibited a phasic firing pattern (Fig. 3B, F). The mean maximum number of action potentials fired was 1.2. No spontaneous activity or repetitive firing was observed. Mean action potential half-width and time to peak were 3.0±0.2 ms and 39.1±3.3 ms, respectively, at threshold, and 2.7±0.3 ms and 5.3±0.3 ms in response to supra-threshold stimulation (+190 pA). Recordings under voltage-clamp conditions show a fast-activating, rapidly inactivating inward current evoked during membrane depolarization that is sensitive to 1 μM TTX, a selective blocker of voltage-gated sodium channels (Fig. 3C, E). The TTX-sensitive sodium current was followed by a sustained outward current (Fig. 3C), which could be attenuated by a combined application of the potassium channel blockers 10 mM TEA and 1 mM 4-AP (Fig. 3D). A comparison of recordings made at 31, 35, 42, and 53 DIV revealed the same phasic firing profile in response to membrane depolarization (Fig. 3F).
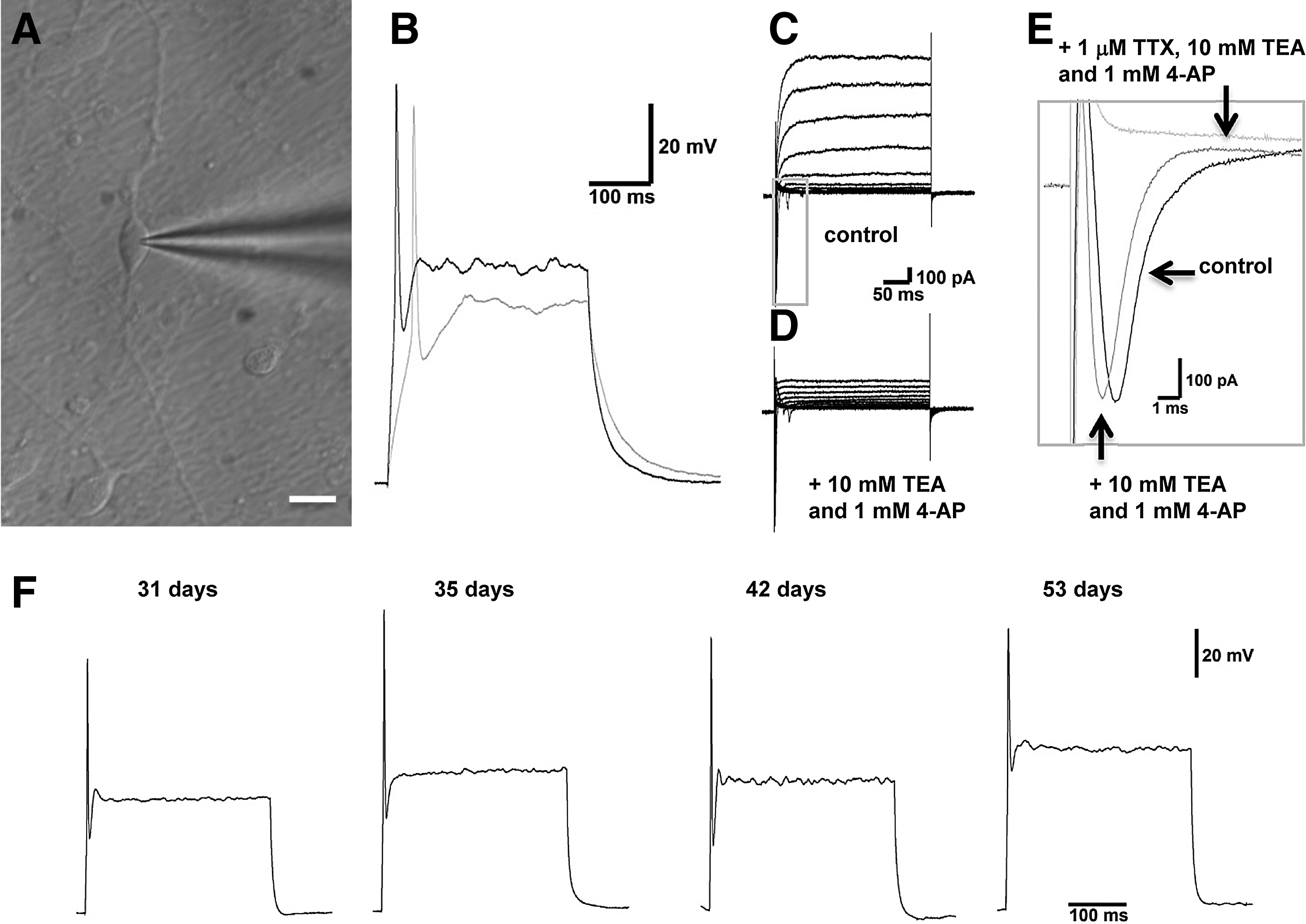
Electrical activity in hESC-derived neurons. hESC-derived neurons
Treated NS grow into the developing cochlear epithelium
Given the similar levels of expression of Pax2, Brn3a, and neuronal cytoskeletal markers in both treated and untreated NS (Fig. 2), we sought to determine whether each of these differentiated systems had the capacity to grow into early postnatal cochlear explants (Stage 3, Fig. 1). We tested this by coculturing 21-day differentiated hESCs (Stage 2, Fig. 1) with P3/P4 cochlear explants. We observed growth of stem cell-derived (GFP-positive) processes into the explant after 10 DIV (n=6, Fig. 4), including extension of processes toward the sensory hair cells located on the peripheral edge of the explant (Fig. 4D, E, G, I). Under a higher magnification, these fine stem cell-derived processes were observed growing on top of the endogenous peripheral processes in the cochlear explant after 10 DIV (refer to Fig. 5). These findings prompted us to investigate whether this process growth was specific to treated NS only, or rather whether it was a property of all hESC-derived NS.
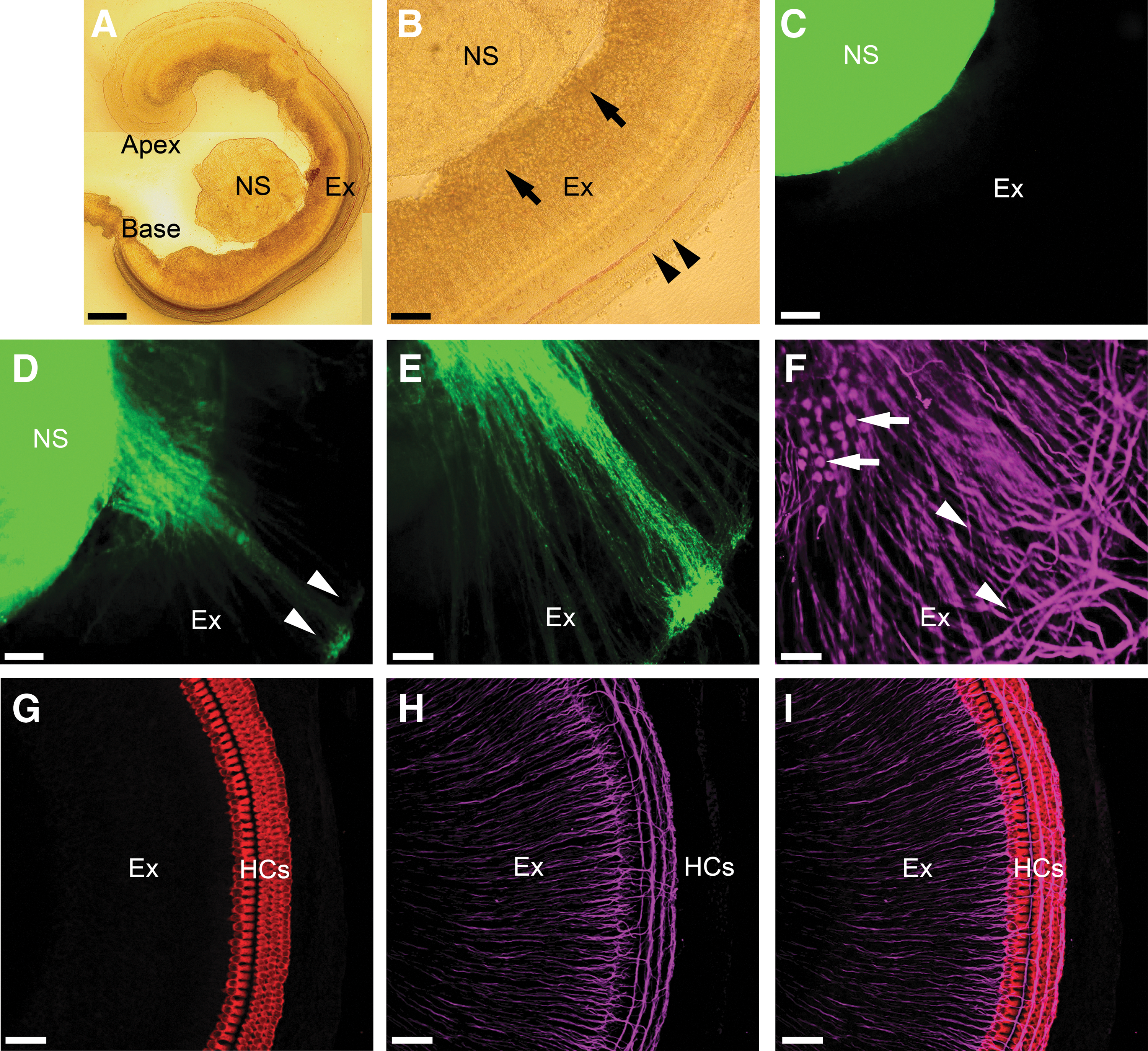
Innervation of cochlear explant cultures by treated NS. Treated NS [NS;
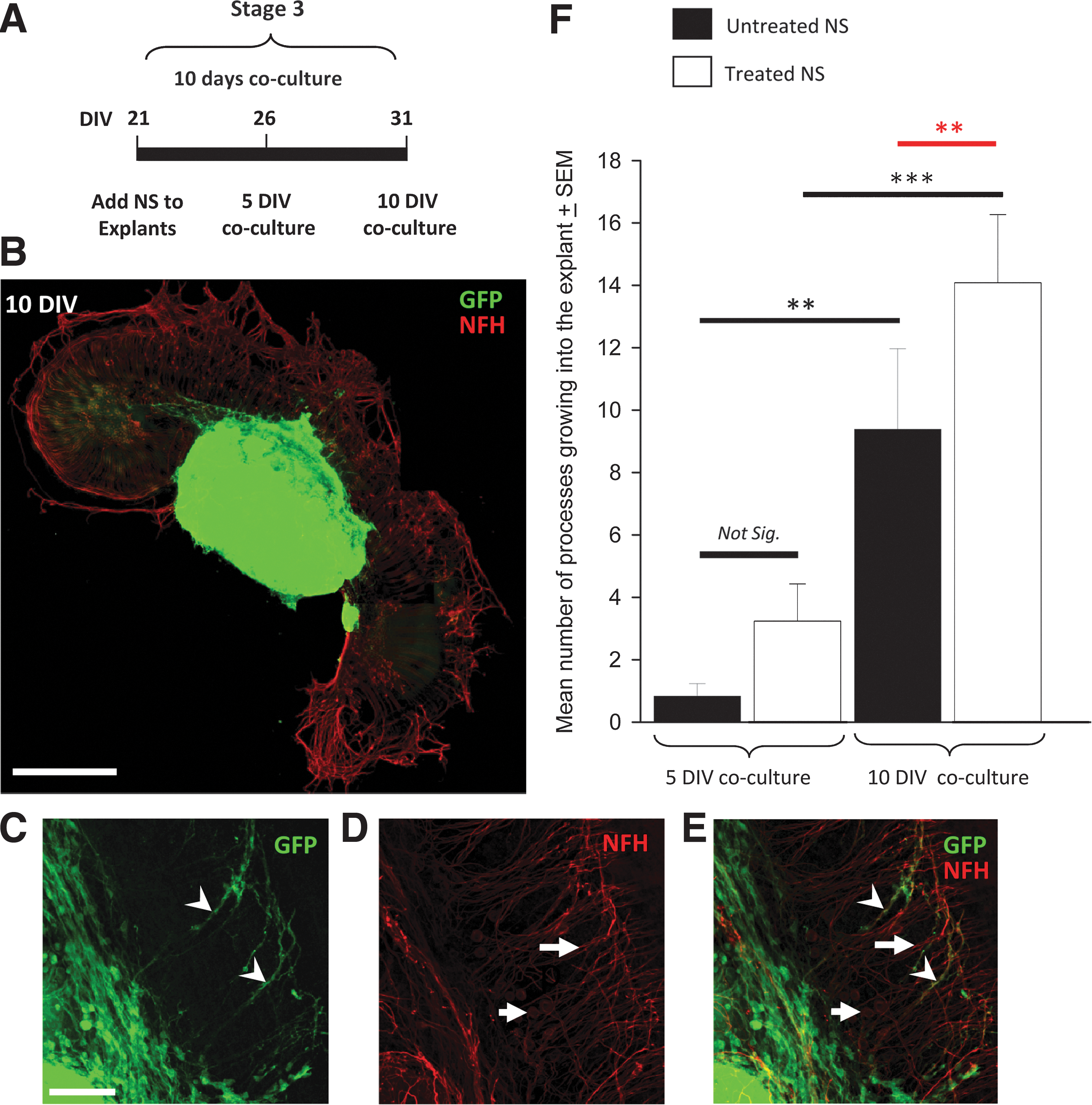
Treated NS innervate cochlear explants with greater efficacy than untreated NS. Cocultures of treated and untreated NS were examined after 5 and 10 DIV
Treated NS were observed to grow into the cochlear explant with greater efficacy than untreated NS
Cocultures of treated NS and untreated NS were subsequently set-up and cultured for either 5 or 10 DIV (Stage 3, Fig. 1), to examine whether there was any difference in the amount of growth into P3/4 cochlear explants at each time point (Fig. 5). An intact strip of sensory epithelium was required to create an in vitro system that mimicked the in vivo microenvironment as closely as possible. Processes from both treated and untreated NS were observed to preferentially grow toward and into the explant (Fig. 5B), and not away from the explant (P<0.001; all treatments and time points examined). Many GFP-positive hESC-derived processes were also observed to be NFH-positive (arrowheads, Fig. 5E) illustrating that a proportion of the stem cell-derived neural progenitors had reached neuronal maturity in the cocultures. Endogenous auditory neuron somata and their processes were observed via NFH labeling alone (arrows, Fig. 5D). In addition, we found that in both treated (P=0.003; black line **, Fig. 5F) and untreated (P<0.001; gray line ***, Fig. 5F) NS, there was a significant increase in the number of processes extending into the explant between 5 and 10 DIV (Fig. 5F). Importantly, there were significantly more processes from the treated NS growing into the explant (in comparison to processes derived from untreated NS), at 10 DIV (P<0.002, red line, Fig. 5F), but not at 5 DIV (P=0.1, NS, Fig. 5F).
To investigate whether the number of surviving hair cells was an influential factor in these experiments, we counted the number of hair cells present in cochlear explants in the following preparations: 1 day in culture (1 DIV), 10 days in culture (10 DIV), and 10 days in coculture with NS (10 DIV). Although slightly fewer hair cells were detected after 10 days both in culture and coculture preparations, there was no significant difference in the overall numbers of hair cells counted between any of the treatments (P=0.537; Supplementary Fig. S1; Supplementary Data are available online at
Processes derived from treated NS grow along hair cell rows in denervated cochlear explants
A denervation assay was employed next to determine whether treated NS could grow toward the sensory hair cells in the early postnatal cochlea, after removal of the endogenous peripheral processes and neurons of the explant (Fig. 6A–D). All denervated explant cocultures were examined after 10 DIV to allow for sufficient outgrowth of processes, and immunochemistry was used to specifically label the hair cells (Myosin VIIa-positive) and any stem cell-derived processes (GFP-positive). We found that stem cell-derived processes grew toward the hair cells in the denervated cochlear explant cocultures, and along the hair cell rows (Fig. 6E–H). Promisingly, we observed growth of hESC-derived neuronal processes toward isolated hair cells in the culture (arrow, Fig. 6G), seemingly illustrating directed growth toward this cell type. These data illustrated that stem cell-derived processes from treated NS were capable of locating the sensory hair cells in the absence of endogenous auditory neurons and their peripheral processes. Note the fine and punctuate morphological appearance of the hESC-derived processes (Fig. 6H), similar to immature, developing neural processes in situ.
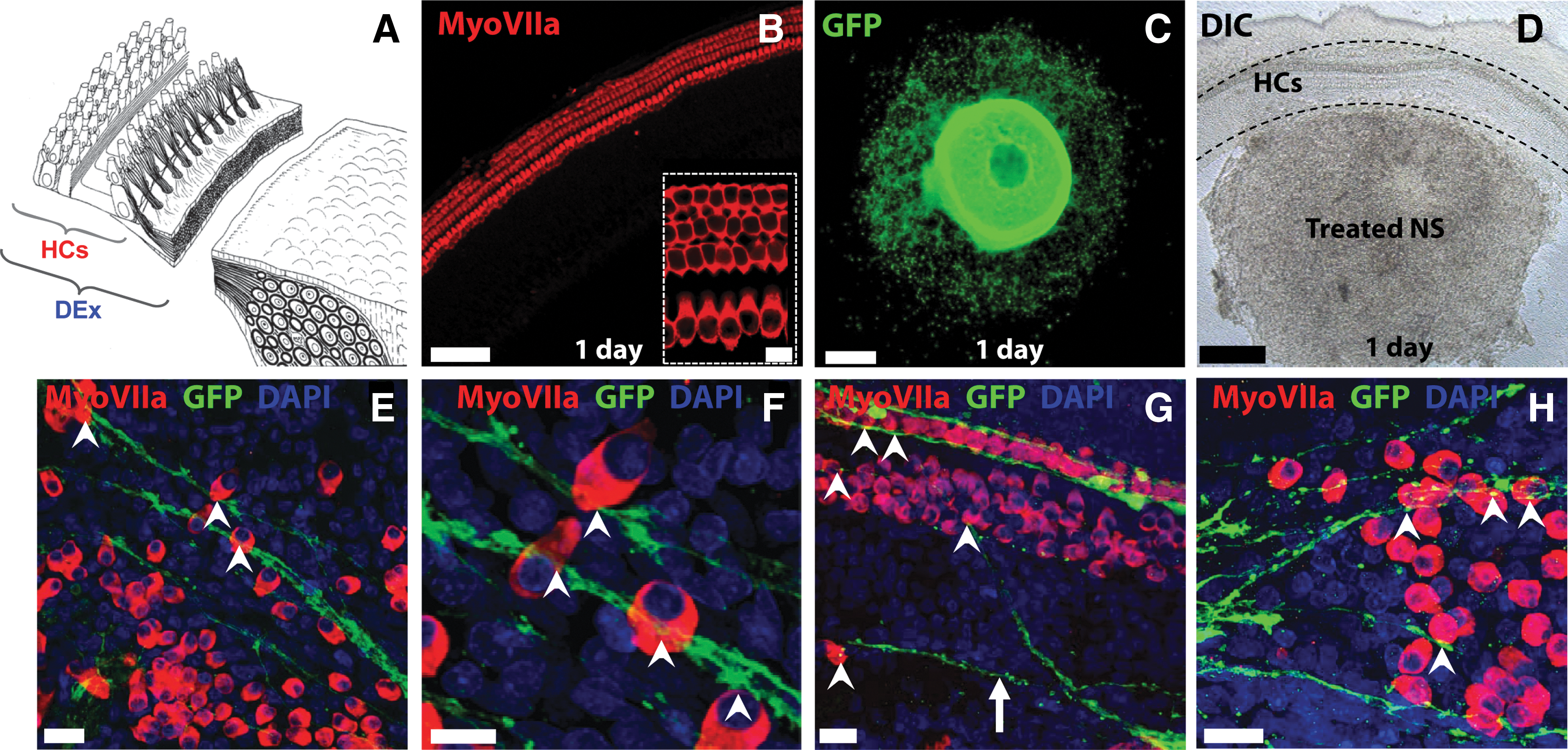
Processes from treated NS innervate denervated cochlear explants. Denervated cochlear explant cultures (DEx) were prepared by dissecting the HCs from the central modulus, as illustrated in
Stem cell-derived processes make en passant presynaptic contacts with sensory hair cells in vitro
When treated NS were cocultured with denervated cochlear explants (containing the sensory hair cells of the cochlea, but not the endogenous neurons or processes), extensive presynaptic (synapsin 1-positive) terminals were observed in the stem cell-derived processes (Fig. 7A–L) and between stem cell-derived processes and hair cells (Fig. 7M–O). Synapsin 1-positive contacts on the hair cells were observed to occur in an en passant manner, that is, a single stem cell-derived neurite made multiple synaptic connections as it extended along the row of hair cells (Fig. 7B, C, H, I, M, O). These presynaptic terminals were observed within a week of the cocultures being established, and were still present after 2 weeks. Synapsin 1 labeling alone is shown in Fig. 7D–F, J–L (corresponding to Fig. 7A–C, G–I, respectively). High-magnification images of the synapsin 1-positive points of contact between stem cell-derived neurites and hair cells are shown in Fig. 7O (arrows). Alternatively, when we examined the same preparations for the presence of postsynaptic terminals, only occasionally were these observed (data not shown). We also noted that distal NF-positive endogenous fibers disappeared after transection from the organ of Corti (Supplementary Fig. 2). Taken together, these data highlight the potential for treated NS derived from hESCs to both grow into and make many synaptic contacts on appropriate sensory tissues in the developing peripheral auditory nervous system.
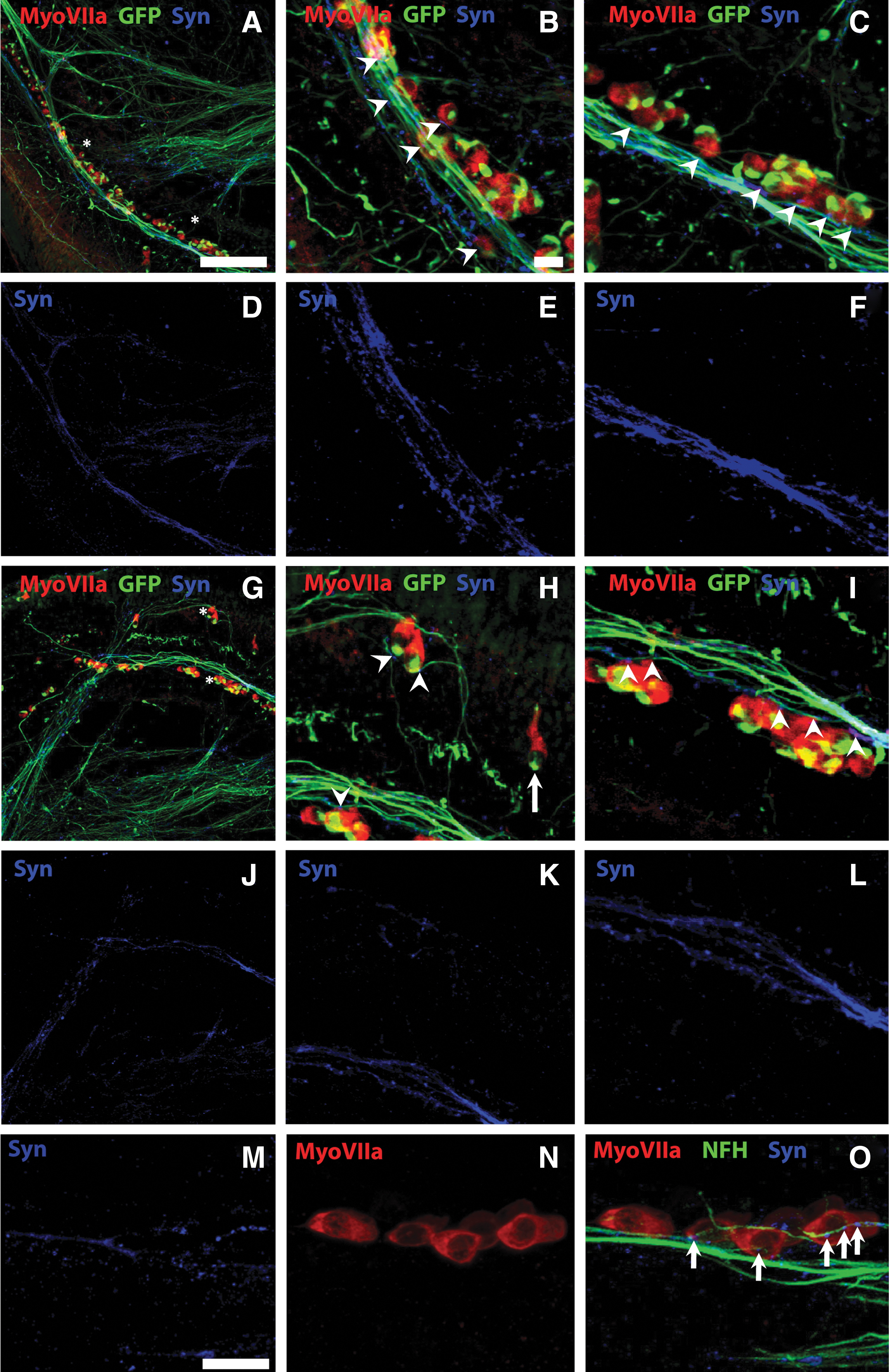
NS-derived processes make en passant presynaptic terminals on sensory HCs in vitro. Treated NS were observed to innervate and make abundant presynaptic contacts (Synapsin 1; blue) when grown in coculture with denervated cochlear explants for 10 DIV. Low-magnification images
Discussion
Stem cell therapy is one way by which auditory neurons could be replaced in the deaf mammalian cochlea, but to be effective, stem cell-derived neurons must be capable of differentiation into an appropriate phenotype, be electrically active, and be capable of innervating appropriate endogenous tissues [14]. Eventually, they will also need to be derived from human sources. In the current study, we demonstrate that hESC-derived neurons can be differentiated to a neurosensory lineage that is electrically active, can innervate the developing cochlear epithelium, and can make synapses on appropriate tissues. While both the treated and untreated NS were observed to express neurosensory proteins at the time points examined, only the treated NS were observed to extend significantly more processes into the explant after 10 DIV.
Possible explanations for these observations may be that the NS treated with Y27632 are better able to extend their processes further into the explant. This hypothesis is supported by previous studies reporting increased proportion of migrating neural crest-like progenitors from MEF/Y27632 treatment of hESC-derived NS [33]. Alternatively, or perhaps in addition, treated NS may contain more auditory neural progenitor-like cells. Although we did not detect a significant difference in the percentage of cells expressing Pax2 or Brn3a between treated and untreated NS (data not included), there were clearly more cells located on the peripheral edge of the treated NS (Fig. 2A, B). The observation that combined Y27632/MEF treatment causes cells to migrate out of the NS toward the periphery is consistent with our previous studies using this differentiation assay [33]. It may be that treatment with MEFs cocultures and/or Y27632, specifically enhances the extension of hESC-derived neurites in general. Further detailed studies are needed, which are beyond the scope of this article, to determine the mechanism of the enhanced outgrowth observed in the treated NS cocultures. Regardless, these findings have important implications for both hair cell regeneration and auditory neuron replacement studies, as transplanted stem cell-derived progenitors ideally need to extend peripheral processes toward the hair cells in the organ of Corti, and/or toward second order neurons in the cochlear nucleus [13]. Priming neural progenitors with Y27632 and MEFs during the differentiation process improves their ability to innervate target tissue, thereby improving the probability that these cells may synapse on the correct tissues following their transplantation in vivo.
When designing the experiments described herein, we hypothesized that the endogenous peripheral processes of the explant might provide a tissue scaffold for stem cell-derived processes to grow along, essentially providing a guide to the hair cells located on the periphery of the explant. In early experimentation, both treated NS (Fig. 4D, E) and untreated NS (Fig. 5B–D) were observed to extend their processes in close proximity to the endogenous peripheral processes of the explant. This was encouraging, given there is usually a complement of auditory neurons that remain after hearing loss, and transplanted stem cells may be able to use these to guide their correct connectivity in vivo. More promising, however, was the observation that these stem cell-derived processes were consistently capable of locating and growing along the hair cell rows, including making multiple synaptic contacts, after the removal of endogenous auditory neurons and their processes. This observation suggests some innate properties of both the neural differentiated hESC processes and an innate signaling mechanism from developing hair cells.
Presynaptic staining is not expected for afferent dendrites; however, we (and others) have observed presynaptic puncta between the peripheral processes of auditory neurons and hair cells in early postnatal cochlear explant cultures (Supplementary Fig. S3, [8]) and adult cochleae [8]. In addition, early postnatal afferent auditory neurons express presynaptic markers (synaptophysin and synapsin), where they come into contact with hair cells, following their dissection from the sensory epithelium [39,41,42]. Presynaptic terminals have also been reported between stem cells and hair cells using a similar in vitro model [8,11]. These observations, therefore, support the idea that stem cell-derived neurons behave in a similar manner to developing auditory neurons, and that putative presynaptic specializations within the neural processes may be attributable, in part, to the limited competition between neural terminals in these cultures [39]. Future experimentation using genetically modified animals [43,44], may start to dissect this process more thoroughly and determine the role of each (hair cells and neurons) in pathfinding and synaptogenesis in the developing cochlea. The more general observation that human stem cell-derived neurites can grow toward and synapse upon hair cells is exciting for future in vitro assays, and provides a model for further exploring the mechanisms involved in cochlear synaptogenesis. Future investigations using paired electrophysiological recordings between stem cell-derived neurons and hair cells, will provide useful information on the characterization and function of these new synapses.
The potential of stem cells to effectively replace auditory neurons lies in their ability to correctly relay auditory information with precise timing. Such competence is only possible if stem cell-derived neurons are capable of firing an action potential in response to depolarization. We observed that hESC-derived neurons were capable of firing action potentials, and their firing profile was consistent between all cells examined. This suggests that the described culture conditions generate a population of cells with similar electrical properties and functional potential. In addition, hESC-derived neurons displayed the phasic profile of activity that has been previously reported for the majority of auditory neurons in vitro [45 –48], and this profile was maintained for up to 49 DIV. While our future investigations are examining and characterizing the involvement of specific ion channels in this firing response, the observations described herein illustrate that hESC can be differentiated into functional neurons with firing properties similar to those reported from auditory neurons grown in vitro [49].
While hESC neural derivatives have been reported to make synaptic connections with hair cells in vitro [11], this has typically occurred in low frequency. Conversely, the present study highlights the extensive formation of presynaptic terminals between hESC-derived neurons and hair cells in vitro. The en passant-like nature of the new presynaptic terminals between the stem cell-derived neural processes and the sensory hair cells is intriguing, because of their similarity to the pattern of type II auditory neuron innervation reported in the normal hearing cochlea [50 –52]. Rather than a single synaptic bouton, which is typical of the type I-inner hair cell synapse, this en passant-like contact may ultimately result in a different physiological function. It is also possible that hESC-derived processes grow in an en passant-like manner during development, but mature into boutons after the onset of spontaneous activity in the cochlea [53], at which time, synaptic input is thought to be refined. This observation is further supported by the observation that hESC-derived processes were negative for the efferent marker acetylcholine, and that extensive presynaptic labeling in developing cochlear explant cultures was also observed (Supplementary Fig. S3). Previous studies have illustrated that the peripheral innervation of the cochlea is initially complex and branched, but matures into more simplified and straight processes during development [54 –56]. The fine, punctate morphological appearance of the hESC-derived processes supports their immaturity, considering the morphological appearance of developing neurites in situ.
Summary
In conclusion, the present study illustrates that treated NS (cultured on MEFs in the presence of Y27632) produce a mixed population of neurosensory progenitors, which are electrically active, can extend neurites into early postnatal cochlear tissues, and form extensive presynaptic contacts with sensory hair cells in vitro. These findings are promising for emerging hair cell regeneration studies, which require a healthy complement of auditory neurons, and our combined stem cell/cochlear implantation experiments, which are aimed at investigating the role of electrical stimulation in improving stem cell function and connectivity in vivo. The ability to form synaptic terminals is the first step toward improving neural function using stem cells in the treatment of neurodegenerative disease.
Footnotes
Acknowledgments
The authors would like to thank Dr. Aurore Brugeaud for her assistance teaching the early postnatal cochlear denervation dissection, and Ms. Ricki Minter and Ms Rachael Sheridan for their technical laboratory support. We also thank Professors David Ryugo, Ed Rubel, Rémy Pujol, and Peter Thorne for helpful discussions regarding the interpretation of experimental data and Dr. Clare Parish for her collaboration and provision of reagents. We gratefully acknowledge financial support from the following organizations: the National Health and Medical Research Council of Australia, the Garnett Passe and Rodney Williams Memorial Foundation, the Friedreich Ataxia Research Association, the University of Melbourne: Departments of Otolaryngology, Pharmacology and the Centre for Neuroscience, and the Royal Victorian Eye and Ear Hospital.
Author Disclosure Statement
The authors disclose no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.