
Abstract
Cellular transplantation strategies for repairing the injured spinal cord have shown consistent benefit in preclinical models, and human clinical trials have begun. Interactions between transplanted cells and host tissue remain poorly understood. Trophic factor secretion is postulated a primary or supplementary mechanism of action for many transplanted cells, however, there is little direct evidence to support trophin production by transplanted cells in situ. In the present study, trophic factor expression was characterized in uninjured, injured-untreated, injured-treated with transplanted cells, and corresponding control tissue from the adult rat spinal cord. Candidate trophic factors were identified in a literature search, and primers were designed for these genes. We examined in vivo trophin expression in 3 paradigms involving transplantation of either brain or spinal cord-derived neural precursor cells (NPCs) or bone marrow stromal cells (BMSCs). Injury without further treatment led to a significant elevation of nerve growth factor (NGF), leukemia inhibitory factor (LIF), insulin-like growth factor-1 (IGF-1), and transforming growth factor-β1 (TGF-β1), and lower expression of vascular endothelial growth factor isoform A (VEGF-A) and platelet-derived growth factor-A (PDGF-A). Transplantation of NPCs led to modest changes in trophin expression, and the co-administration of intrathecal trophins resulted in significant elevation of the neurotrophins, glial-derived neurotrophic factor (GDNF), LIF, and basic fibroblast growth factor (bFGF). BMSCs transplantation upregulated NGF, LIF, and IGF-1. NPCs isolated after transplantation into the injured spinal cord expressed the neurotrophins, ciliary neurotrophic factor (CNTF), epidermal growth factor (EGF), and bFGF at higher levels than host cord. These data show that trophin expression in the spinal cord is influenced by injury and cell transplantation, particularly when combined with intrathecal trophin infusion. Trophins may contribute to the benefits associated with cell-based repair strategies for spinal cord injury.
Introduction
C
Trophic factor production has been postulated a primary or supplementary mechanism of action for transplanted cells, in particular where neuroprotective effects have been described and where benefit persists despite death of the transplanted cells [5,7 –9]. Although alternate mechanisms such as environmental detoxification or provision of metabolic support could also explain these findings [10], neurotrophins and related trophic factors could underlie these beneficial effects, as they have been ascribed wide-ranging, repair-promoting effects within the CNS [5,11 –13]. Trophic factors recruit and stimulate proliferation and differentiation of neural precursor cells (NPCs) [14]. They also have antiapoptotic effects [15] and enhance axonal regrowth [16], remyelination [17,18], and neuronal plasticity [19,20]. Since such effects have been noted after transplantation of NPCs and bone marrow stromal cells (BMSCs) [21 –24], it is possible that both cell types could provide trophic support either by secreting trophins or by inducing trophin production by host cells. Currently there is little direct evidence to support this. Recently, we have found differences in trophic factor expression in NPCs and BMSCs in vitro in various culture conditions [25]. The purpose of the present study was to characterize the in vivo expression profile of trophic factors in the uninjured and injured rat spinal cord and after NPC and BMSC transplantation into the injured cord.
To determine whether trophins could contribute to recovery after SCI, we first characterized trophin production in the normal and injured spinal cord. We then examined trophin production after transplantation of either brain or spinal cord-derived NPCs or BMSCs into the injured rat spinal cord. These protocols are similar to those employed in trials involving human SCI patients [26 –28]. In addition, we used fluorescence-activated cell sorting (FACS) to isolate exogenous cells after transplantation in one of these models, facilitating the characterization of a pure isolate of these cells. This is the first study that characterizes trophin expression in vivo after NPC and BMSC transplantation into the injured rat spinal cord, and in a pure population of exogenous cells isolated post-transplant. These results enhance our understanding of trophic factor expression after SCI and cell transplantation, which is important for the development of therapeutic strategies for SCI.
Materials and Methods
General animal care and surgical procedures
All animal work was conducted in accordance with the Canadian Council on Animal Care guidelines, and local institutional ethics approval was obtained for the experiments performed. Prophylactic preoperative amoxicillin-clavulinic acid was provided as infection prophylaxis. Animal surgeries were performed using sterile technique in conjunction with halothane or isofluorane anesthesia. Buprenorphine was administered postoperatively for analgesia. Animals were given food and water ad libitum, and their bladders were manually expressed thrice a day. Blinding to group assignment in all 3 transplant paradigms was maintained until statistical analysis was completed. Sample sizes employed are presented in Supplementary Table S1; Supplementary Data are available online at
Isolation of cellular substrates
Spinal cord NPCs employed in transplant Paradigm 1 were isolated from the spinal cords of transgenic adult female Wistar rats expressing enhanced green fluorescent protein (eGFP) (Wistar-TgN(CAG-GFP)184ys) (YS Institute Inc.) [29]. The isolation of NPCs derived from the adult periventricular spinal cord was performed as previously described [30]. Cervical and thoracic spinal cord was excised under sterile conditions and washed in Dulbecco's phosphate-buffered saline supplemented with 30% glucose (Sigma-Aldrich) and 1% penicillin/streptomycin (Sigma-Aldrich). The overlying meninges, blood vessels, and white matter were removed. Tissue from the periventricular region was then harvested. Dissected spinal cord tissue was enzymatically dissociated in a solution containing 0.01% papain and 0.01% DNase I for 1 h at 37°C, and then mechanically dissociated into a cell suspension that was centrifuged using a discontinuous density gradient to remove cell membrane fragments. Cells were resuspended in Neurobasal-A medium (Gibco-Invitrogen) supplemented with B27 (Gibco-Invitrogen), L-glutamine (Gibco-Invitrogen), penicillin/streptomycin (Gibco-Invitrogen), 20 ng/mL each of epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF) (Sigma-Aldrich), 2 μg/mL heparin (Sigma-Aldrich), and hormone mix as described [30]. Cells were plated in uncoated Nunc T25 culture flasks (VWR International), and the neurospheres generated were passaged weekly by mechanical dissociation in the serum-free medium just described.
To generate NPCs from adult mouse brains employed in transplant Paradigm 2, enhanced yellow fluorescent protein (eYFP)-bearing mice were killed by cervical dissociation, and brains were removed using the sterile technique. The subventricular zone of the forebrain was dissected as previously described [31]. To dissociate cells, isolated tissue was transferred to a 30 mL low calcium artificial cerebrospinal fluid enzyme solution containing 40 mg of trypsin (Sigma), 20 mg of hyaluronidase (Sigma), and 3.5 mg of kynurenic acid (Sigma) for 1 h at 37°C. Trypsin was then inactivated, and cells were mechanically dissociated by agitation with a Pasteur pipette. Cells were then transferred to uncoated T25 tissue culture flasks in 10 mL of serum-free medium containing 1 mL of DMEM/F-12, 0.2 mL of 30% glucose, 0.15 mL of 7.5% NaHCO3, 0.05 mL of 1 M HEPES, 0.05 μg of bFGF, 0.1 μg of EGF, 1% L-glutamine (Gibco-Invitrogen), and 1% penicillin/streptomycin (Sigma) for 7 days. The neurospheres generated were passaged weekly by mechanical dissociation in the same medium.
BMSCs employed in transplant Paradigm 3 were cultured as previously described [32,33]. Briefly, bone marrow was collected from the femurs, and the tibia of the eGFP transgenic rats just described (∼40–50×107 cells per rat) and resuspended in long-term bone marrow culture medium. The cells were incubated at 37°C in 5% CO2. Cells were passaged after 2 weeks, and then every 5–7 days. Cells were characterized according to the methods recommended by Dominici et al. [34]. Using this protocol, we have previously shown that BMSCs show no evidence of differentiation in vitro into astrocytes, oligodendrocytes, or neurons [32,35].
Transplantation paradigms
Paradigm 1—adult spinal cord NPC transplantation
Adult female Wistar rats were subject to a 1 min 26g clip compression injury [36] at the T8 bony level following a laminectomy from T8-9. One week later, 4 animals were randomized to transplantation with 2×105 P3 or P4 adult spinal cord-derived NPCs, and 3 animals in the control group were injected with culture medium alone. Rats were anesthetized with inhalation of halothane, and the injury site was re-exposed. Five microliters of cells or media was injected at 2 intraspinal sites at the midline, ∼1 mm rostral, and caudal to the injury epicentre. Both groups were treated with daily doses of cyclosporin (10 mg/kg; Novartis, administered subcutaneously). Animals from this paradigm were sacrificed one week after experimental or control transplant. 1 cm of perilesional cord centered on the epicenter was then extracted from each animal for RNA isolation.
Paradigm 2—adult brain NPC xenotransplantation with trophin infusion
Adult female Wistar rats were subject to a 1 min 23g clip compression injury [36] at the T7 bony level after a laminectomy from T6-8. Two weeks later, the animals were randomly assigned to one of 5 experimental groups, 3 of which were previously investigated by our group [31]. The Injured No Treatment group received no further treatment. For the other experimental groups, ‘IS’ denotes a 10 days course of minocycline (50 mg/kg; Sigma, administered intraperitoneally) and daily cyclosporin (10 mg/kg; Novartis, administered subcutaneously) both starting 2 days before control or cell transplantation. ‘GF’ denotes implantion of an osmotic mini-pump (Alzet model 1007D, primed overnight) delivering EGF (3 μg/100 μL; Sigma), bFGF (3 μg/100 μL; Sigma), and platelet-derived growth factor-A (PDGF-A) (1 μg/100 μL; Sigma) for 7 days at the time of cell or control transplantation. Distinct from our group's previous work, the catheter tip was placed 1 cm rostral to the epicenter to prevent dislodgement. Cellular and control transplantation was conducted at 4 intraspinal sites, ∼2 mm rostral, and caudal to the injury epicenter on both sides of the midline (2 μL each). Injured + GF/IS animals underwent sham transplantation with cell suspension media alone. Animals transplanted with NPCs received a total of 4×105 live cells. The Dead Cell + GF/IS group was transplanted with the same number of NPCs killed by 10 freeze-thaw cycles [37]. In this paradigm, at least 3 animals per group were employed in quantitative polymerase chain reaction (qPCR) and protein studies.
Animals from this paradigm were sacrificed 1 week after experimental or control transplant. One centimeter of perilesional spinal cord was then extracted from each animal for RNA and protein isolation. These segments were centered on the epicenter and spanned both rostral and caudal transplant sites. 1 cm of uninjured spinal cord tissue was also extracted from equivalent regions of uninjured rat spinal cord for both RNA and protein analyses to facilitate comparison.
Paradigm 3—adult BMSC transplantation via lumbar puncture and infusion
Adult female Wistar rats were subject to a 1 min 26g clip compression injury [36] at the T8 bony level after a laminectomy from T8-9. One week later, animals were randomized to transplantation with 2×106 adult rat-derived BMSCs administered via lumbar puncture and the control group that was infused with vehicle [0.1M phosphate-buffered saline (PBS) and 0.2% bovine serum albumin (BSA)]. Rats were re-anesthetized as just described and placed prone. A longitudinal incision was made over the L3-5 spinous processes, and the skin was retracted as previously described [35]. A neonatal 25 gauge LP needle was advanced into the spinal canal at L3-4 or L4-5. Proper placement of the needle was determined by the presence of CSF in the hub of the needle. The CSF present in the needle hub was aspirated using a micropipette and 40 μL containing 2×106 BMSC in 0.1M PBS and 0.2% BSA or vehicle alone was slowly injected into the intrathecal space [35]. Both groups were treated with daily doses of cyclosporin (10 mg/kg; Novartis, administered subcutaneously). Animals from this paradigm were sacrificed one week after experimental or control transplant. 1 cm of perilesional cord centered on the epicenter was then extracted from each animal for RNA isolation. To characterize delivery of transplanted cells to the uninjured spinal cord as well as a potential trophin influence in uninjured regions, control tissue was also extracted from all animals from a 1 cm region just above the conus medullaris and subject to the same analysis.
Fluorescence-activated cell sorting
1 cm spinal cord segments centered on the injury epicenter from NPC + GF/IS animals (Paradigm 2) were extracted and then incubated in 0.1% collagenase for 1 h at 37°C. After trituration, digested tissue was re-suspended in medium containing 1×HBSS (Ca and Mg free), 10 mM HEPES pH 7.2, 2% fetal bovine serum, and 10 mM NaN3 and passed through a 70 μm cell strainer. Cells were then re-suspended at a concentration of 5–10 million cells/mL, and incubated with propidium iodide (10 μg/mL) for 30 min at room temperature to facilitate exclusion of dead cells. A Becton Dickinson FACS Aria 13-colour cell sorter was employed to isolate eYFP+ and eYFP- cells after calibration with positive and negative controls. Samples from 3 animals were pooled and diluted to the minimum volume required to perform qPCR in triplicate with all primer pairs.
RNA isolation and cDNA synthesis
For RNA isolation, RNaseZAP® (Gibco-Invitrogen) was used to treat all tools, gloves, and working areas employed. Cultured cells were collected and homogenized in Trizol® (Gibco-Invitrogen). After extraction with chloroform, removal of the aqueous layer, precipitation with isopropyl alcohol, and a wash with 75% ethanol, the RNA pellets were resuspended in RNase free water. Spectroscopy was then performed using a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific) to ensure successful RNA harvest and determine its concentration. cDNA was subsequently synthesized using Superscript II reverse transcriptase (Gibco-Invitrogen) and Oligo(dT)12–18 primers (Gibco-Invitrogen).
qPCR analysis
Candidate trophic factors were identified in a literature review. Primers were designed for factors consistently found to be produced by neural cells, upregulated at the epicenter of a CNS injury, or of demonstrated benefit in CNS repair [12,38
–46]. Primers were designed for nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT-3), neurotrophin-4/5 (NT-4/5), glial-derived neurotrophic factor (GDNF), ciliary neurotrophic factor (CNTF), PDGF-A, EGF, bFGF, leukemia inhibitory factor (LIF), insulin-like growth factor-1 (IGF-1), glial growth factor 2 (GGF2), transforming growth factor-β1 (TGF-β1), and vascular endothelial growth factor isoform A (VEGF-A). Primers were also designed for the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH), which has been employed in trophin studies by several other groups [47
–49]. Primers were designed with careful consideration of the structure of each gene ensuring the sequence was common to all active isoforms or unique to specific isoforms where indicated. The most highly conserved sequence in these regions was entered into PrimerQuest software (
With the exception of specimens obtained from FACS sorting, cDNA template was diluted to 3 ng/μL. qPCR reactions were performed using SYBR Green chemistry and an ABI 7900HT fast real-time PCR thermal cycler (Applied Biosystems). Absolute quantitation was performed, and all assays were performed in at least triplicate. Analysis was performed using SDS 2.3 software.
Western blotting
In Paradigm 2, 1 cm of perilesional spinal cord tissue centered on the epicenter was extracted from distinct animals in Ringer's Lactate maintained at 4°C. Protein samples were prepared after determination of protein concentration with the Lowry Method. Samples were then run on 12% polyacrylamide gels and then transferred onto nitrocellulose overnight. Coomassie Blue staining was employed to verify the presence of protein. Confirmatory western blotting for VEGF-A was performed using an antibody specific for the ‘A’ isoform (Santa Cruz; sc-152, 1:200 dilution incubated at 4°C overnight). Confirmatory western blotting for BNDF was also performed (Santa Cruz, 1:500 dilution incubated at 4°C overnight). Normalization was performed to β-actin (Chemicon; MAB1501R, 1:400 dilution incubated at 4°C overnight). Anti-mouse-HRP conjugated secondary antibody was subsequently employed for 1h at room temperature (Sigma; A-3682, 1:2,000). Images were generated for densitometry measurements using a Fluor-S MultiImager (Bio-Rad), and measurements were made using Quantity One software (Bio-Rad, version 4.2.1). Alternate normalization to GAPDH (Sigma; G8795, 1:7,000) provided equivalent results.
Statistical analysis
SAS 9.2 software was used for statistical analysis. Results were analyzed with analysis of variance as a first step in all analyses and when significant at the 0.05 level, subsequent inter-group comparisons were made with a conservative Bonferroni adjustment for multiple comparisons.
Results
Trophin expression in the uninjured lower thoracic spinal cord
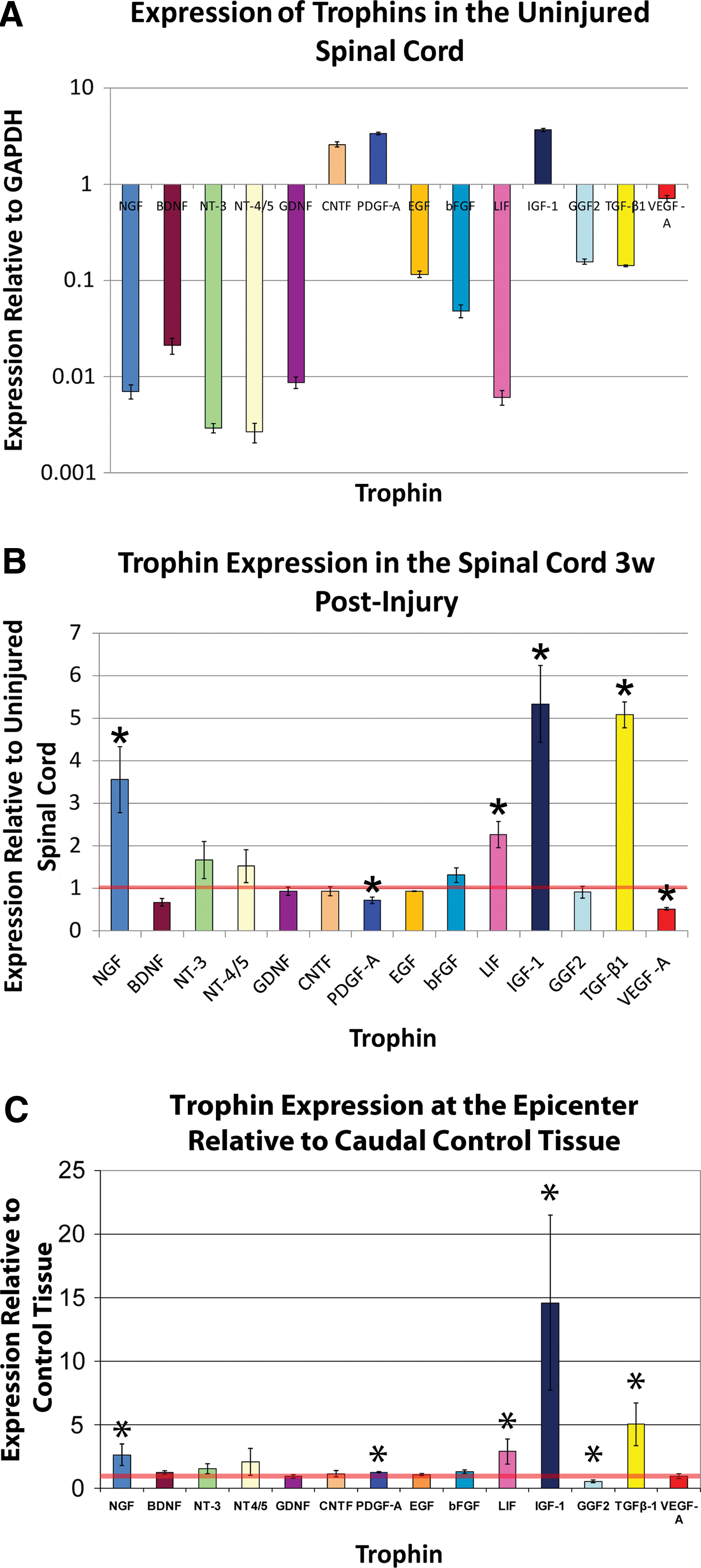
Figure 1A demonstrates that the uninjured lower thoracic spinal cord expresses detectable levels of all assayed trophins including NGF, BDNF, NT-3, NT-4/5, GDNF, CNTF, PDGF-A, EGF, bFGF, LIF, IGF-1, GGF2, TGF-β1, and VEGF-A. The expression level varied greatly among the various factors when compared with expression of the house-keeping gene GAPDH. The neurotrophins, GDNF, and LIF were expressed at the lowest levels. Only CNTF, PDGF-A, and IGF-1 were expressed at levels higher than GAPDH.
SCI influences trophin expression
In a comparison of trophin expression in equivalent segments of injured (Injured No Treatment animals from Paradigm 2) and uninjured lower thoracic spinal cord in distinct animals 3 weeks after SCI, qPCR found significantly elevated expression of NGF, LIF, IGF-1, and TGF-β1 (Fig. 1B). Expression of PDGF-A and VEGF-A was significantly reduced.
In control animals from Paradigm 3, a similar comparison was made. Here, injured regions of the spinal cord were compared with caudal uninjured regions of the same spinal cords 2 week postinjury in animals treated with cyclosporin. Figure 1C shows very similar results to the aforementioned comparison of injured and uninjured tissue at 3 week post-transplant. Unlike the previous comparison, however, here the injured tissue demonstrated significant upregulation of PDGF-A, significant downregulation of GGF2, and no change in VEGF-A expression.
NPC transplantation alone has modest effects on trophin expression
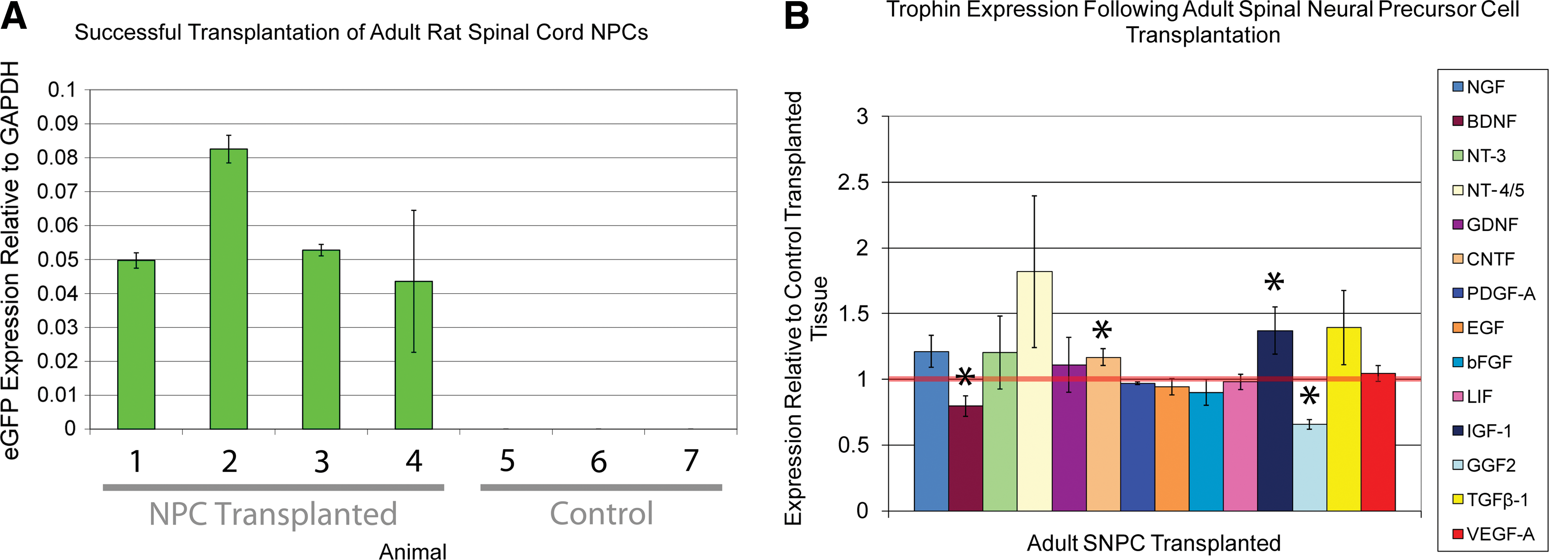
In Paradigm 1, a simple NPC transplantation protocol in which both NPC transplanted and control transplanted rats were treated with cyclosporin, trophin expression was compared in the region of the epicenter. In this model, all 4 animals transplanted with NPCs expressed detectable eGFP levels indicating successful transplantation (Fig. 2A). NPC transplantation was associated with a significant increase in CNTF and IGF-1 as well as significant decreases in BDNF and TGF-β1 (Fig. 2B). The magnitude of these differences is small and of uncertain biological significance.
NPC transplantation and trophin infusion is associated with a synergistic increase in trophin expression
Trophin expression in perilesional tissue was also assessed in a distinct combinatorial transplantation protocol (Paradigm 2). NPC transplantation combined with perilesional trophin infusion was associated with a significant increase in the expression of NGF, BDNF, NT-3, NT-4/5, GDNF, LIF, and bFGF (Fig. 3). Western blotting was performed for BDNF and VEGF-A, both of which demonstrated a close match between protein production and expression data (Fig. 4).
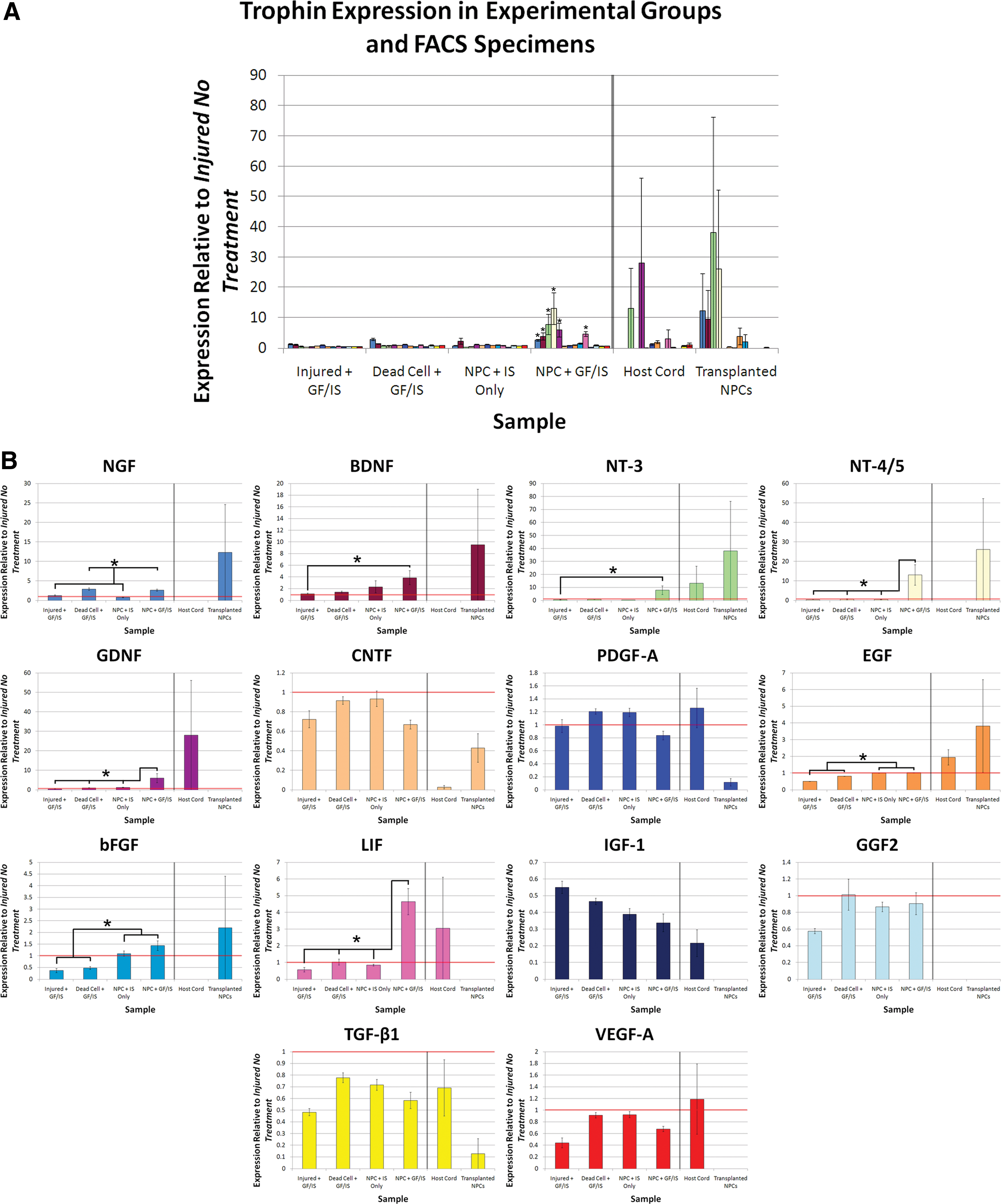
Trophin infusion and NPC transplantation is associated with a synergistic increase in trophin expression. (
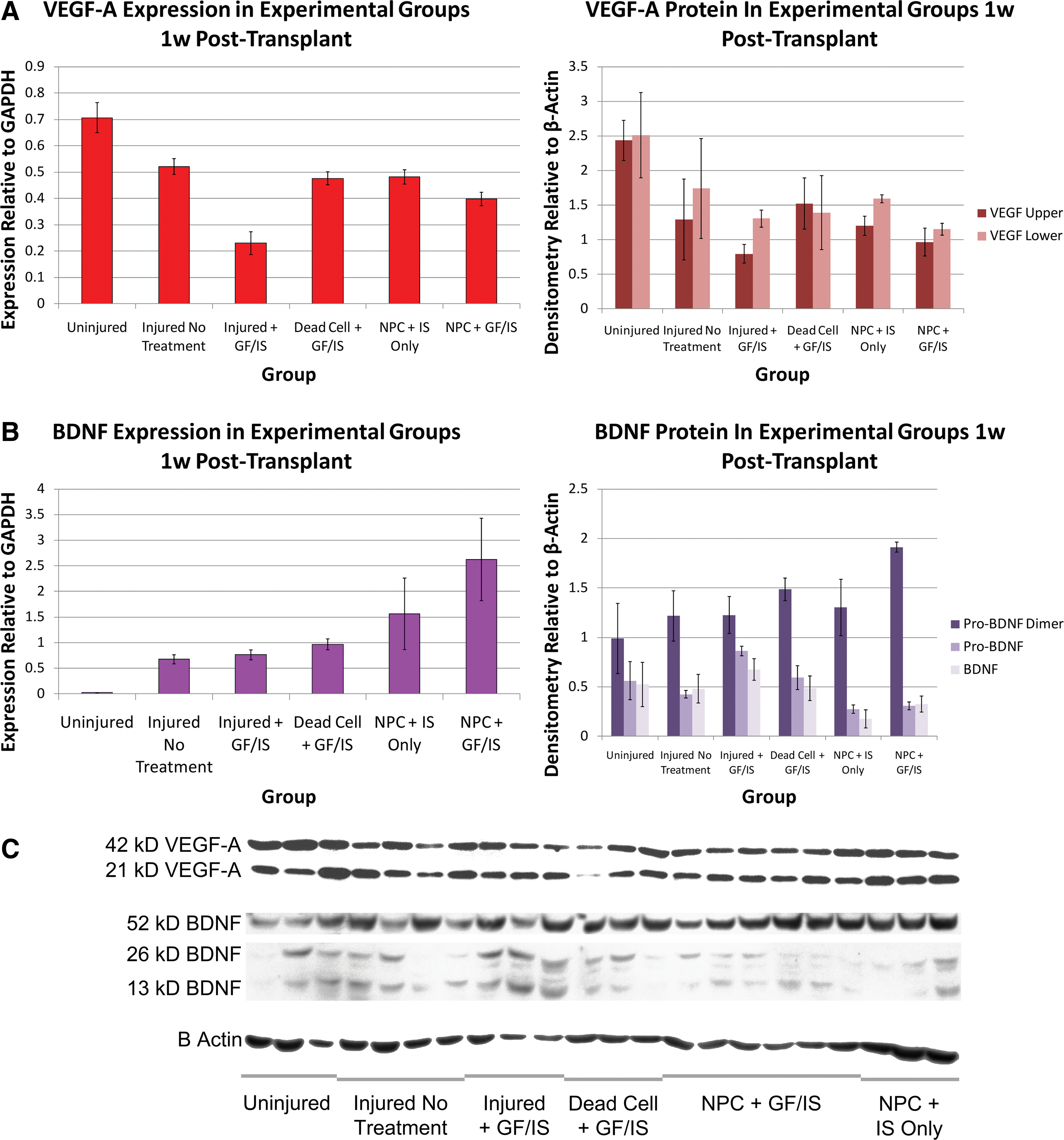
Trophin expression and protein quantitation for VEGF-A and BDNF are concordant. Concordant RNA (left) and protein (right) quantitation is shown for VEGF
The effect of NPC transplantation in combination with pharmacotherapy and trophin infusion is distinct from the administration of either alone. In this transplant paradigm, trophin infusion and pharmacotherapy alone was associated with a significant downregulation of EGF, bFGF, IGF-1, TGF-β1, and VEGF-A, while NPC transplantation and pharmacotherapy without trophin infusion was associated with significant downregulation of only IGF-1 and TGF-β1. A comparison of NPC + GF/IS and NPC + IS only groups suggests that the addition of a trophin pump is associated with a significant increase in NGF, NT-3, GDNF, and LIF with a significant decrease in PDGF.
In this paradigm, eYFP expression indicated successful transplantation of all 6 animals with NPCs (Supplementary Fig. S1). Trophin pump implantation did not increase the number of surviving NPCs. eYFP expression in the NPC + GF/IS group was 0.82% that of the pure eYFP+ FACS isolate and in the NPC + IS only group it was 2.22%. This suggests that the transplanted NPCs comprised <3% of all cells within the harvested region of the spinal cord.
NPCs express trophins after transplantation
FACS was employed to determine whether the aforementioned expression changes were attributable to host spinal cord or transplanted cells (Supplementary Fig. S1). In the 3 transplanted cords analyzed, the number of eYFP+ cells isolated ranged from 662 to 117,223 corresponding to 0.0037%–2.9% of cells in the harvested region, which is very consistent with the estimate calculated based on eYFP expression as just reported.
qPCR for trophin expression performed on eYFP+ and eYFP- FACS specimens found that the eYFP+ NPC-derived exogenous cells expressed detectable levels of all neurotrophins, CNTF, EGF, PDGF-A, and TGF-β1 and that the transplanted cells expressed higher levels of all 4 neurotrophins, CNTF, bFGF, and EGF than eYFP host cord cells. Although the result for CNTF approached statistical significance (P=0.0543), all statistical comparisons were not significant. Despite pooling the RNA obtained from 3 animals for this assay, trophin expression in these samples was near the limit of detection and subject to high variability necessitating careful interpretation.
BMSC transplantation upregulates several trophins
Of the 4 animals transplanted intrathecally with eGFP+ BMSCs via lumbar puncture, eGFP expression was detected in only 3 of the animals. The fourth was, thus, excluded from subsequent qPCR analysis (Supplementary Table S1). In all 3 successfully transplanted animals, higher eGFP expression was noted at the epicenter than at the caudal control region (Fig. 5). Intrathecal administration of BMSCs was associated with significant upregulation of NGF, LIF, and IGF-1 at the epicenter (Fig. 5) as well as a significant reduction in PDGF-A. BMSC transplantation did not lead to significant expression changes in the caudal control region, though BMSCs were detected there based on positive eGFP expression.
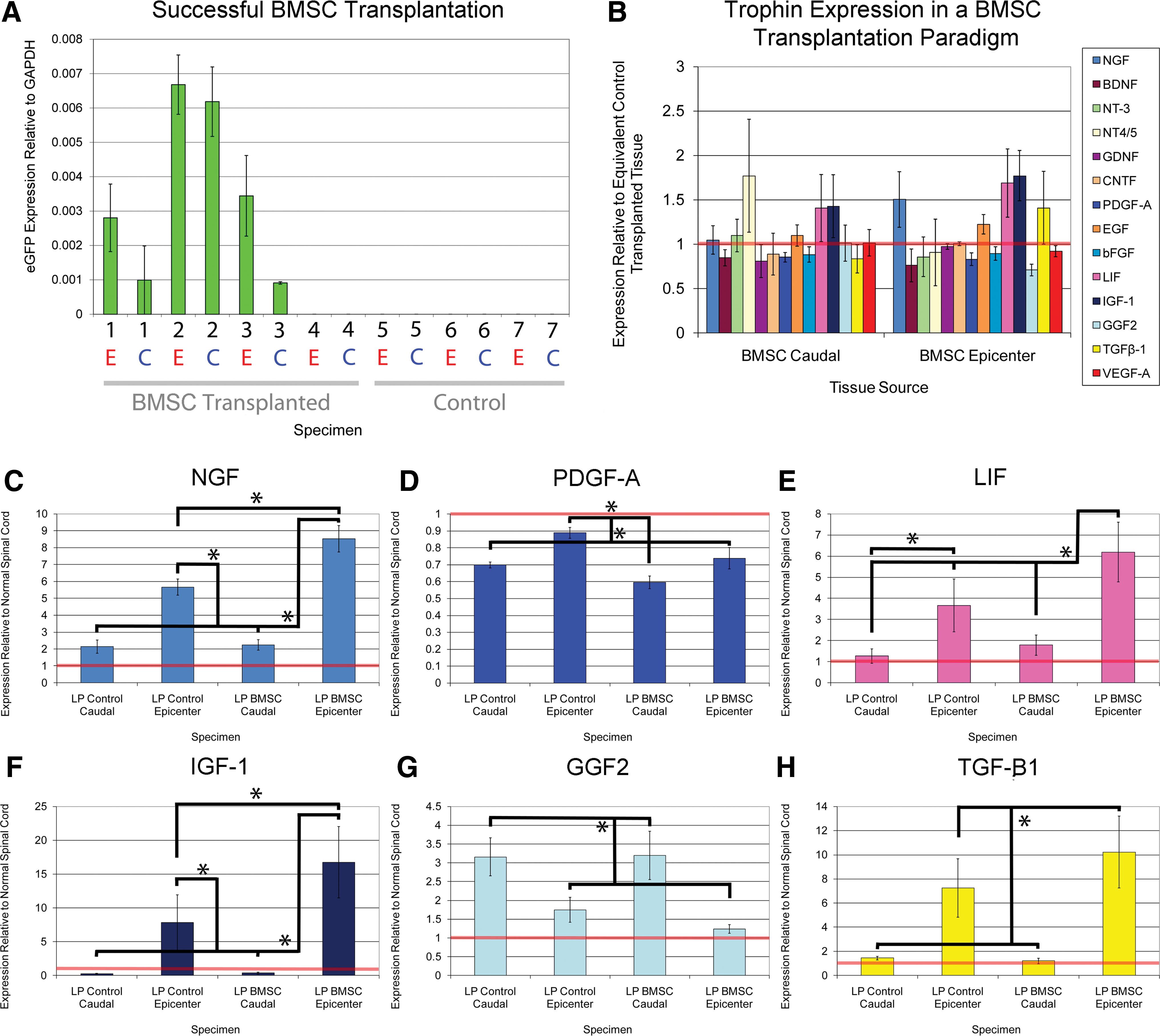
BMSC transplantation upregulates several trophins.
Cellular transplantation for SCI upregulates expression of inflammatory mediators
Tissue subject to qPCR from all 3 transplant paradigms was also analyzed with regard to expression of the inflammatory mediators IL-1β, IL-6, and TNF-α (Fig. 6). In all 3 cellular transplantation paradigms, transplanted cells expressed these inflammatory mediators at higher levels than control tissue. The increase in all 3 mediators reached significance on post-hoc testing for the NPC + GF/IS group in Paradigm 2. In this paradigm, NPC transplantation alone (NPC + IS only) was associated with lower levels of inflammatory mediator expression than an injured untreated SCI when NPCs were administered along with the immunosuppressant drugs minocycline and cyclosporin. In Paradigm 3, IL-1β was significantly increased in the region of the epicenter by BMSC transplantation.
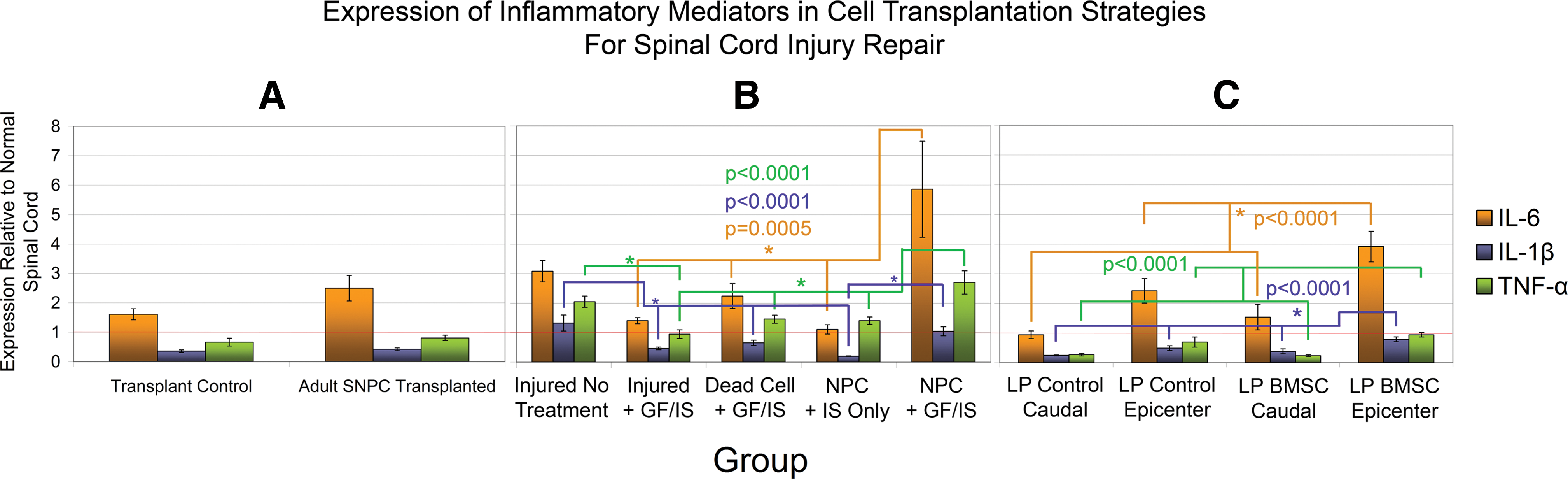
Cellular transplantation upregulates inflammatory mediators. Expression of inflammatory mediators IL-6, IL-1β, and TNF-α is plotted for Paradigms 1, 2, and 3 in
Discussion
Currently, there is little information about trophic factor expression in the normal and injured spinal cord, and even less is known about how cellular transplantation alters this expression. To date, the present study is the most extensive characterization of trophin expression in normal, injured, and transplanted spinal cord. Analysis of 3 distinct cellular transplantation paradigms provides valuable insights into how trophins could contribute to neural repair in this novel therapeutic strategy. Within each paradigm, strong conclusions can be made with regard to how experimental variables influence trophin expression; however, comparisons between paradigms should be cautiously made, as numerous confounds prevent similarly strong conclusions. To our knowledge, this study is also the first thorough characterization of a pure population of exogenous cells isolated post-transplant. Our findings suggest that trophins are poised to contribute to neural recovery after cellular transplantation for SCI; for NPCs, this was particularly true when transplantation was combined with trophin infusion in the intrathecal space.
Trophin expression is a valuable means of characterizing trophin production. qPCR is highly sensitive and able to detect minute levels of transcript inherent to trophin proteins that have biological activities at picomolar concentrations. Although one cannot be certain whether these levels of expression reflect protein levels, protein assays are not inherently superior for trophins, as Okragly et al. have demonstrated inaccuracies of protein assays related to tissue- and species-specific sequestration of trophin proteins [50].
Trophin expression in the adult, uninjured spinal cord
To date, few studies have characterized trophin expression in the normal and injured spinal cord. The expression of a few trophic factors has been studied, and many report data examining only one factor. The most commonly examined trophins include NGF, BDNF, and NT-3 [12,51 –67]. However, there is very little information regarding the expression of other trophic factors including bFGF, GDNF, CNTF, LIF, and TGF-β1 [12,13,48,49,52,62,68]. Therefore, the present study greatly expands knowledge of trophin production in the normal and injured spinal cord.
We found that all assayed trophins were expressed in the uninjured rat spinal cord. However, many of the factors were expressed at low levels relative to GAPDH. This low level of neurotrophin expression is in accordance with low levels reported by others in the adult spinal cord [52]. For example, Dougherty et al. demonstrated low levels of BDNF and NT-3 in astrocytes, oligodendrocytes, and microglia/macrophages in the normal spinal cord with immunohistochemical analysis [51].
Trophin expression in the injured spinal cord
Trophin levels after injury may play an important role in the normal reparative processes that follow an SCI. Marked upregulation of trophins was observed after injury to the peripheral nervous system, as compared with more limited upregulation after injury to the CNS—which has been postulated to account, at least in part, for the marked difference in their regenerative potential [12]. Similarly, trophin levels are reportedly lower in the injured spinal cords of adults as compared with neonatal animals, and it has likewise been postulated to at least partially explain the discrepancy in age-related regenerative responses [48].
To date, studies have reported highly variable findings related to trophin expression after SCI, likely resulting from varied injury models, injury severity, time points examined, species employed, and differences in tissue sampling and trophin assays [12,58,62]. Only a few studies have explored trophin expression more than a week after injury as we have done in the present study [48,56]. Most studies have shown upregulation of trophins after SCI [12,13,48,51,52,54,56,58 –60,62,63,65 –68], whereas others have reported a decrease in trophin levels after injury [48,59,61,62,66], or no change [48,52,57,58,64]. We found an upregulation of NGF, LIF, IGF-1, and TGF-β1 expression after injury in both models of Paradigms 2 and 3, suggesting that there is a robust trophic factor response in the weeks after compressive SCI. Indeed, NGF production is upregulated by the inflammatory response, which is well described after SCI [67,69 –71]. NGF may have both beneficial and deleterious roles in this context; although it stimulates survival of sympathetic and sensory neurons, it can also cause neuropathic pain, and its pro-form induces cell death [72]. LIF has been shown to be upregulated after CNS injury [68,73]. Although it induces cell survival [68,74 –78] and corticospinal tract growth [44], LIF increases astrogliosis [79 –82] and increases the inflammatory response [68]. LIF was also shown to increase the production of IGF-1 by activated microglia and macrophages [78] and, thus, it is noteworthy that IGF-1 was also consistently upregulated in our study. IGF-1 has been demonstrated to preserve neurons after SCI [83,84] and has been shown to increase the outgrowth of corticospinal axons [85]. TGF-β1 is an anti-inflammatory cytokine known to downregulate nitric oxide synthase [49,86]. TGF-β1 was shown to protect neurons from loss after an ex vivo model of CNS injury [87] and improved functional outcome after TGF-β1 infusion at the site of SCI [86]. This may relate to the ability of TGF-β1 to potentiate the effects of bFGF, CNTF and GDNF [88].
The differences noted for PDGF-A and VEGF-A expression between transplantation Paradigms 2 and 3 may be related to differences in the time from injury, differential presence of minocycline, cyclosporin, or differences inherent to distinct regions of spinal cord. Indeed, within the 3 paradigms, care was taken to compare equivalent regions of spinal cord, as there is a suggestion that trophin expression may vary with spinal cord level [62]. Nonetheless, the suggestion that VEGF-A may be downregulated in injured CNS tissue provides impetus for angiogenic therapeutic strategies for SCI [89].
Trophins could contribute to the benefits associated with cellular transplantation
Numerous groups have reported tissue sparing associated with cellular transplantation for SCI. Although mechanisms other than trophin secretion could account for this effect [10], it does raise the possibility that trophin secretion could be an important mechanism by which transplanted cells act. Interestingly, Keirstead's group have reported tissue sparing after the transplantation of both human oligodendrocyte progenitor cells (OPCs) and human motor neuron progenitor cells [21,90]. When the latter cells were transplanted, they also found sprouting of endogenous seritonergic projections and enhanced survival of endogenous neurons [90]. Bambakidis et al. have also reported white matter sparing associated with the transplantation of OPCs [22]. This raises the possibility that trophins produced by transplanted cells, or by the host in response to transplanted cells, could contribute to the functional benefits associated with cellular transplantation. However, rather than characterizing trophin production in vivo, numerous studies suggest that transplanted cells could have a trophic effect based on extrapolation of in vitro findings [21,90 –93].
Trophin changes associated with NPC transplantation
To date, a few studies have explored trophin production in tissue transplanted with neural cells [21,47,94]. Lu et al. reported expression of NGF, BDNF, and GDNF in tissue transplanted with C17.2 NPCs using immunohistochemistry and RT-PCR [47]. Yan et al. characterized the same 3 trophins and found similar results [94]. C17.2 are progenitor cells of the neonatal mouse cerebellum immortalized by transduction with the avian myc oncogene so, it is uncertain whether results from these cells will be similar to NPCs from other sources. In a different transplant paradigm, Sharp et al. found that OPCs produce TGF-β1 and neuregulins 1 and 2 in vitro and that hepatocyte growth factor was significantly upregulated in the region of the injury epicentre after OPC transplantation, while TGF-β2 was not significantly changed [21]. However, this study did not report expression data for any of the trophins investigated in the present study [21].
In the present study, Paradigm 1 found that NPC transplantation was associated with a significant reduction in BDNF and GGF2 expression in nonenriched perilesional tissue, while IGF-1 and CNTF expression were significantly increased. In Paradigm 2, NPC transplantation and pharmacotherapy alone was associated with significant perilesional downregulation of IGF-1 and TGF-β1 in nonenriched tissue. The difference in these results may stem from a different time post-injury or the differential presence of minocycline or cyclosporin. Since the magnitude of these differences in both paradigms are small, it is also possible that these differences reflect biological variability without biological significance.
Synergistic changes in trophin expression associated with co-administration of NPCs pharmacotherapy and trophin infusion
In Paradigm 2, numerous control groups allow a delineation of how different aspects of the applied combinatorial treatment strategy altered trophin expression. The effect of minocycline, cyclosporin, or trophin infusion on endogenous trophin production has not been previously reported to our knowledge. We have shown that animals treated with the anti-inflammatory and neuroprotective drugs minocycline and cyclosporin and intrathecal infusion of EGF, bFGF, and PDGF-A by implanted osmotic mini-pump after sham transplantation demonstrate modest but significant downregulation of EGF, bFGF, IGF-1, TGF-β1, and VEGF-A in perilesional spinal cord tissue. Downregulation of EGF and bFGF may represent negative feedback in the context of exogenous administration.
Transplantation of NPCs and pharmacotherapy alone was associated with significant downregulation of IGF-1 and TGF-β1 in perilesional tissue. The addition of a trophin infusion led to marked and statistically significant upregulation of the neurotrophins, GDNF, LIF and bFGF, demonstrating a synergistic effect of these combinatorial treatments. These expression changes could assist recovery from SCI. BDNF has been demonstrated to improve survival [95 –101] and regrowth [98,99] of motor neurons and to prevent apoptosis of oligodendrocytes [15]. NT-4/5 has similar effects to BDNF likely because both bind to the Trk B receptor [74,101,102]. NT-3 has similar effects [103,104] and may also enhance angiogenesis [105] and inhibit myelination [106,107]. GDNF has been demonstrated to promote the survival and outgrowth of many types of neurons [108,109]. bFGF is known to stimulate angiogenesis and the proliferation of NPCs in the adult spinal cord [13]. bFGF has also demonstrated neuroprotective effects on neurons [110] and oligodendrocytes [111] but was also shown to increase scar formation around CNS lesions [55]. The putative effects of NGF and LIF have been described above.
In this transplant paradigm, the presence of transplanted cells alone was associated with a significant increase in EGF and bFGF (when considering both the NPC + GF/IS and NPC + IS Only groups). However, trophin infusion was not associated with a statistically significant increase in these factors. EGF is a mitogen for many cell types that has also been shown to stimulate survival and outgrowth of neurons [112].
We hypothesized that the expression changes reported in transplanted cord homogenates predominantly reflect expression in host cells as exogenous cells were demonstrated to be a minority of the cells in this tissue. To explore this possibility and to characterize trophin expression in a pure sample of NPCs isolated from tissue one week after transplantation, we analyzed eYFP positive and negative specimens obtained by FACS sorting perilesional transplanted tissue from 3 NPC + GF/IS animals. Despite pooling cells obtained from 3 samples, trophin expression was near the limit of detection mandating careful interpretation. Careful interpretation is also warranted, because tissue harvest, dissociation, and FACS sorting may have influenced trophin expression; moreover, FACS may have introduced a selection bias for healthier cells—potentially those expressing higher than average trophin levels. These results suggest, however, that trophin changes in both host and transplanted cells may contribute to the trophin upregulation seen in the transplanted cord homogenate. Elevated trophin levels from both sources, thus, appear poised to contribute to neural repair processes.
Trophin changes associated with BMSC transplantation
Since neural differentiation of BMSCs has been unconvincing to date [4,113], trophin augmentation has seemed a likely mechanism of action for BMSCs especially when transplanted cells fail to survive [10]. Potential mechanisms by which BMSCs may function have been thoroughly reviewed by Parr et al. [10]. Consistent with a possible trophic effect, multiple groups have noted cell and tissue sparing or enhanced axonal regeneration after BMSC transplantation [23,24,114 –118]. Numerous groups have also noted reduced cyst formation after BMSC transplantation [8,9,119 –121]. Although BMSCs have been reported to produce NGF, BDNF, and VEGF in vitro [16,122,123], we are unaware of previous studies exploring trophin production by these cells after transplantation or by the host in response to their transplantation.
The present study demonstrates that intrathecal transplantation of BMSCs via lumbar puncture is associated with changes in trophin expression one week post-transplant (2 week postinjury) (Fig. 5). Here, BMSC transplantation significantly increased expression of NGF, LIF, IGF-1, and TGF-β1 relative to injured control tissue in distinct animals and significantly reduced expression of PDGF-A. Although the BMSC marker eGFP was also detected in caudal control tissue remote from the injury, which is consistent with the distribution of cells after intrathecal transplantation [35], there was no significant change in trophin expression in this uninjured tissue. This pattern of trophin upregulation is distinct from that seen resulting from NPC transplantation in either Paradigms 1 or 2. Interestingly, this is the same pattern of trophin alteration we twice demonstrated after SCI (Fig. 1). This raises the possibility that BMSCs, at least when transplanted via intrathecal infusion, may benefit by further upregulating trophins already upregulated by the SCI itself. A possible explanation for this effect would be an exacerbation of the inflammatory response known to follow SCI, prompting our analysis of inflammatory mediators in all 3 transplant paradigms.
Expression of inflammatory mediators following cellular transplantation
The inflammatory response that follows SCI has been thoroughly characterized [124]. IL-1, IL-6, and TNF-α are 3 commonly characterized inflammatory mediators known to be upregulated after SCI [49,125,126]. As shown in Fig. 6, increased expression of inflammatory mediators was demonstrated in injured spinal cord tissue relative to the uninjured cord. In Paradigms 1 and 3 (Fig. 6A, C) cellular transplantation increased the expression of inflammatory mediators relative to control tissue when both groups of animals were treated with cyclosporin. This did not reach statistical significance except for IL-1β in Paradigm 3. In Paradigm 2, immunosuppression with minocycline and cycosporin appeared to be effective as the expression of inflammatory mediators was lower in animals receiving these immunosuppressants than injured, untreated animals that did not receive these drugs. The exception, however, was the group treated with NPC transplantation and implantation of a trophin pump (NPC + GF/IS). Paralleling the synergistic increase in trophins in this group, a synergistic increase in inflammatory mediator expression was also seen. It is possible that this could be a physical or chemical effect of trophin infusion [127 –129]. Given the inter-relationship of NGF and the inflammatory cascade [67,71], it is possible that the increase in trophin response and the increase in inflammatory mediators is inter-related. Indeed, activated microglia and macrophages have been reported to produce NGF, NT-3, and bFGF [130]. It is also probable that the robust upregulation of NGF noted in the group transplanted with freeze-thaw killed NPCs stems from an immune response to this transplanted organic matter (Fig. 3B).
It is interesting to note that BMSCs have been reported by several groups to have anti-inflammatory properties [113] after transplantation. In the context of our findings, it is possible that the upregulation of the anti-inflammatory factor TGF-β1 is dominant over the upregulated pro-inflammatory cytokines IL-1β, IL-6 and TNF-α.
In conclusion, the present study significantly increases our understanding of trophic factor production in the normal and injured spinal cord. We have also thoroughly characterized trophin production in 3 preclinical cellular transplantation paradigms aimed at ameliorating the deficits of SCI. Although the biological activities of measured trophins is uncertain given the distinct effects of their pro-forms and binding to high- or low-affinity receptors, our work suggests that trophins could contribute to the benefits seen in all 3 transplantation paradigms. Future work will be required to determine whether this trophin elevation is responsible for the functional improvements seen, and if so, which trophins—alone or in combination—are responsible.
Footnotes
Acknowledgments
The authors wish to thank Drs. Xing-Hua Wang and Armand Keating for provision of BMSCs. The authors also wish to thank the Craig H. Neilsen Foundation, the Cervical Spine Research Society, and the Canadian Institutes of Health Research who provided funding for these experiments. G. Hawryluk was supported by fellowships from Johnson and Johnson Medical Products, the McLaughlin Center for Molecular Medicine, and the Congress of Neurological Surgeons/American Association of Neurological Surgeons.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.