
Abstract
Oral mucosal lamina propria progenitor cells (OMLP-PCs) are a novel, clonally derived PC population of neural crest origin with the potential to differentiate down both mesenchymal and neuronal cell lineages. In this study we aimed to determine the immunological properties of OMLP-PCs and to establish whether they would be suitable candidates for allogeneic tissue engineering and in the treatment of immune-related diseases. OMLP-PCs demonstrated no inherent immunogenicity with insignificant expression of costimulatory molecules (CD40, CD80, CD86, CD154, and CD178) or human leukocyte antigen (HLA) class II. OMLP-PCs required 7 days of stimulation with interferon-γ (IFN-γ) to induce cell surface expression of HLA II. Mixed lymphocyte cultures and mitogen stimulation demonstrated the potent immunosuppressive capability of OMLP-PCs in a contact-independent manner. Complete inhibition of lymphocyte proliferation was seen at doses as low as 0.001% OMLP-PCs to responder lymphocytes, while annexin V staining confirmed that this immunosuppressive effect was not due to the induction of lymphocyte apoptosis. These data demonstrate, for the first time, that OMLP-PC immunomodulation, unlike that for mesenchymal stem cells, occurs via a dose- and HLA II–independent mechanism by the release of immunosuppressive soluble factors and suggests these cells may have wide ranging potential in future immune-related therapies.
Introduction
O
Originating from biopsies of the oral buccal mucosa, these human PCs are isolated by differential adhesion to fibronectin and are clonally expanded, with each clone originating from a single cell. Clonal PC populations offer distinct advantages over heterogeneous stem cell populations [eg, mesenchymal stem cell (MSC) preparations] for stem cell therapy, as the cells can be more distinctly characterized, generating more reproducible results and giving a far more accurate indication as to how these cells would respond and differentiate upon, for example, transplantation. The advantages of such stem cell clonal sorting have been reported by Kotobuki et al., who demonstrated the enhancement of osteoblastic potential of sorted osteoblast-like cells for bone tissue engineering applications [2].
The potential for adult stem cells to be used for allogeneic tissue engineering applications has generated much interest recently, since many of these MSC sources have demonstrated significant immunosuppressive potential in vitro [3 –5]. Indeed, bone marrow MSCs (BMMSCs) have been known to modulate the immune system for many years, which are characterized by the expression of intermediate levels of human leukocyte antigen (HLA) class I and low levels of HLA class II and Fas Ligand. Furthermore, they do not express the costimulatory molecules CD40, CD80, CD86, or CD40L [6 –8]. The mode of action of MSC immunomodulation is likely to be multifactorial. Candidates such as indoleamine 2, 3-dioxygenase (IDO) and nitric oxide (NO) appear to be species specific, with only IDO being described as having a role to play in human MSC–mediated immunosuppression [9]. Despite the fact that no definitive mechanism of action has been determined, the ability of these cells to suppress the immune system is already being exploited in clinical trials for the treatment of immune-related disorders such as acute [10 –12] and chronic graft versus host disease (GVHD) [13] and in enhancing hematopoietic stem cell (HSC) transplantation [10]. However, an optimal stem cell–based therapy for the future treatment of GVHD remains dependent on further definition of the best stem cell source, dosage, and donor [ie, autologous (patient's own MSCs)/allogeneic (MSCs from the same donor as the HSCs)/third party donor (different donors for transplanted HSCs and MSCs)] [14].
Despite their obvious potential for allogeneic cell–based therapies, due to their immunoprivileged and immunomodulatory nature, BMMSC harvesting requires invasive surgery and it has been well documented that the numbers of BMMSCs decrease with age [15,16]. The isolation, therefore, of PCs from such an easily accessible and minimally invasive biopsy site (the buccal mucosa), which heals with no/minimal scar formation, offers distinct advantages over other current stem cell sources such as BMMSCs.
In this study we report, for the first time, the immunomodulatory effects of the recently described OMLP-PCs and their potential for use in allogeneic tissue engineering applications and in the treatment of immune-related disorders. This appears to be the first report of immunomodulation by a human neural crest–derived adult PC population and describes the potent immunosuppressive properties of OMLP-PCs in a dose- and contact-independent manner. This report demonstrates that OMLP-PCs can reduce lymphocyte proliferation in allogeneic mixed lymphocyte cultures (MLCs) at doses as low as 0.001% OMLP-PCs to responder cells, with complete inhibition of lymphocyte proliferation in Transwell cultures. These data demonstrate the importance of one/more soluble factors in exerting the immunomodulatory effects of OMLP-PCs on peripheral blood lymphocytes (PBLs). Hence, these cells offer a potentially preferential PC cell source for future allogeneic tissue engineering and for the treatment of immune-related disorders.
Materials and Methods
Isolation of PCs and maintenance in culture
Normal, disease-free buccal mucosa biopsies were obtained from consented patients (n=3) undergoing routine dental procedures at the School of Dentistry, University Hospital of Wales, Cardiff, United Kingdom. Local ethical committee approval had been previously obtained for this study and donors were informed in accordance with the Helsinki convention.
Single-cell suspensions were obtained following enzymatic digestion of the biopsies as previously described [17]. Briefly, tissue was incubated overnight at 4°C with dispase (Sigma; 2 mg/mL) to separate epithelial and lamina propria tissue. OMLP tissue was then disaggregated overnight at 37°C with Clostridium histolyticum A collagenase (Roche Life Sciences; 1 mg/mL).
PCs were separated by subsequent differential adhesion to fibronectin as previously described [1]. Briefly, OMLP single-cell suspensions in basal culture media (Dulbecco's modified Eagle's medium supplemented with 2 mM
Flow cytometry (FACS)
Expanded colonies (n=3) were grown±100 U/mL of interferon-γ (IFN-γ; Sigma) for 1, 2, or 7 days (just 7 days for FasL/CD178 and costimulatory molecules) before passaging by treatment with Accutase™ (Sigma) and counted. Cell suspensions were washed in phosphate-buffered saline (PBS) twice to remove residual serum before staining with HLA class I (ABC) R-Phycoerythrin (RPE; clone W6/32; 20 μg/mL), HLA class II (DP/DQ/DR) fluorescein isothiocyanate (FITC; clone CR3/43; 5 μg/mL; antibodies supplied by Dako UK Ltd.), FasL (CD178; clone Alf-2.1a; FITC; 20 μg/mL), and the costimulatory molecules CD40-FITC (clone EA-5; 5 μg/mL), CD154-FITC (clone 24–31; 10 μg/mL), CD80-FITC (clone P1.H5.A1.A1; 20 μg/mL), and CD86-FITC (clone BU63; 5 μg/mL; antibodies supplied by Ancell, Inc.). Cells were washed twice in PBS, fixed in 2% paraformaldehyde, and resuspended in 200 μL of PBS before detection. Cells were analyzed on an FACSCalibur™ flow cytometer (BD Biosciences) equipped with a 488 and 633 nm laser excitation source with 20,000 events recorded per sample. Fluorophore-conjugated immunoglobulins were used as controls. All data were compensated before analysis with FlowJo Version 7.2.2 software (Tree Star, Inc.).
Western blotting
The expression of HLA class II and IDO was investigated within expanded colonies (n=3) grown±100 U/mL of IFN-γ for 1, 2, or 7 days. Treated cells were lysed in 1% Triton® X-100, 0.25 M sucrose buffer supplemented with Complete™ protease inhibitor cocktail (Roche Applied Science). Lysates were sonicated before the addition of Laemmli buffer [62 mM Tris-HCl (pH 6.8), 10% glycerol, 5% sodium dodecyl sulfate (SDS), 0.005% bromophenol blue, and 5% β-mercaptoethanol] and boiled for 1 min. Five micrograms (10 μg for IDO blots) of total protein was loaded for each clone onto 10% (IDO) or 12% (HLA class II) SDS polyacrylamide gels (Invitrogen) and electrophoresis was performed. Total protein concentration was determined by bicinchoninic acid protein assay (Pierce) to ensure equal loading. Proteins were transferred to nitrocellulose membranes (Biorad) in transfer buffer containing 25 mM Tris, 192 mM glycine, and 20% (v/v) methanol overnight at 30 V at 4°C. Residual binding sites were blocked with 5% nonfat milk powder diluted in Tris-buffered saline with 0.1% Tween20™ (TBS-T) for 1 h with agitation at room temperature. Primary antibody [HLA class II (DP/DQ/DR), CR3/43; Dako; 1 μg/mL, or IDO, H-110; Santa Cruz Biotechnology, Inc.; 2 μg/mL] in 2% nonfat milk powder was added and incubated for 1 or 2 h (IDO) with agitation at room temperature. Membranes were subsequently washed repeatedly in TBS-T and incubated in appropriate horse-radish peroxidase conjugated secondary antibody (goat anti-mouse 1:3,000, Biorad; goat anti-rabbit 1:2,000, Dako) diluted in 2% nonfat milk for 1 h at room temperature with agitation. Membranes were washed in TBS-T twice and once in TBS before development using ECL™ and exposure to Hyperfilm (GE Healthcare).
MLCs and mitogenic stimulation
PBLs were prepared by centrifugation of heparinized blood on Ficoll-Isopaque (Lymphoprep™) and cultured in RPMI 1640 medium supplemented with penicillin (100 U/mL), streptomycin (100 μg/mL),
Transwell cultures
MLCs (n=3) were carried out using a 12-well plate Transwell culture system (BD Biosciences). Irradiated (20 Gy) OMLP-PCs (at 10% or 1% to responder cells)±prestimulation with IFN-γ for 7 days were seeded onto plate inserts with a 0.4 μm pore size membrane. PBLs (1×106) were mixed with an equal number of irradiated pooled PBLs in the bottom of the 12-well plate. Cells were cultured in RPMI 1640 medium supplemented with penicillin (100 U/mL), streptomycin (100 μg/mL),
Assessment of apoptosis
The viability of PBLs after MLC (n=3) was assessed by trypan blue exclusion. PBLs were cultured for 5 days in the presence/absence of OMLP-PCs±prestimulation with IFN-γ for 7 days. PBLs were harvested and resuspended in 0.4% trypan blue. Viable (exclude dye) and nonviable (include dye) cells were counted using a hematocytometer. Data are presented as percentage of viable cells compared with controls (A+Px).
Apoptosis was assessed by annexin V staining. PBLs were cultured for 3 days with 10 μg/mL PHA, in the presence/absence of irradiated OMLP-PCs (±prestimulation with IFN-γ for 7 days; n=3). PBLs were harvested and stained with LIVE/DEAD® fixable far-red dead cell stain kit (Invitrogen) for 30 min on ice, washed with PBS, and further incubated with annexin V-FITC (5 μL; FITC Annexin V Apoptosis Detection Kit I; BD Biosciences) for 15 min at room temperature. After washing with the provided binding buffer, cells were fixed in 2% paraformaldehyde, resuspended in 200 μL of PBS, and analyzed using an FACSCalibur™ flow cytometer (BD Biosciences) equipped with a 488 and 633 nm laser excitation source, with 30,000 events recorded per sample. Unstained and single-stained samples were used as controls. All data were compensated before analysis with FlowJo Version 7.2.2 software (Tree Star, Inc.).
IDO activity
IDO activity was determined by measuring the concentration of the tryptophan metabolite,
Statistical analysis
Where statistical analysis was appropriate, data were assessed for normality using the Anderson-Darling test (P>0.05) and statistically tested using a one-way ANOVA with Tukey-Kramer post hoc test. Where data did not demonstrate Gaussian distribution, a Kruskal Wallis test with a Dunn's post hoc test was employed. Significance was assumed at P>0.05.
Results
OMLP-PCs do not express HLA class II or costimulatory molecules
The cell surface expression (and intracellular in the case of HLA class II) of key markers of alloantigen presentation was measured using flow cytometry and western blotting (HLA class II only). Analysis was performed on OMLP-PCs cultured±100 U/mL IFN-γ for 1, 2, and 7 days to assess the effect of a proinflammatory stimulus on the expression of these markers.
Flow cytometry confirmed the constitutive expression of HLA class I on the cell surface of OMLP-PCs (Fig. 1ai) and its slight upregulation with exposure to IFN-γ after 1, 2, and 7 days (Fig. 1aii). In contrast OMLP-PCs were found to have no constitutive cell surface or intracellular expression of HLA class II (Fig. 1aiii, b). However, exposure to IFN-γ did induce intracellular HLA class II expression, as visualized by western blotting, within 1 day but this was not detected on the cell surface by flow cytometry until day 7 (Fig. 1aiv and b).
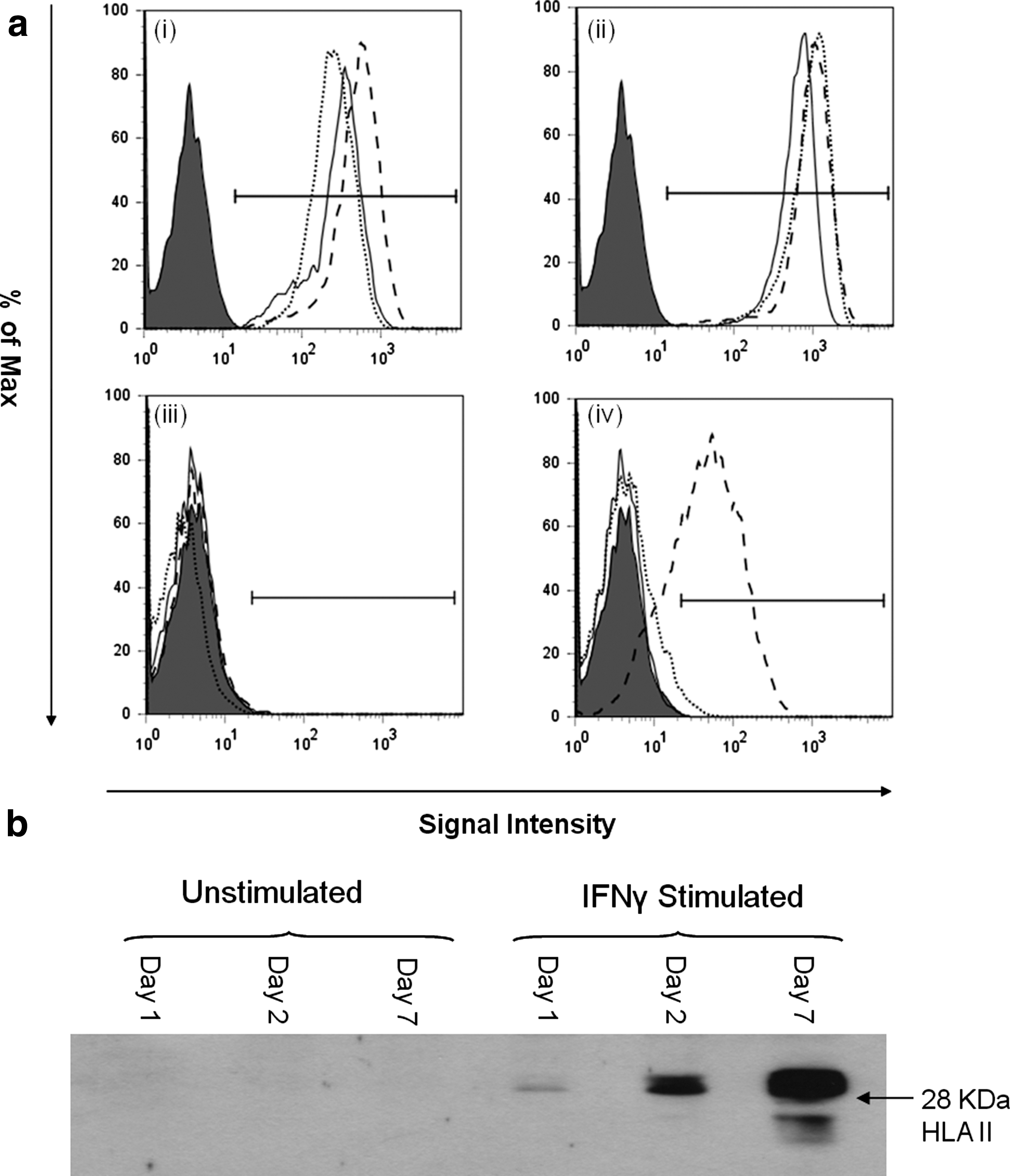
Oral mucosal lamina propria progenitor cells (OMLP-PCs) constitutively express human leukocyte antigen (HLA) class I but not HLA class II.
When analyzed by flow cytometry, OMLP-PCs did not express FasL (CD178) or the costimulatory molecule CD40 and its ligand CD154, CD80, or CD86 in the presence or absence of IFN-γ for 7 days (Table 1).
Flow cytometry was performed for the cell surface presentation of the costimulatory molecules CD40, CD80, CD86, CD154, and CD178 by oral mucosal lamina propria progenitor cells cultured±IFN-γ. Data are expressed as the average percentage of live positive cells±SD of the mean (n=3).
IFN-γ, interferon-γ; SD, standard deviation.
OMLP-PCs inhibit lymphocyte proliferation by mitogens and in MLC
The ability of OMLP-PCs to suppress the proliferation of PBLs was assessed by one-way MLC and stimulation by the T cell mitogen PHA. OMLP-PCs±prestimulation with IFN-γ for 7 days (to ensure cell surface HLA class II expression) were cocultured with PBLs and PHA or in one-way MLC for 3 or 5 days, respectively. MLC coculturing of responder cells (A) with OMLP-PCs±IFN-γ prestimulation did not significantly affect PBL proliferation compared with controls (A+Ax; Fig. 2a). Exposure of responder cells to irradiated PBLs from a pool of donors demonstrated a significant increase in PBL proliferation compared with controls (A+Ax; Fig. 2b; P≤0.001). The further addition of OMLP-PCs at proportions ranging from 10% to 0.001% (equivalent of 1×104–1 cell/reaction) of responder cells significantly inhibited the proliferation of the PBLs (P≤0.001) in a dose-independent manner to levels comparable with A+Ax controls (Fig. 2b). Positive controls (A+Px) confirmed that this dose-independent effect was not due to an artifact of the system.
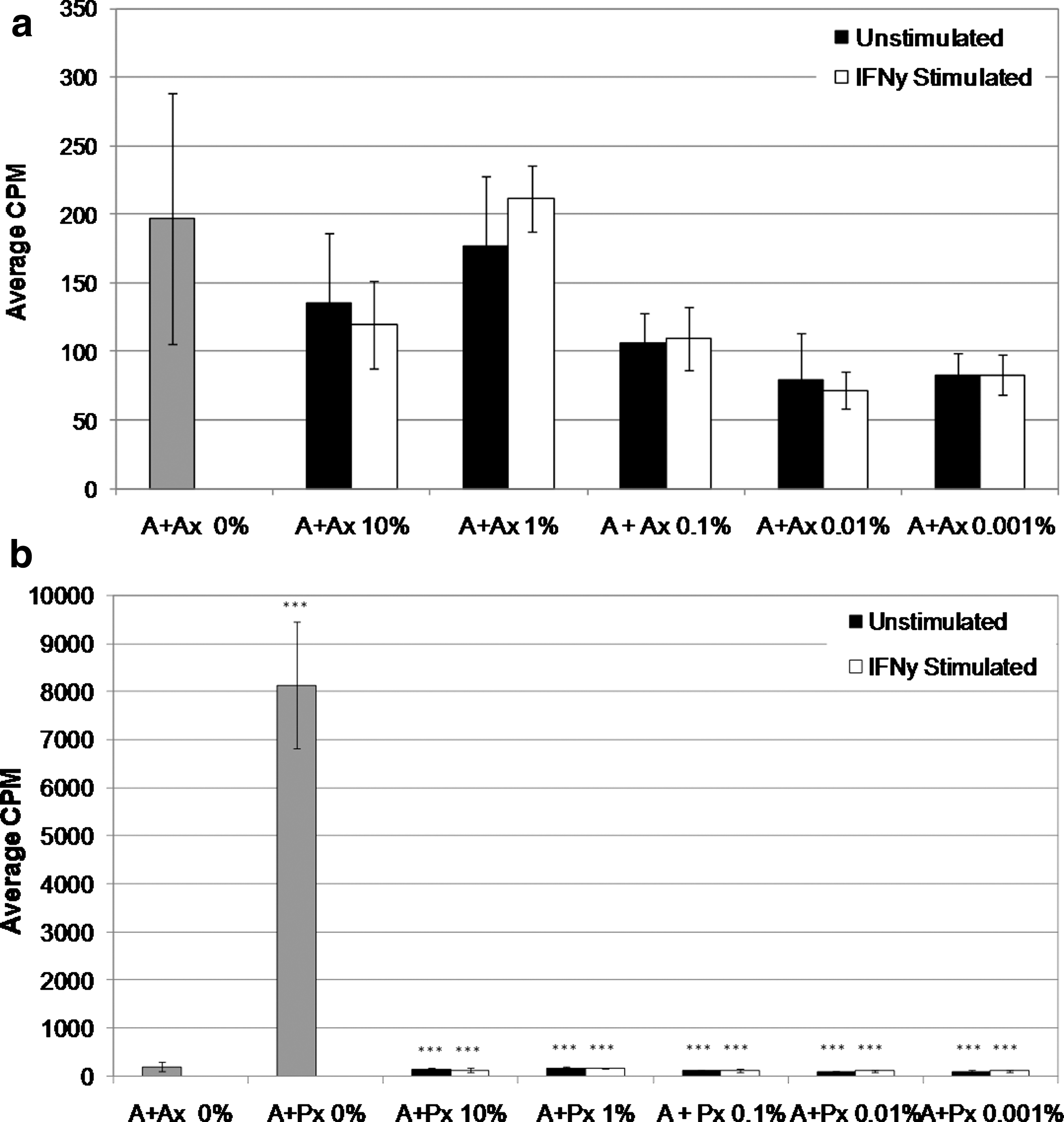
OMLP-PCs suppress the proliferation of lymphocytes in a dose- and HLA-independent manner. OMLP-PCs (10%–0.001% to responder cells)±prestimulation with IFN-γ for 7 days were cocultured with peripheral blood lymphocyte (PBLs) for 5 days. Responder cells (A) were cocultured with irradiated (x)
A similar effect was also observed with mitogen (PHA) stimulation of the PBLs. Addition of OMLP-PCs at both 10% and 1% PCs of responder cells significantly decreased PBL proliferation (P≤0.001) back to levels comparable with the baseline control (A), demonstrating the ability of OMLP-PCs to respond to both allogeneic and mitogenic stimuli (Fig. 3).
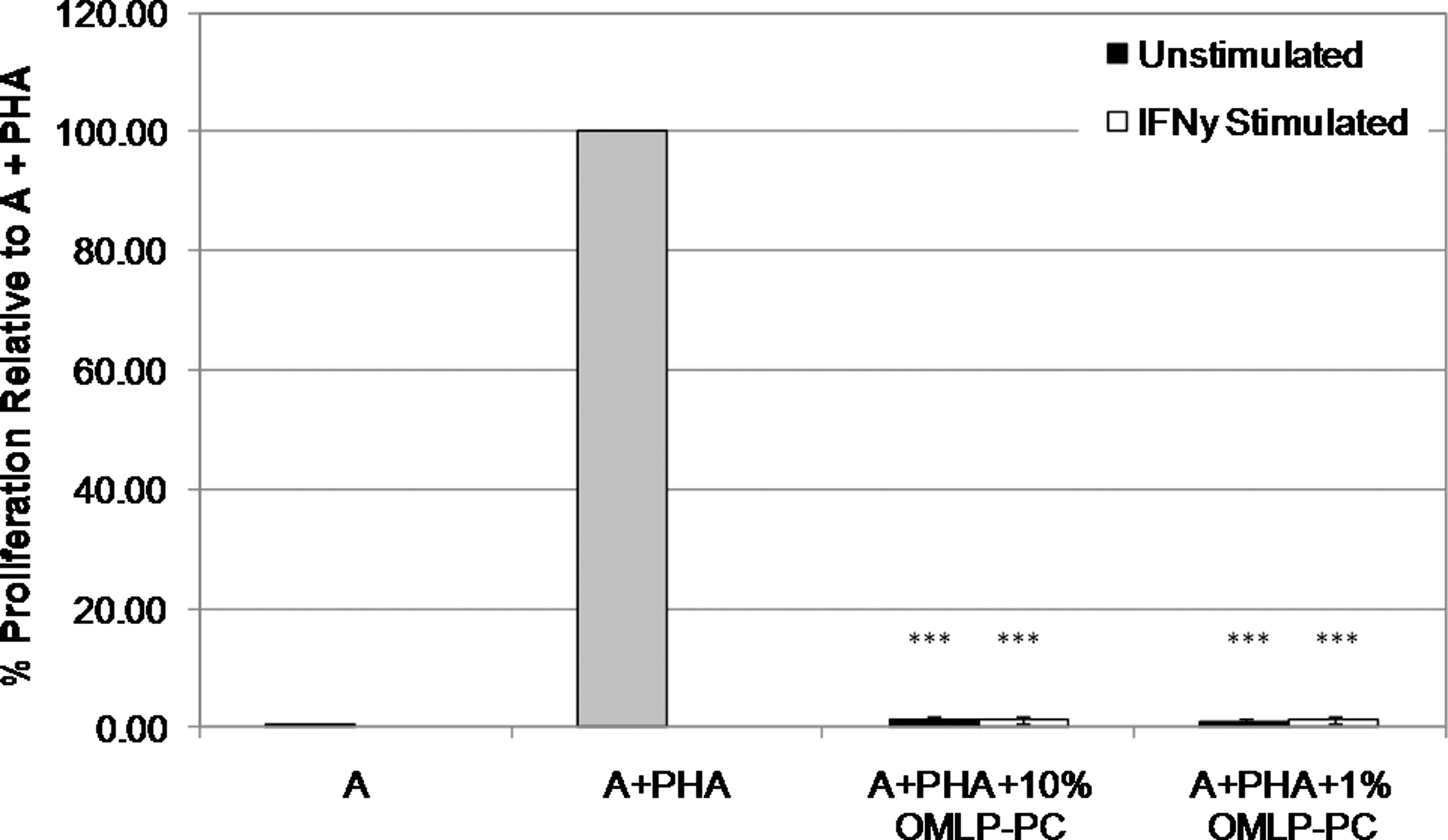
OMLP-PCs suppress the proliferation of lymphocytes stimulated by the T cell mitogen phytohemagglutinin (PHA) in a dose- and HLA-independent manner. OMLP-PCs±prestimulation with IFN-γ for 7 days were cocultured with PHA-stimulated PBLs for 3 days. PHA-stimulated controls have been set at 100% to account for variation between experiments. Data are expressed as average percentage proliferation compared with A+PHA controls±SD of the mean (n=3); ***P≤0.001.
Induction of HLA class II expression of the cell surface of the OMLP-PCs (via stimulation with IFN-γ prior to MLC or mitogen stimulation) did not significantly affect the suppression of PBL proliferation when compared with unstimulated OMLP-PCs, suggesting that OMLP-PC immunosuppression occurs via a HLA class II–independent mechanism (Figs. 2 and 3).
OMLP-PCs inhibit lymphocyte proliferation in a contact-independent manner
To determine whether OMLP-PC–induced inhibition of PBL proliferation was via a contact-dependent or -independent mechanism, one-way MLCs were set up with OMLP-PCs in direct contact with the PBLs and separated by a Transwell membrane. The addition of OMLP-PCs at 10% and 1% of responder cells, in direct contact or Transwell cultures, significantly inhibited PBL proliferation compared with controls (P≤0.001; Fig. 4) to levels comparable with baseline controls (A). No significant differences in the level of inhibition were seen between control (A) and Transwell groups, suggesting that OMLP-PCs are able to completely suppress the proliferation of PBLs through the release of soluble factors, requiring no direct contact with the responder cells. The release of these soluble factors was also dose and HLA class II independent.
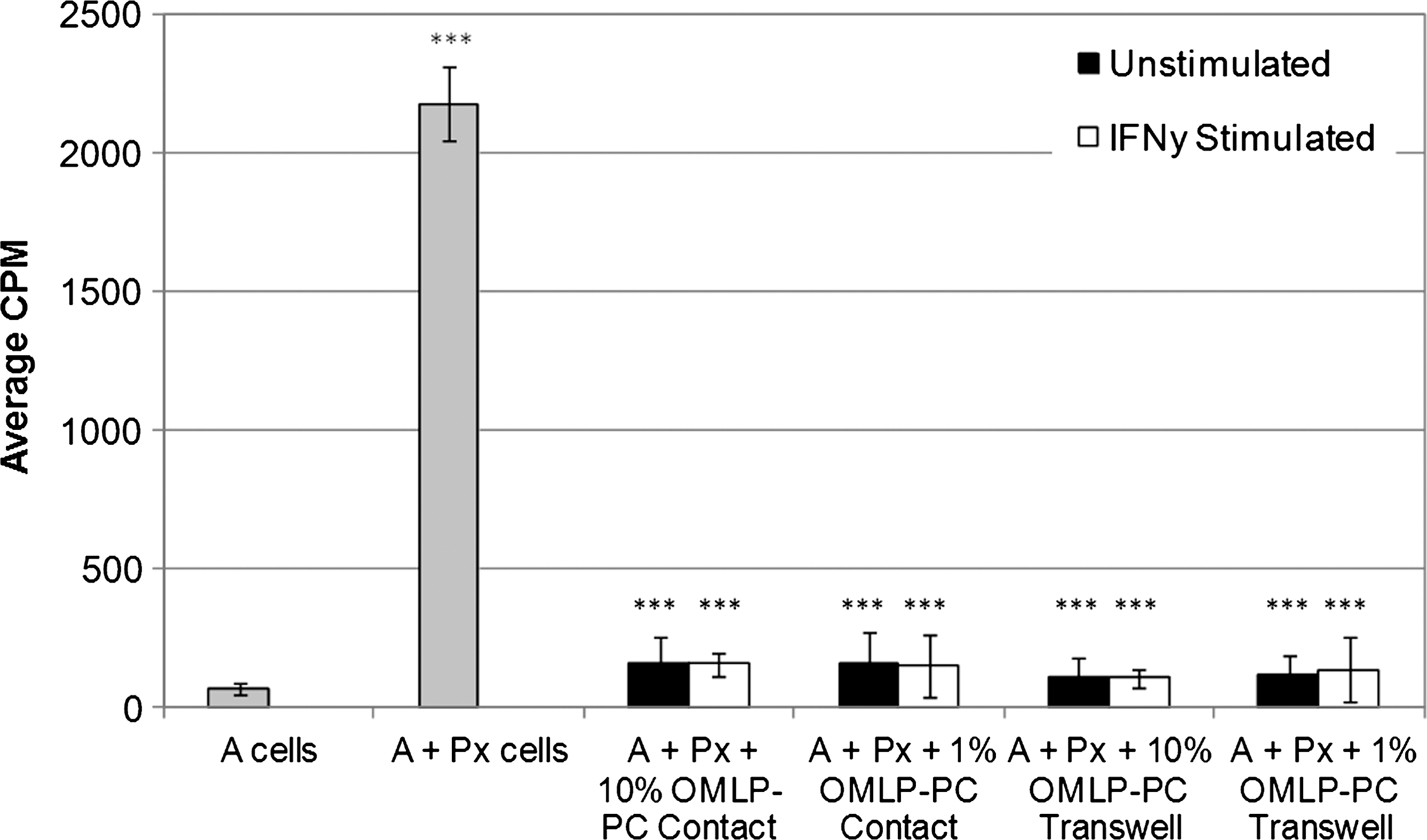
OMLP-PCs suppress the proliferation of lymphocytes through the release of soluble factors. OMLP-PCs±prestimulation with IFN-γ for 7 days were cocultured with PBLs in direct contact and in Transwell systems for 5 days. Responder cells (A) or A cells cocultured with PBLs pooled from 5 different donors (Px) acted as baseline and positive controls, respectively. Data are expressed as average CPM±SD of the mean (n=3); ***P≤0.001.
OMLP-PCs do not inhibit lymphocyte proliferation by inducing cell death
PBL viability was assessed after one-way MLC in the presence of OMLP-PCs to confirm that the inhibition of PBL proliferation by the PCs was not due to an induction of cell death. PBLs were harvested after 5 days in MLC and their viability was assessed by trypan blue exclusion. No significant difference in PBL viability was seen after MLC in the presence of OMLP-PCs at 10% or 1% to responder cells (A)±prestimulation with IFN-γ compared with the control (A+Px; Fig. 5a). The induction of PBL apoptosis was also investigated by annexin V staining. PBLs cultured for 3 days in the presence of PHA and OMLP-PCs (±prestimulation with IFN-γ) at 10% to responder cells were stained with annexin V and LIVE/DEAD® fixable cell stain. Flow cytometry confirmed a significant decrease in the percentage of annexin V+ LIVE/DEAD®− apoptotic PBLs on coculture with OMLP-PCs compared with PHA-treated PBL controls (P≤0.05; Fig. 5b). No significant difference was seen in the numbers of necrotic PBLs between treatments, confirming that OMLP-PCs do not induce PBL immunosuppression through induction of cell death, but rather appear to protect PBLs against entry into apoptosis.
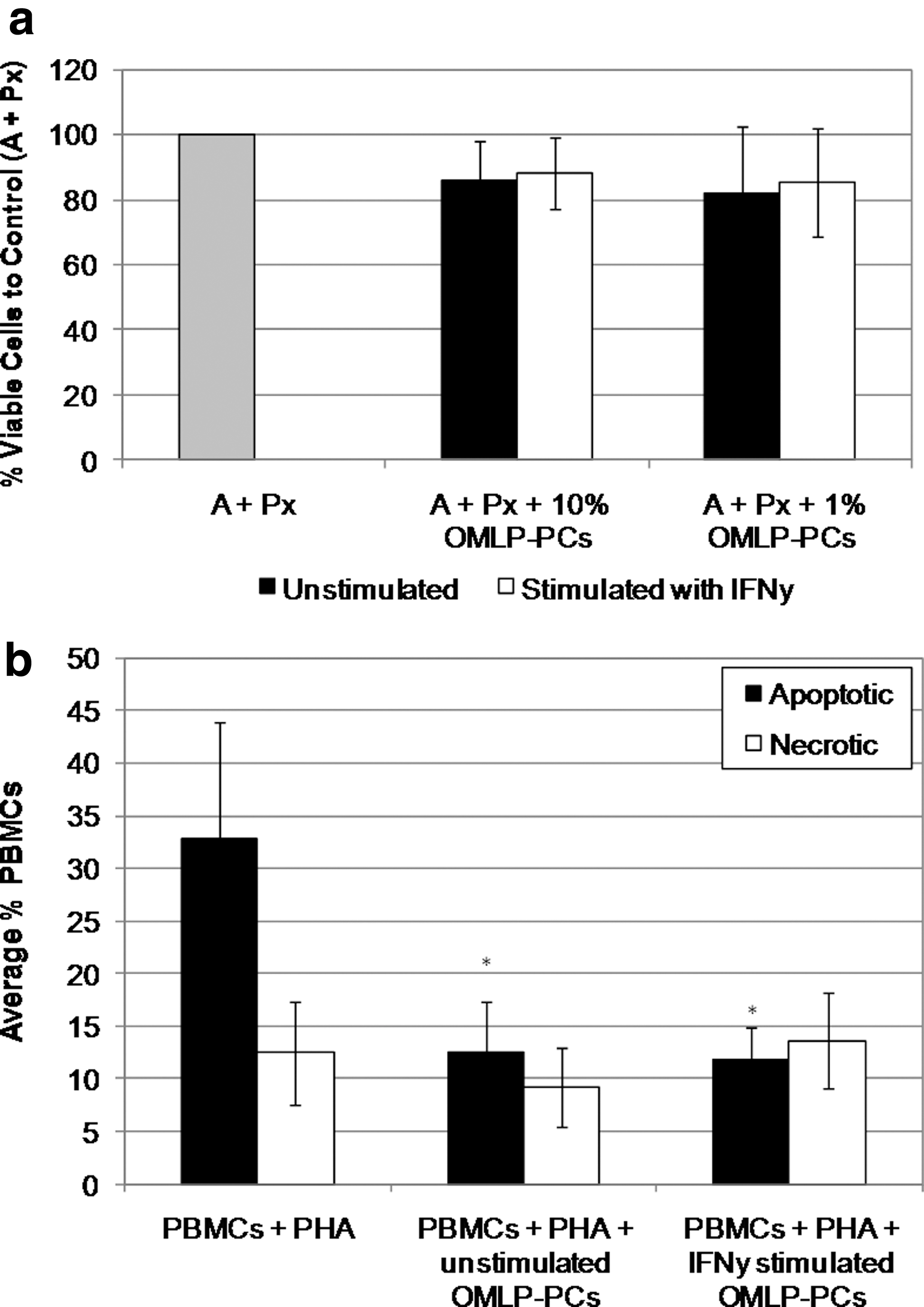
OMLP-PCs do not immunosuppress through the induction of PBL cell death.
IDO expression and activity is induced in OMLP-PCs exposed to allogeneic lymphocytes
IDO has been implicated in immunosuppression in a number of tissue environments including the immunosuppression of tumors and in maternal tolerance of the fetus [20,21]. It has also recently been demonstrated to be one of the potential mechanisms in human BMMSC immunosuppression [9]. The expression of IDO by OMLP-PCs±IFN-γ for 1, 2, and 7 days was investigated by western blotting. OMLP-PCs were demonstrated not to express IDO unless exposed to IFN-γ for a minimum of 48 h (Fig. 6a). IDO activity within the conditioned media of OMLP-PCs±IFN-γ exposure for 7 days and also after one-way MLC (conditioned media collected from the Transwell experiment in Fig. 4) was investigated by the measurement of the tryptophan metabolite,
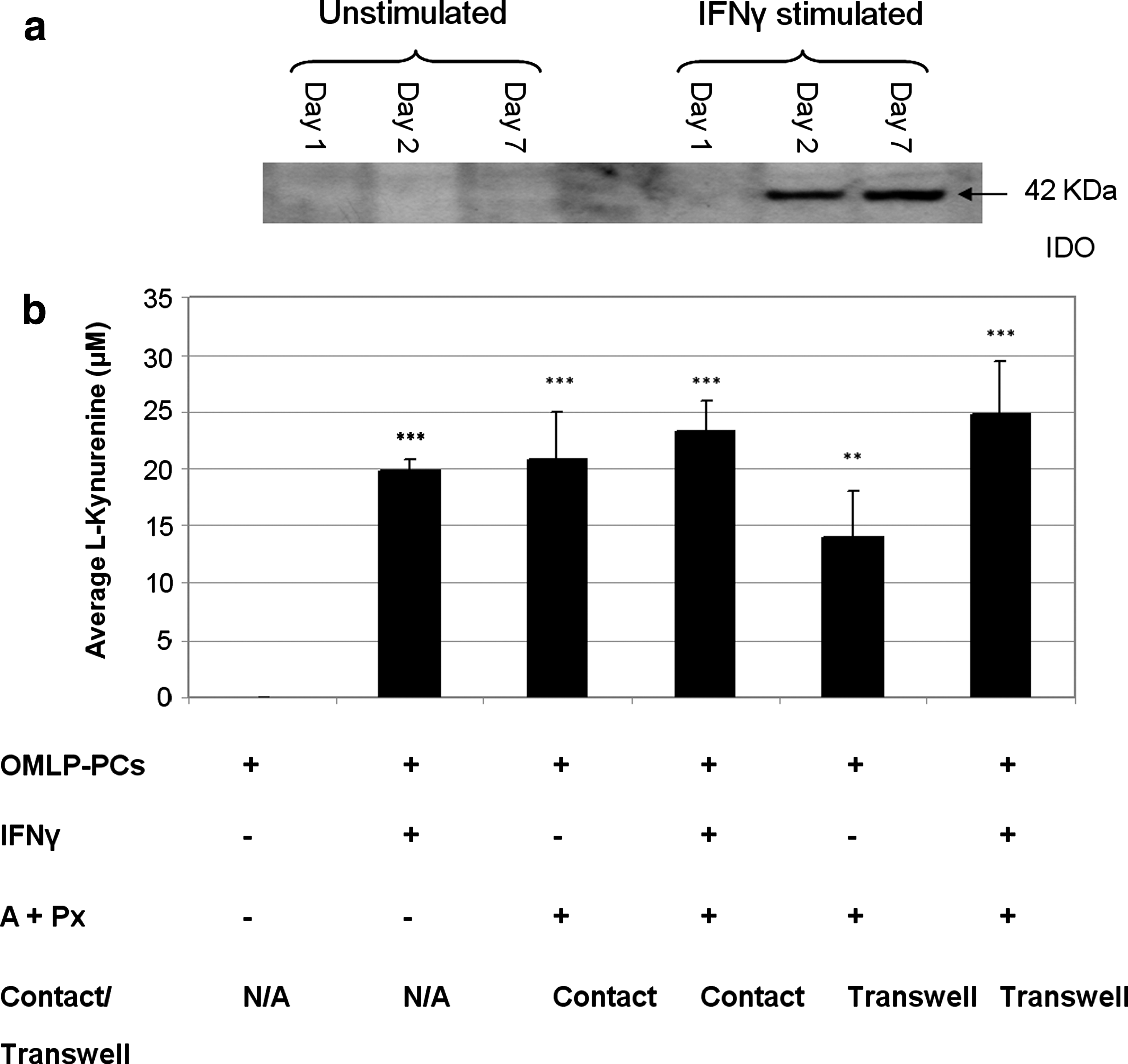
OMLP-PCs induce the expression and activation of indoleamine 2, 3-dioxygenase (IDO) in response to stimulation with IFN-γ and/or coculture with PBLs.
Discussion
The potential for BMMSCs in immunomodulation of the immune system has long been appreciated. The nonimmunogenic nature and ability of MSCs to modulate the immune system from a number of tissues has been documented [3,4,22]. Numerous studies have illustrated the ability of human MSCs to inhibit, in vitro, T and B cell proliferation, differentiation, and chemotaxis as well as promoting the expansion of CD4+CD25+FoxP3+ regulatory T cells [23 –25]. Their immunosuppressive nature within allogeneic cocultures in vitro is of clinical relevance for use in allogeneic transplantation, to reduce both the incidence and severity of GVHD. Frassoni et al. [26] suggested that cotransplantation of MSCs and HSCs from HLA-identical siblings resulted in a significant decrease in acute and chronic GVHD. However, MSC heterogeneity means that trial results are variable and also large cell numbers are required to deliver an immunosuppressive effect that can sometimes initiate an immune response. Such issues may potentially be overcome through the use of clonally derived OMLP-PCs due to their potent immunosuppressive effects and the fact that the tissue from which they are isolated is easily accessible/minimally invasive and heals rapidly with no/minimal resultant scar formation for the patient. In addition, the use of BMMSCs for cellular therapeutics is not always suitable, due to the significant decreases in cell number and proliferation with age and the highly invasive procedure for obtaining the cells [27].
In this study the potent immunosuppressive capabilities of OMLP-PCs have been demonstrated. These cells are clonally expanded, originating from a single PC, thereby generating a homogenous cell population that is well characterized and which has both predictable and reproducible results. Our previous work has already demonstrated the expression of numerous stem/PC markers on the cell surface of OMLP-PCs [1]. These cells have a similar profile to BMMSCs in their characterization as CD90+, CD105+, CD166+, CD44+, CD34−, and CD45−[6]. Further analysis has also confirmed that these cells are CD29+ and CD73+ (data not shown). OMLP-PCs however, demonstrate distinct differences in terms of their developmental origin to BMMSCs, originating from the neural crest they express markers such as Slug, Snail, Sox10, and Twist [1]. FACS analysis confirmed that OMLP-PCs demonstrate a similar immunophenotype to BMMSCs, constitutively expressing HLA class I, but HLA class II negative [4,22]. As reported for BMMSCs [28,29], insignificant levels of the costimulatory molecules CD40, CD80, CD86, CD154, and CD178 were demonstrated on the cell surface of OMLP-PCs cultured in the presence/absence of an inflammatory environment (IFN-γ for 7 days). In contrast to BMMSCs, which express cell surface HLA class II within 2 days of IFN-γ treatment [22], HLA class II expression was not apparent on the cell surface of OMLP-PCs until day 7 of IFN-γ treatment. Western blotting confirmed that unstimulated OMLP-PCs do not synthesize any HLA class II. Intracellular expression was detectable after 1 day of stimulation, with this taking a further 6 days to appear at the cell surface. These results demonstrate significant differences to those previously reported for BMMSCs that constitutively express intracellular HLA class II [22]. The data in this study demonstrate distinct similarities to that previously documented for fetal MSCs [30] as opposed to adult MSC populations, thereby suggesting that OMLP-PCs have a more fetal phenotype with respect to immune recognition. This is corroborated by our previous findings that have demonstrated the “embryonic-like” phenotype of OMLP-PCs in their expression of embryonic markers such as Oct4a and Nanog and their expression of active telomerase [1]. This HLA and costimulatory molecule expression profile confirms that human OMLP-PCs are not inherently immunogenic.
One-way allogeneic MLCs clearly demonstrated the potent immunosuppressive action that OMLP-PCs have. These experiments demonstrated that OMLP-PCs are able to suppress PBL proliferation back to baseline control levels in a dose- and HLA class II–independent manner. This is in contrast to BMMSCs where pre-exposure to IFN-γ has been demonstrated to increase their inhibition of PBL proliferation [22,31]. This is the first time that a PC population has been demonstrated to immunosuppress to this extent and in such a dose-independent manner. Numerous stem cell populations including BMMSCs, periodontal ligament stem cells, and human umbilical cord perivascular cells have demonstrated a dose-dependent immunosuppressive effect in MLC, with an optimum proportion of 10% stem cell to responder cells [3,4,22] (potentially 10,000× less effective than OMLP-PCs, although a direct comparison would need to be undertaken). Indeed it has previously been demonstrated that a low concentration of BMMSCs in MLC in fact increases, as opposed to suppressing, T cell proliferation in a HLA-independent manner [32]. Recently Najar et al. have suggested that this stimulatory effect is IFN-γ and HLA class II independent [33]. In this study we demonstrated complete suppression of PBL proliferation even at 0.001% OMLP-PCs to responder cells. These results may be in part due to the clonogenic nature of OMLP-PCs resulting in a reproducible, potent immunosuppressive effect between patients.
Analysis of PBL viability subsequent to MLC confirmed that the observed immunosuppressive effects are due to a true inhibitory effect rather than simply due to the induction of apoptosis. These findings have implications in the clinical application of stem cell populations for both allogeneic tissue engineering applications as well as in the treatment of immune-related disorders such as GVHD where BMMSCs are currently the preferred cell source. In such clinical investigations, due to the differential response of BMMSCs at different concentrations, dosing for stem cell therapy has become a significant concern [14]. These results with extremely low doses of OMLP-PCs reported here suggest that the use of such oral progenitors would be of distinct clinical benefit.
Repetition of these MLCs in both contact and Transwell systems demonstrated that OMLP-PCs exert their immunosuppressive effects through the release of soluble factors. Indeed, no direct cell–cell contact was required for complete immunosuppression to be demonstrated. This contact-independent effect was also HLA class II independent. Controversy currently exists in elucidation of the mechanism of action by which MSCs exert their immunosuppressive effects. It has been previously reported that although suppression of lymphocyte proliferation does occur in a contact-independent system (suggesting the importance of soluble factors), cell–cell contact is also necessary for complete inhibition [33,34]. These findings have recently been supported by data confirming that periodontal ligament stem cells are only able to partially suppress the proliferation of PBLs in contact-independent coculture [4]. Such results suggest that OMLP-PCs may act via a different mechanism of immunosuppressive action to MSCs.
IDO and NO are both soluble factors known to have immunosuppressive effects [9,35]. IDO is known to have roles in immunosuppression in tumor development and in the immunoprivileged environment of the cornea [20,21]. Specifically IDO has also been suggested to be one of the most important soluble factors in human MSC immunomodulation with NO being suggested to have more of a role in murine MSC immunomodulation [9]. IDO is the enzyme involved in catalyzing tryptophan degradation, producing metabolites such as
This study has demonstrated for the first time the potent immunological properties of OMLP-PCs. The ability of OMLP-PCs to potently suppress the proliferation of PBLs in a dose- and contact-independent manner has wide ranging ramifications for clinical therapies/applications. We have previously demonstrated the advantages of this cell source—offering a multipotent PC population, capable of differentiation down both mesenchymal and neuronal lineages after minimally invasive and reproducible isolation [1]. The ability of OMLP-PCs to suppress lymphocyte proliferation lends them to uses in allogeneic tissue engineering applications, as well as their obvious potential in the direct treatment of immune-related disorders. These cells offer direct advantages over currently utilized BMMSCs in that not only would fewer cells be required to induce immunosuppression (hence a significant impact on scale up and cost for development as a therapeutic) but cells can be reliably isolated from an easily accessible biopsy site with no/minimal scarring for the donor. Furthermore, the clonogenic nature of this cell source, in comparison to heterogeneous populations of BMMSCs, means that reproducible, predictable results and clinical end points could be achieved.
Footnotes
Acknowledgments
The authors gratefully acknowledge funding from the Wellcome Trust, the Medical Research Council (MRC; G0901562), National Institute for Social Care and Health Research (HA09-034), the British Society of Oral and Dental Research (BSODR), and the Cardiff Institute of Tissue Engineering and Repair (CITER). Buccal mucosal biopsies were kindly obtained from patients undergoing routine dental surgery by Mr. Andrew Cronin, Consultant Oral and Maxillofacial Surgeon, University Hospital of Wales, Cardiff. This work is subjected to patent filing (Patent Application Nos. GB0811865.5, GB810841.7, GB0820012.3, PCT/GB2009/001443, AU2009259053, EP09761964.7, CN200980122121.0, JP501125275, and US12/997,363).
Author Disclosure Statement
No competing financial interests exist.