
Abstract
The derivation of pluripotent embryonic stem cells (ESCs) from a variety of genetic backgrounds remains a desirable objective in the generation of mice functionally deficient in genes of interest and the modeling of human disease. Nevertheless, disparity in the ease with which different strains of mice yield ESC lines has long been acknowledged. Indeed, the generation of bona fide ESCs from the non obese diabetic (NOD) mouse, a well-characterized model of human type I diabetes, has historically proved especially difficult to achieve. Here, we report the development of protocols for the derivation of novel ESC lines from C57Bl/6 mice based on the combined use of high concentrations of leukemia inhibitory factor and serum-replacement, which is equally applicable to fresh and cryo-preserved embryos. Further, we demonstrate the success of this approach using Balb/K and CBA/Ca mice, widely considered to be refractory strains. CBA/Ca ESCs contributed to the somatic germ layers of chimeras and displayed a very high competence at germline transmission. Importantly, we were able to use the same protocol for the derivation of ESC lines from nonpermissive NOD mice. These ESCs displayed a normal karyotype that was robustly stable during long-term culture, were capable of forming teratomas in vivo and germline competent chimeras after injection into recipient blastocysts. Further, these novel ESC lines efficiently formed embryoid bodies in vitro and could be directed in their differentiation along the dendritic cell lineage, thus illustrating their potential application to the generation of cell types of relevance to the pathogenesis of type I diabetes.
Introduction
M
A number of different strategies and refinements have been implemented to improve the efficiency of existing protocols. These include the use of hybrid embryos between permissive (129, C57Bl/6) and nonpermissive (CBA/Ca, NOD) strains [7,8], strategies for genetic modification of either the zygote [7] or fibroblast feeder cells [9], or the optimization of media for stem cell growth [10] by the addition of cytokines [11] or small-molecule inhibitors [12 –14]. In addition, technical advances, such as explanting an immunosurgically isolated inner cell mass (ICM) [2] or microsurgically isolated epiblast [15], have improved the efficiency of ESC derivation from some, but not all strains, the NOD mouse remaining largely intractable, despite such interventions.
Fetal bovine serum (FBS) serves as a source of the soluble growth factor, bone morphogenetic protein 4 (BMP4), which is essential for ESC derivation, but significant variability between batches of FBS has led to the development of alternatives, such as KnockOut serum replacement (KSR; Invitrogen). This chemically defined serum-free substitute for FBS has been used in several studies reporting improved establishment of C57Bl/6J ESC lines [10,16] and has been found to be more effective than serum-containing medium for obtaining induced pluripotent stem cells (iPSC) from mouse embryonic and adult fibroblasts [17].
One of the most significant findings for the optimal maintenance of ESCs in culture was the requirement for recombinant murine leukemia inhibitory factor (rmLIF) [18,19], which is typically used at a concentration of 103 U/mL. LIF is an interleukin-6 (IL-6) family multifunctional cytokine that binds to a cell surface complex composed of the LIF receptor, LIFRβ, and the transmembrane signaling molecule gp130, thereby resulting in activation of the transcription factor signal transducers and activators of transcription (STAT)-3, which is essential and sufficient for suppression of mouse ESC differentiation [18,20,21]. The LIF/STAT3 pathway combines with the Wnt/GSK3β/β-catenin pathway to promote self-renewal by activation of pluripotency genes, including the transcription factors Nanog, Oct3/4, and Klf4. LIF has been reported to enhance in vitro preimplantation embryo development in several different species including mice [22], cows [23], and sheep [24], although its role in humans has yet to be resolved [25,26]. There have been several reports of improved rates of ESC derivation from refractory strains when LIF has been present at concentrations higher than those used for routine culture [13,27,28]. Indeed, strain difference in response to soluble growth factors is evident from the original reports of mouse ESC derivation: Evans and Kaufman were able to establish ESC lines from permissive 129SvE embryos solely by culturing on fibroblast feeder cells, whereas Martin was not able to generate ESC lines from (ICR x SWR/J)F1 and (C3H x C57BL/6)F1 embryos unless conditioned medium, equivalent to a 5-fold concentration of LIF, was also added. More recently, derivation rates of between 5% and 66% efficiency across 10 different inbred strains, including the previously nonpermissive CBA/CaOla mouse, were achieved by culturing blastocysts in conditioned medium from genetically modified fibroblasts, secreting rabbit LIF at 15 ng/mL [9].
Despite these refinements, the rates of ESC derivation for many strains remains unimpressive. Recently, the use of small molecules (“2i”) that inhibit critical signaling pathways has enabled the generation of germline-competent ESCs from the most refractory strain of all, the NOD mouse [14]. This was achieved by simultaneously promoting proliferation and self-renewal via glycogen synthase kinase 3 (GSK3)-inhibition and preventing spontaneous differentiation by the exclusion of exogenous signaling factors provided by trophoblast tissue, feeder cells, or FBS and chemically suppressing differentiation via mitogen-activated protein kinase/extracellular signal-related kinase (MEK/ERK) inhibition. Nevertheless, such an approach resulted in a high incidence of aneuploidy among ESCs and their limited capacity for in vitro differentiation, as evidenced by the failure to form embryoid bodies (EBs) in culture [29]. We present here a simple and inexpensive protocol for the efficient derivation of novel ESC lines from refractory strains, including CBA/Ca and NOD, which does not require chemical or microsurgical perturbation. Using this approach, ESC lines with normal karyotype and germline competence can be established at high rates of efficiency and maintained in standard ESC culture media. Further, we demonstrate that the resulting NOD ESCs can differentiate normally in vitro, via the formation of EBs, and can be directed to differentiate into dendritic cells (DC), implicated in the pathogenesis of type I diabetes.
Materials and Methods
Derivation and culture of ESC lines
Mouse embryonic fibroblasts (MEFs) were prepared from day 13 to 14 embryos [30] and used to make feeder cells for ESC derivation by mitotic inactivation through exposure in culture to 1 μg/mL solution of mitomycin C (MMC) (Sigma, Cat# M4287) in Dulbecco's modified Eagle's medium (DMEM) (Biowhittaker, Cat# BE12-604F) plus 10% FBS (Gibco, Cat# 16000) supplemented with 2 mM L-glutamine (PAA, Cat# M11-004), 50 U/mL penicillin, 50 μg/mL streptomycin (PAA, Cat# P11-010), 1 mM sodium pyruvate (PAA, Cat# S11-003), 0.1 mM MEM nonessential amino-acids (PAA, Cat# M11-003), and 0.2 mM 2-mercaptoethanol (Sigma, Cat# M7522) (Feeder medium), for 2 hours. After 3 washes in Dulbecco's phosphate-buffered saline (DPBS) (Gibco, Cat# 14190), feeder cells were plated at a density of 5×104 cells per cm2 in feeder medium.
Embryos used for ESC derivation were recovered from the uteri of day 4 pregnant females that had been maintained on a 14 h light: 10 h dark cycle and naturally mated, with the exception of C57Bl/6 and NOD/ShiLtJ mice. These embryos were supplied as 4-cell to 8-cell embryos cryopreserved under liquid nitrogen (The Jackson Laboratory) and were each thawed by rapidly passing through 10 volumes of DPBS and cultured overnight in modified synthetic oviductal medium enriched with potassium and containing 4 mg/mL bovine serum albumin (BSA; Sigma, Cat# A3311) (mKSOM) [31]. HEPES-buffered mKSOM medium was used to flush morulae and early blastocysts from uterine horns on the afternoon of the 4th day after detection of copulation plugs. Embryos for ESC derivation were plated one embryo per well into Nunclon Δ 4-well plates (Nunc, Cat# 176740) that had been seeded 1 day earlier with mitotically inactivated MEFs at passage 3 (p3). The embryos were cultured (humidified at 37°C, 5% CO2) in 0.5 mL per well of Knockout DMEM (KO DMEM) (Invitrogen, Cat# 10829) supplemented as just described containing 15% KSR (Invitrogen, Cat# 10828) and 5×103 U/mL rmLIF (Chemicon Int., ESGRO, Cat# ESG1107) (KSR/5× LIF medium) for between 5 and 7 days until substantial epiblast outgrowths had formed. During this period, embryos equivalent to 3.5 day development at time of plating formed fully expanded blastocysts within the first 24–36 h of culture. Any blastocysts that failed to hatch from their zona pellucida within the next 24 h were enabled to do so by brief exposure to acidified Tyrodes solution (Sigma, Cat# T1788). Hatched blastocysts would typically attach within 24 h and form epiblast outgrowths suitable for picking within the next 48 to 72 h.
Healthy epiblast egg cylinders were detached from the trophoblast monolayer outgrowth using sterile siliconized Pasteur pipettes and transferred via several DPBS and Tryple Express (Gibco, Cat# 12604) rinse drops, to 15 μL drops of Tryple Express under mineral oil (Sigma, Cat# M8410) in a 60 mm bacteriological plate (Sterilin, Cat# 123) and incubated until the epiblasts had clearly dissociated into small clumps of cells or a single-cell suspension. The disaggregated epiblast cells were then collected in a pipette precharged with serum-containing medium and dispersed into a fresh well of inactivated MEFs at p3-5, in KSR/5×LIF medium. The 4-well plates were left undisturbed in culture for 48 h and thereafter examined daily for the appearance of primary ESC colonies. Typically 4–5 days after picking and disaggregation of epiblasts, feeder wells containing primary ESC colonies could either be dissociated directly to 35 mm tissue culture dishes (Falcon, Cat# 3001) containing feeder cells in 2 mL of KSR/5× LIF, or individual primary colonies could be picked and disaggregated in micro-drops of Tryple Express for clonal cell line expansion via 4-well feeder plates in KSR/5× LIF medium. The ESC lines were subsequently expanded by passaging every 3–4 days in fully supplemented KO DMEM containing either 15% KSR or 15% FBS and with 103 U/mL LIF, with changes of medium as required.
Formation of EBs
Feeder-free ESC were obtained either by passaging twice on tissue culture flasks (Corning, Cat# 430372) coated with 0.1% gelatin (Sigma, Cat# G1890) in DPBS, or by culturing the ESC suspension in a minimal volume of serum containing medium on tissue culture plastic for several 30 min periods to allow feeder cells to attach while leaving the ESCs in suspension. Feeder-free ESCs were then plated at 4×105 cells per 90 mm bacteriological plate (Sterilin) in fully supplemented DMEM containing 15% FBS and cultured for 14 days. For in vitro differentiation studies, EBs were plated onto glass slides (Millipore Millicell EZ slides, Cat# PEZGS0416) and cultured in DMEM+15% FBS before staining with fluorescently labeled monoclonal antibodies (mAbs).
Sex determination of novel ESC lines
Four to 6 EBs from each ESC line were washed in DPBS and transferred in minimal carry-over volume to 50 μL of 200 μg/mL Proteinase K (Sigma) in digestion buffer (50 mM KCl in 10 mM Tris-HCl, pH 8.3, plus 0.1 mg/mL gelatin, 0.45% NP40, and 0.45% Tween 20) in a PCR tube and digested at 55°C for 5 min in a Techne TC-412 thermocycler. Proteinase K enzymatic activity was halted by heating to 95°C for 6 min, followed by a cooling step. A sample of the digested EB preparation (22 μL) was added to an equal volume of master-mix of JumpStart REDTaq Ready Mix PCR 2× Reaction mix (Sigma, Cat# P0982), primers, and autoclaved water. The primers used were Sry (402 bp), IL3 (544 bp) [35], and MyoG (250 bp) [33], the sequences of which are given in Supplementary Table S1; Supplementary Data are available online at
PCR products were separated on a 1.5%–2% agarose/TAE gel containing 0.5 μL/mL ethidium bromide (Sigma, Cat# E1385) at 68V for up to one h, alongside a 100 bp DNA ladder (Thermo Scientific, Cat# SLL-100S-LD). By UV trans-illumination, male ESC lines displayed an SRY band flanked above and below by IL3 and MyoG bands; whereas female lines give only the latter 2 bands.
Karyotyping
Male and female ESC lines that were able to form EBs and which had given unambiguous results for sex determination by PCR were karyotyped by G-banding [30]. Briefly, ESC at day 3 of passage were dissociated to a single-cell suspension as for routine passaging, and half the cell suspension was plated onto fresh MEFs in a 25 cm2 tissue culture flask with 7 mL medium containing LIF and cultured overnight. A 40 μL drop of 10 μg/mL colcemid (Sigma, Cat# D1925) was added, and the cells were incubated for 30 min. The ESC and feeder cells were scraped off and briefly exposed to a hypotonic solution, and then fixed in methanol/acetic acid. Chromosome spreads were prepared, and Giemsa were stained for karyotype analysis. At least 30 G-banded mitotic cells were routinely scored for chromosome number, together with the sex chromosomes and the normality of the autosomes, and the findings were expressed as percent normal cell spreads.
Production of chimeric mice and GLT
Blastocyst injection studies and Mouse Contract Services were provided by the Mary Lyon Centre at MRC Harwell, United Kingdom (
Teratoma formation
The ability of ESC-derived EBs to differentiate into tissues representative of the 3 primary germ layers was determined by unilateral transplantation of 3–4 EBs under the kidney capsule of immune-compromised adult CBA.RAG1−/− mice. EBs were washed in DPBS and maintained on ice before implantation. Teratomas were recovered after 21 days, rinsed in DPBS, and embedded in optimum cutting temperature compound (VWR). Cryosections of 7 μm thickness were air dried, rinsed in DPBS, and either stained with hematoxylin and eosin or fixed with fresh 4% paraformaldehyde in DPBS and immunofluorescence stained.
Hematoxylin (0.5%, Harris') and eosin (1%) stained cryosections were viewed and photographed on a Nikon Coolscan system using ×5, ×10, and ×20 objective lenses. Immunofluorescence antibody stained material was viewed and photographed on an Olympus confocal microscope using Olympus proprietary acquisition software. Images were edited in Adobe Photoshop CS4.
Immunohistochemistry
Cryosections from teratomas were first air dried for 15 min at room temperature. ESC colonies, EBs cultured on glass slides, and cryosections were rinsed in DPBS fixed in 4% paraformaldehyde in DPBS for 20 min at room temperature. After DPBS rinses, slides were permeabilized, if required, in 2 changes of 0.1% Triton-X in DPBS for 15 min at room temperature. Slides were then blocked in 10% normal donkey serum (NDS) in DPBS containing 1 μL/mL mouse Seroblock FcR (AbD Serotec, Cat# BUF041B) for 1–2 h at room temperature. The slides were then incubated in primary mAb diluted in 1% NDS in DPBS according to the manufacturer's instructions. Primary mAb incubations were performed for either 1–2 h at room temperature or overnight at 4°C. Where directly fluorescently conjugated primary antibodies were not used, appropriate secondary antibodies diluted in 1% NDS/DPBS were applied for 1–2 h at room temperature. Slides were coverslip mounted in 4′,6-diamidino-2-phenylindole dihydrochloride containing mountant (Vectashield, Cat# H-1200) and viewed with an Olympus confocal microscope with×20 air or×40 oil immersion lenses.
Differentiation of ESC to DCs
EBs at day 14 were transferred onto 100 mm tissue culture plates (Corning, Cat# 430167) in DMEM supplemented with 2 mM L-glutamine, 1 mM sodium pyruvate, 0.1 mM MEM nonessential amino-acids, 0.2 mM 2-mercaptoethanol, and 15% FBS, further supplemented with 200 U/mL recombinant IL-3 (R&D Systems) and 25 ng/mL of granulocyte-macrophage colony stimulating factor (GM-CSF) [ESC-derived DCs (esDC) medium] [32]. By 14 days of culture, esDCs could be harvested and induced to mature by addition of 1 μg/mL of lipopolysaccharide (LPS) from E. coli Serotype 0127:B8 (Sigma). After 16 h of exposure to LPS, the medium was replaced with fresh esDC medium for a further 24 h, and nonadherent mature esDCs were harvested and stained for flow cytometric analysis.
Flow cytometry
EBs, ESCs, and esDCs were prepared for analysis on a Becton Dickinson FACSCalibur using CellQuest software, as follows. Single-cell suspensions were made when necessary, either using Cell Dissociation buffer (Gibco, Cat# 13150-016) or using Tryple Express followed by a recovery incubation period. Cells were washed with 0.01% EDTA in DPBS and incubated on ice in blocking buffer (5% heat-inactivated normal rabbit serum, 1% BSA, 0.1% sodium azide in DPBS) containing 1 μL/mL Seroblock FcR for 20 min. The cells were then washed in cold 1% FBS, 0.1% sodium azide in DPBS (Wash buffer) and exposed to primary mAbs for 30 min on ice. Antibodies were diluted in Wash buffer. Isotype-matched primary antibodies were used as negative controls. Appropriate secondary antibodies were applied when necessary, for 20–30 min on ice, after first washing the cells. An equivalent volume of 2× 7-Aminoactinomycin D (Sigma, Cat# A9400) in DPBS (8 μg/mL) was applied for the final 10 min of staining. After 3 washes, the cells were fixed in 4% paraformaldehyde in DPBS for 20 min at room temperature or overnight at 4°C, and flow cytometry was carried out as soon as possible. Data were further analyzed using TreeStar Flowjo software. Primary and secondary antibodies used for flow cytometry are shown in Supplementary Table S2.
Mixed leukocyte reaction (MLR)
Immature and LPS-matured esDCs were mitotically in-activated with MMC and co-cultured with nylon-wool purified T cells from allogeneic strains of mice in RPMI 1640 (Lonza, Cat# BE12-702F) supplemented with 2 mM L-glutamine, 50 U/mL penicillin, 50 μg/mL streptomycin, and 0.2 mM 2-mercaptoethanol plus 10% FBS (R10 medium) in 96 well-round bottomed plates (Falcon, Cat# 3077) for 72 h. The co-cultures were pulsed with 0.5 μCi/well 3H-thymidine (Amersham, Cat# TR120) for a further 16 h, harvested, and levels of 3H incorporation were assessed using a flat-bed scintillation counter.
Statistical analysis
χ 2-analysis was applied to Tables 1 and 2 using 4 degrees of freedom.
Two of the 3 epiblasts were disaggregated into a single well, therefore unknown whether one or both contributed to primary colonies.
Chi Square gives P=0.402 or 0.249 (for “Number of epiblasts generating ESC lines”=2 or=3), d.o.f.=4.
ESC, embryonic stem cell; LIF, leukemia inhibitory factor; FBS, fetal bovine serum; d.o.f., degrees of freedom.
For “Number epiblasts picked” v “Number epiblasts giving ESC lines”:-
Chi Square=18.4.
d.o.f.=4.
P=0.001.
KSR, knockout serum replacement.
Results
The use of high concentrations of LIF, in conjunction with KSR, increases efficiency of ESC derivation
Since C57Bl/6 and Balb/c ESC lines had previously been established using 5×103 U/mL LIF [34], we investigated whether similar conditions might prove successful for the related strain, Balb/K. Three-and-a-half-day blastocysts from Balb/K mice were, therefore, cultured on fresh MEF feeder cells in fully supplemented KO DMEM, containing either 15% FBS or 15% KSR. The concentration of LIF was varied so that embryos were exposed to either 1×103 U/mL (1×) or 5×103 U/mL (5×). RESGRO culture medium (Millipore), which has been reported to improve ESC derivation for the related Balb/c mouse strain, was also included for comparison. Table 1 shows that, although the sample size is too small to reach statistical significance (P>0.249), medium containing 15% KSR with 5×103 U/mL LIF (KSR/5× LIF) showed greater efficacy, both in terms of the proportion of embryos that yielded ESC lines and the number of lines generated from individual embryos.
Given these preliminary results, we next investigated whether the same conditions might permit the derivation of ESCs from the CBA/Ca mouse, a strain regarded as one of the most refractory strains. The range of culture conditions examined was extended to include FBS in conjunction with LIF concentrations of 1×103 U/mL (FBS/LIF) or 5×103 U/mL (FBS/5× LIF), and KSR with LIF at 1×103 U/mL (KSR/LIF), 2.5×103 U/mL (KSR/2.5× LIF) or 5×103 U/mL (KSR/5× LIF). The results in Table 2 strongly indicate that the combination of KSR and 5× LIF was, once again, the most efficient protocol for ESC derivation (P=0.001). Further, other strains, such as C57Bl/6, including several knockout strains on a C57Bl/6 genetic background, could also be derived at substantially higher efficiency using KSR/5× LIF than have been previously reported using standard FBS/LIF (Table 3). Further, the same culture conditions proved applicable to either freshly isolated or cryo-preserved mouse embryos. Most importantly, however, using these conditions, we were able to generate novel ESC lines from cryo-preserved mouse embryos of the NOD/ShiLtJ strain, which has historically proved to be the most nonpermissive strain for ESC derivation by standard techniques, earlier requiring the use of small molecule inhibitors (2i) for efficient derivation [14]. NOD ESC colonies, derived under these conditions, displayed typical ESC morphology (Fig. 1A) and demonstrated expression of the pluripotency genes Nanog, Oct3/4, and SSEA-1 by immunofluorescence staining and flow cytometry (Fig. 1B, C).
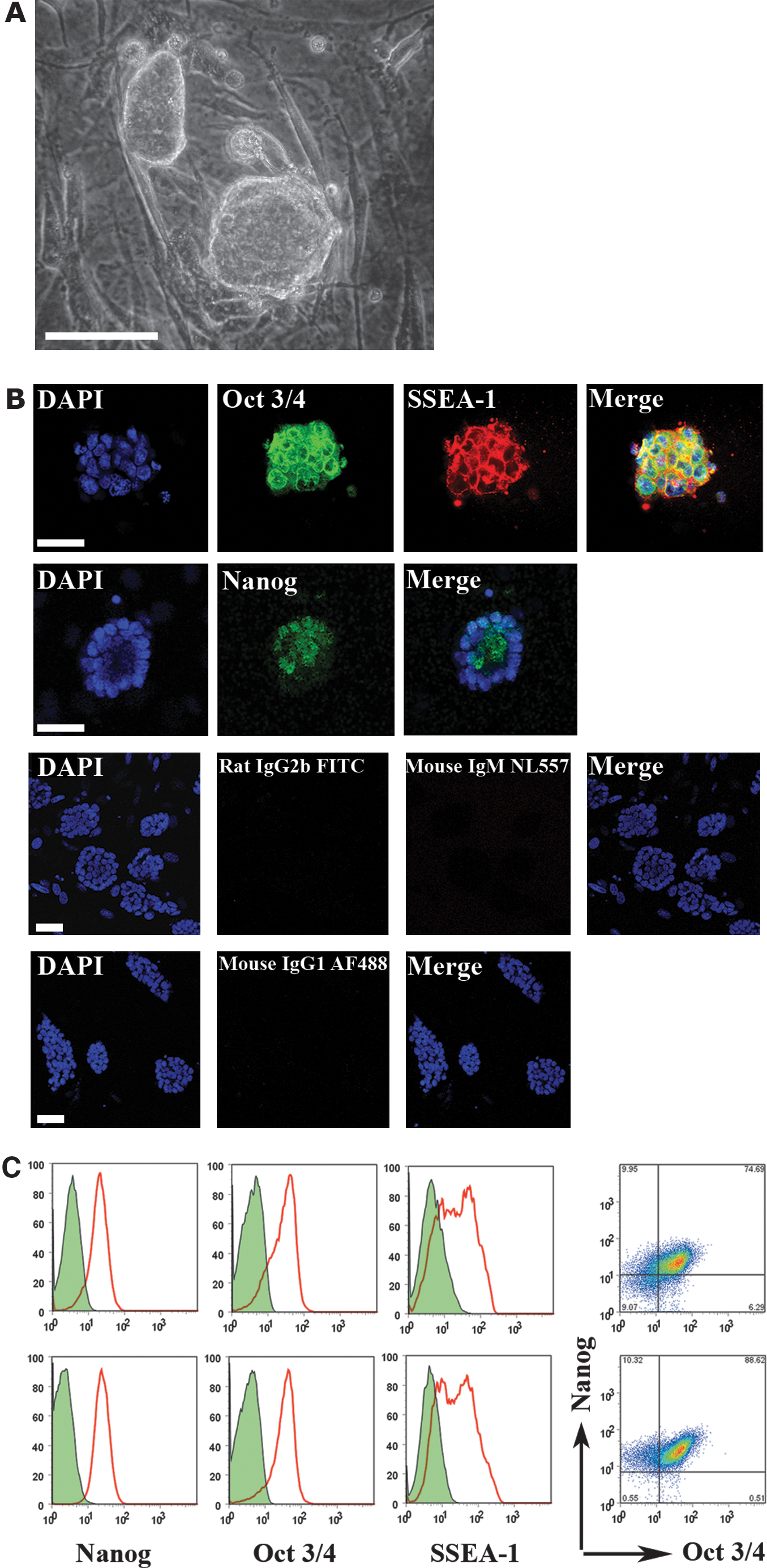
Novel ESC lines express pluripotency genes.
Two epiblasts dissociated into the same well.
NOD, non obese diabetic.
Karyotypic stability of ESC lines
Novel ESC lines, generated using KSR/5× LIF, were sexed by means of multiplex PCR amplification of the male-specific Sry gene sequence on the Y chromosome [35]. Primers for the autosomal genes IL3 (chromosome 11) and Myogenin (Myo G, chromosome 1) were used as controls to ensure the proper amplification of the genomic DNA template (Fig. 2A). The results in Table 4 show that it was possible to derive both male and female ESC lines from each strain using our protocols, with the exception of the Bcat-1−/− mutant strain on a C57Bl/6 background.
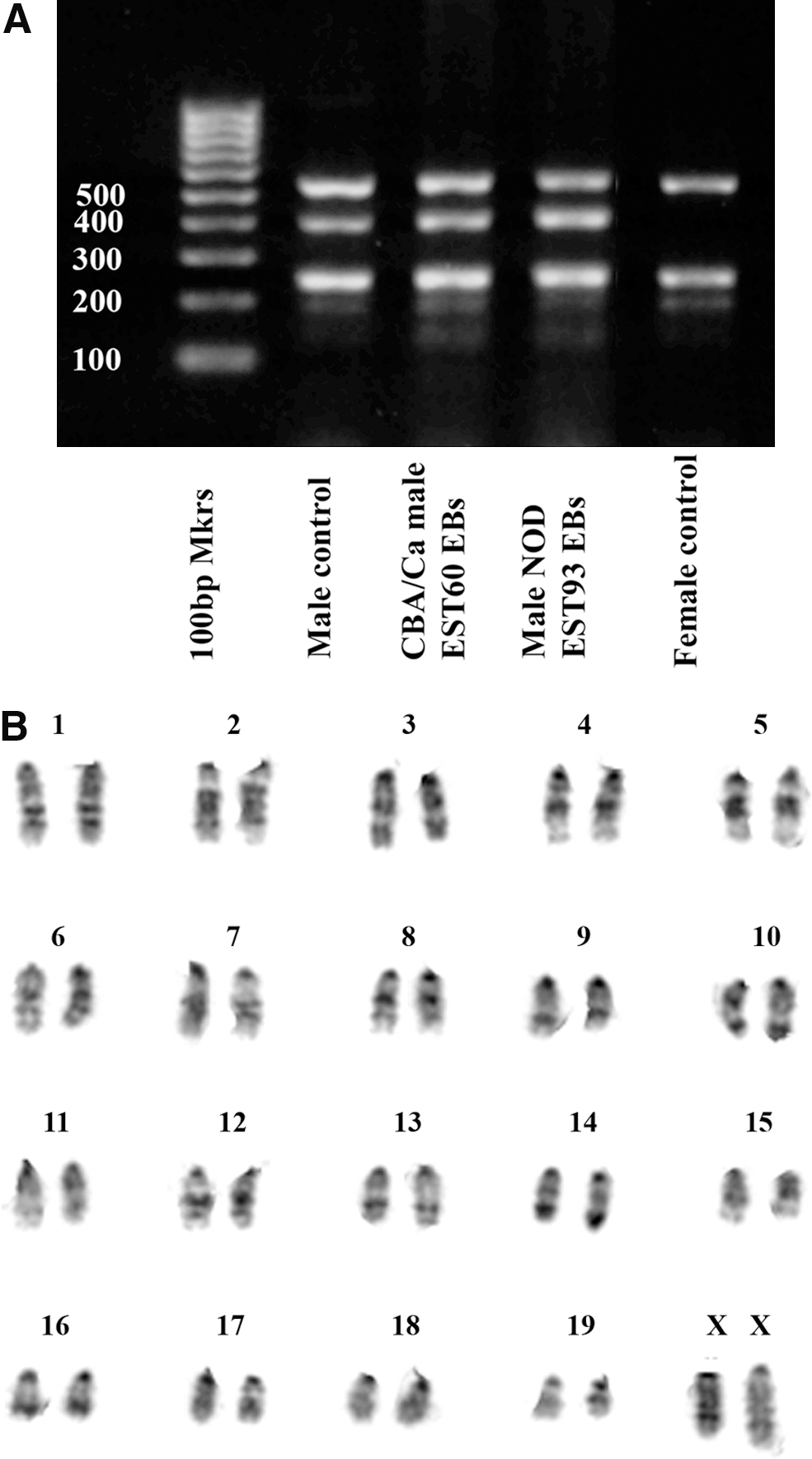
ESC lines derived in knockout serum replacement/5× LIF have a robustly stable karyotype.
Not all embryos giving ESC lines were typed by PCR for SRY. There was no significant male:female bias among those embryos tested.
For 2× Balb/K males, Chi Square=9.09; d.o.f.=6, P=0.168.
For 3× Balb/K males, Chi Square=9.83; d.o.f.=6, P=0.132.
Most novel ESC lines tested proved to be stable in culture to high passage number, requiring only standard ESC culture conditions from passage 2 onward. Of the 21 male ESC lines and sub-lines karyotyped by G-banding at p7–p12, 19 had a normal diploid karyotype, including the NOD ESC line, EST93. Interestingly, of the 7 female ESC lines karyotyped, 5 were 39 XO, but the 2 female NOD sub-lines were found to be 40XX. Untypically for female ESCs, which invariably lose an X chromosome during early culture, the female NOD ESC line, EST94, retained both X-chromosomes until late passage (p17), while maintaining a robustly normal karyotype (Fig. 2B, Supplementary Table S3). Even at p22, there was some retention of both X chromosomes (13%) by the NOD sub-line EST95.
Spontaneous differentiation of novel ESC lines
To investigate the capacity for differentiation of ESC lines derived in KSR/5× LIF, we selected a representative CBA/Ca line, EST60, and both male and female NOD lines (EST93 and EST94, respectively) and assessed their ability to differentiate both in vitro and in vivo. NOD ESC lines derived earlier under 2i conditions showed impaired capacity for differentiation in vitro, as evidenced by their failure to form EBs [29], whereas both EST93 and EST94 consistently formed EBs in suspension over the same time frame as the control CBA/Ca line, EST60. EBs cultured on either glass slides or tissue culture plastic produced cell types representative of each of the 3 primary germ layers. Tissues of endoderm origin were morphologically distinguishable by their ‘cobblestone’ appearance and the presence of glandular epithelial structures (Fig. 3A, arrows), ectoderm derivatives by neuronal precursor cells (Fig. 3B, arrows) and mesoderm by red blood cells, vascular structures, and beating cardiomyocytes (Fig. 3C, D and Supplementary Movie;). Lineage commitment was confirmed by immunofluorescence staining using antibodies specific for β-catenin (endoderm), nestin (ectoderm), and smooth muscle actin (SMA) (mesoderm) (Supplementary Fig. S1).
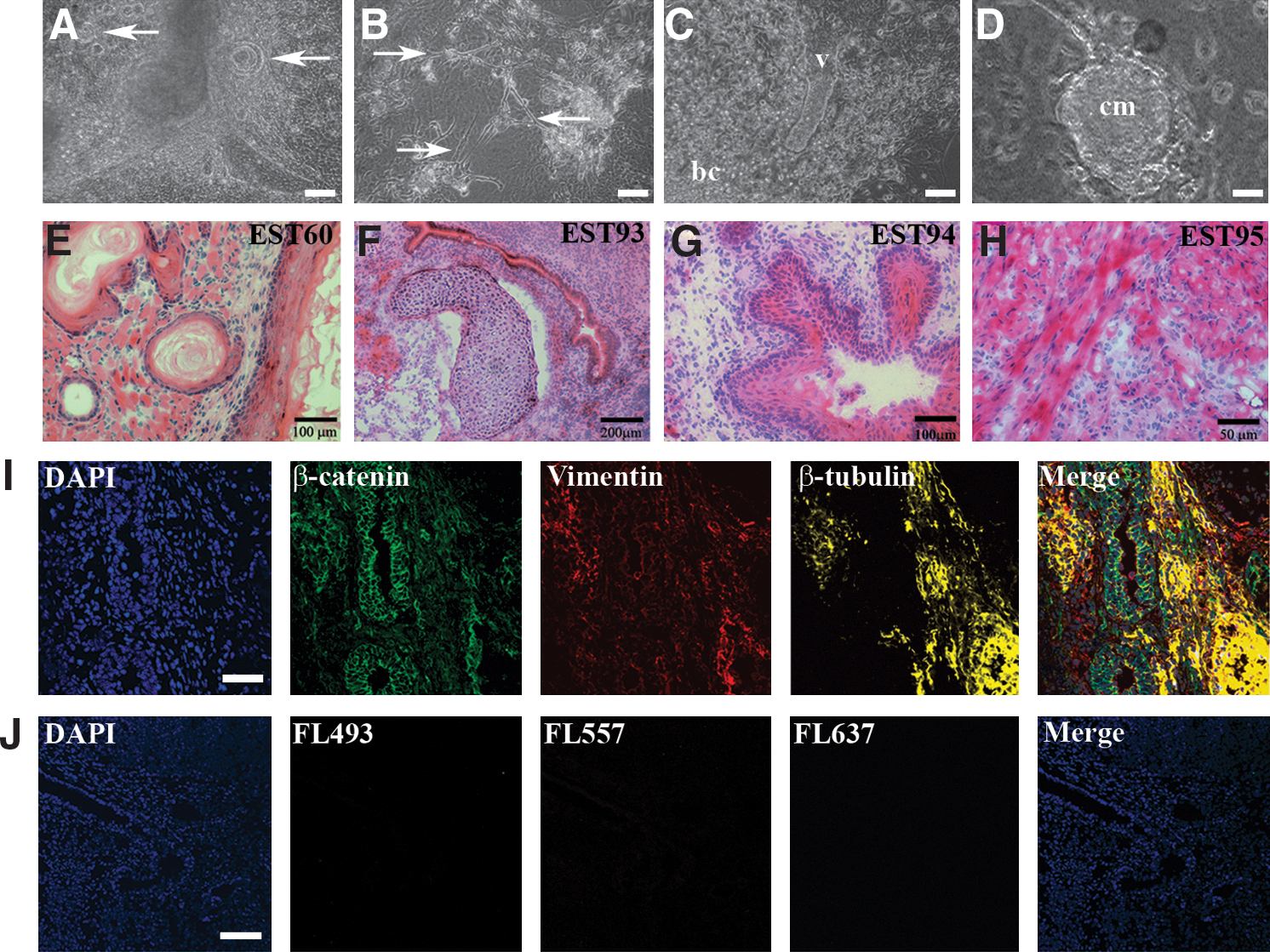
In vitro and in vivo differentiation of ESC lines into derivatives of the 3 primary germ layers.
The ability of these same ESC lines to differentiate in vivo was examined by transplantation of 14 day EBs under the kidney capsule of immune-compromised CBA.RAG1−/− mice (Table 5). Each of the CBA/Ca and NOD lines readily formed teratomas that were harvested and sectioned 21 days later. Hematoxylin and eosin staining of cryosections revealed a variety of disordered tissues and anatomical structures originating from each of the 3 germ layers (Fig. 3E–H). Immunofluorescence staining confirmed the presence of tissues derived from endoderm (as defined by staining for β-catenin), ectoderm (βIII-tubulin and nestin), and mesoderm (SMA, desmin, and vimentin) (Fig. 3I, J and Supplementary Fig. S1).
Contribution of CBA/Ca and NOD ESCs to the formation of chimeric mice
We next investigated the capacity of ESC lines generated using KSR/5× LIF to contribute to the formation of chimeras, capable of GLT. Accordingly, when the male CBA/Ca ESC line, EST60, was injected into C57Bl/6 host blastocysts and transferred to pseudopregnant recipients, it was able to generate chimeric offspring, as judged by coat color (Table 6, Fig. 4). Two male chimeras with high ESC contribution (40%, 80%) from a litter of 8 pups arising from injections of EST60 were highly germline competent, yielding 87% and 100% agouti offspring respectively, when crossed with C57Bl/6 females. Importantly, both the male (EST93) and female (EST94) NOD ESC lines also gave rise to chimeras of either sex. The male EST94 chimera displayed a 10% contribution from the NOD ESCs, as judged by coat color; whereas the female chimera showed up to 70% NOD ESC contribution. In one set of injections, the EST93 cell line gave rise to 6 chimeras (24% of offspring), 3 of either sex, which showed significant variation in the extent of chimerism from 5% to 70% (Fig. 4).
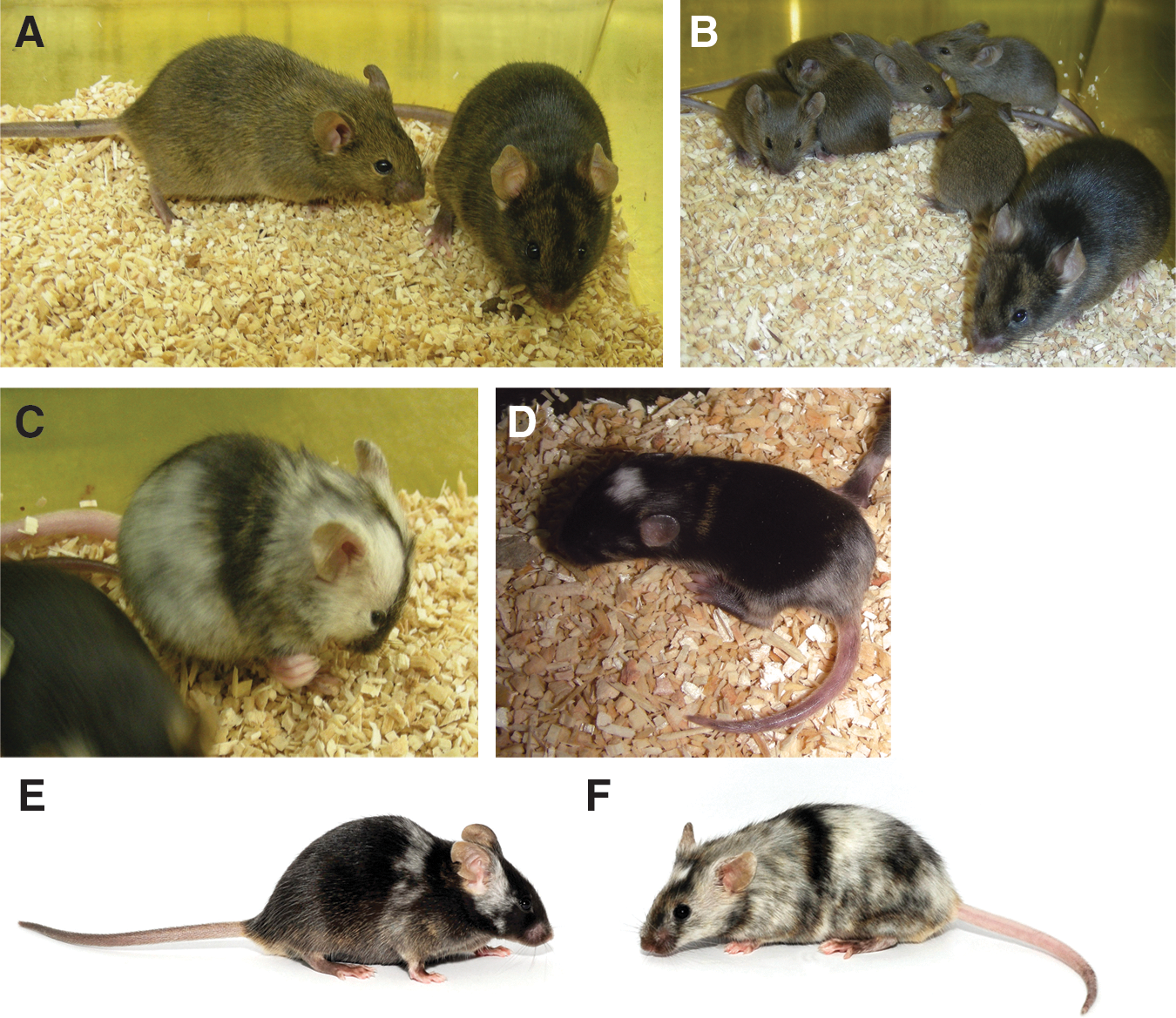
Novel ESC lines generate chimeras and germline competent offspring.
One litter of 7 pups was killed by the mother before it was possible to assess chimerism.
ND, not determined.
Directed differentiation
Given that NOD ESCs derived with KSR/5× LIF not only contributed to chimeras but readily formed EBs in suspension culture, we investigated their capacity for directed differentiation in vitro into cell types, such as DC, implicated in the pathogenesis of type I diabetes. EBs cultured for 14 days in suspension, were, therefore, plated onto tissue culture plastic in complete esDC medium, containing GM-CSF and IL-3, for a further 14–21 days, during which DC accumulated to significant numbers around and between EBs (Fig. 5A), as previously described for other strains [32]. Commitment to the DC lineage was confirmed by flow cytometric analysis of their surface phenotype. NOD esDCs derived from the male line, EST93, expressed CD11c, and, after maturation in response to the TLR4 agonist LPS, showed up-regulation of the co-stimulatory molecules CD54 and CD86 (Fig. 4B). In addition, NOD esDCs strongly up-regulated major histocompatibility complex (MHC) class II determinants of the H-2Ag7 haplotype, unique to this disease-prone strain of mouse, and assumed a highly dendritic morphology (Fig. 5A). Further, in accordance with this immunostimulatory phenotype, mature NOD esDC stimulated potent proliferative responses among naïve allogeneic CBA/Ca T cells, compared with their immature counterparts (Fig. 5C).
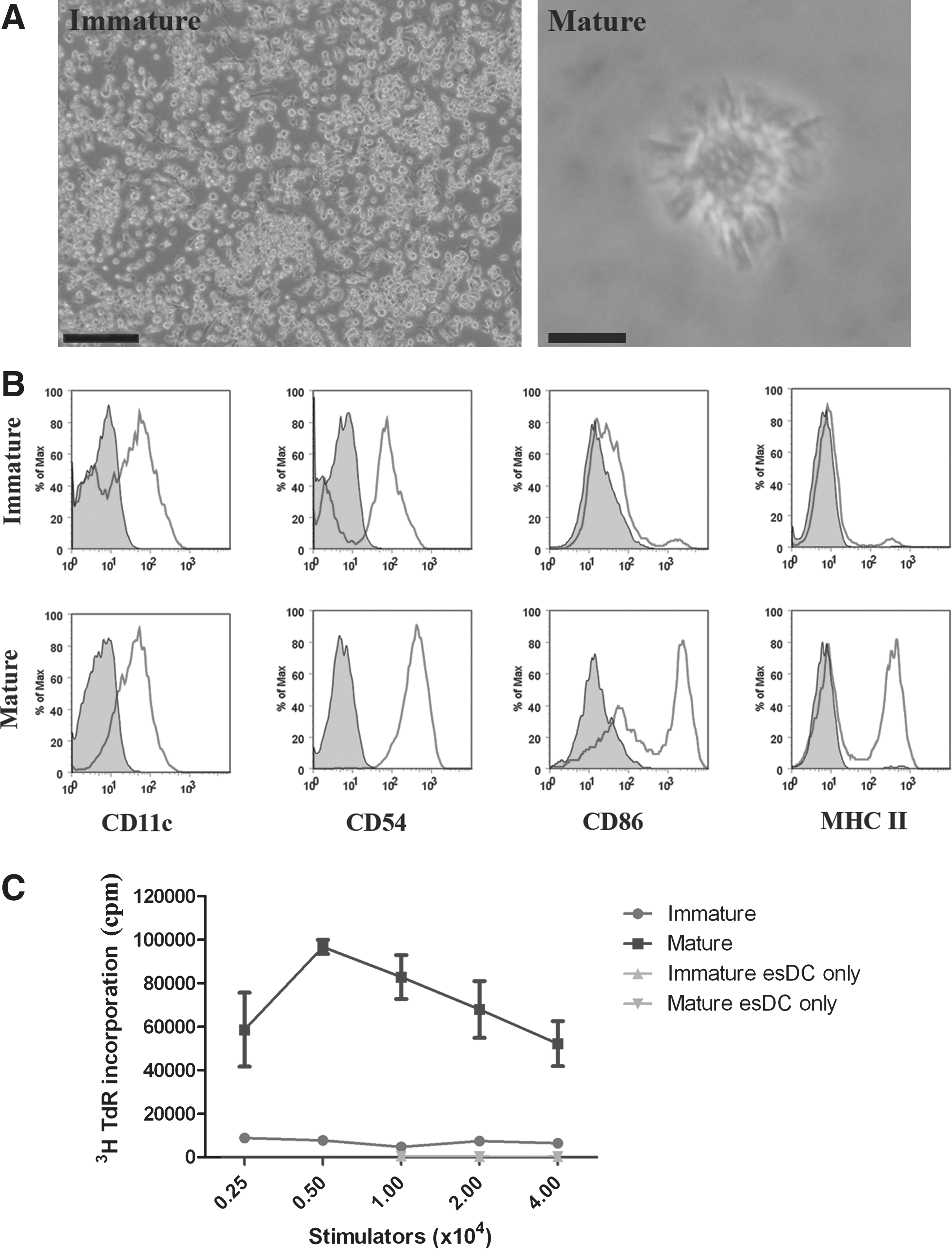
Directed differentiation of NOD ESCs into functional DCs.
Discussion
We describe here a simplified method for the efficient generation of ESC lines using a combination of KSR and high LIF concentration, which can be applied to even the most refractory mouse strains. Novel ESC lines, produced using this protocol, have predominantly normal and stable karyotypes; express the pluriportency-associated genes Nanog, Oct3/4, and SSEA-1; are capable of forming teratomas that display derivatives of each of the 3 primary germ layers; and can form germline competent chimeras when injected into host blastocysts. Although KSR/5× LIF is essential for the initial stages of ESC derivation, only small volumes of this medium are required (typically between 1.5 and 3 mL per embryo) without the requirement for additional reagents, such as growth factors or small molecule inhibitors. Subsequent culture of novel ESC lines can be performed under standard conditions, thus making this protocol both efficient and cost effective.
The relatively permissive C57Bl/6 strain served as a viability control for the refractory NOD mouse, both being obtained as cryo-preserved embryos from the same commercial source and derived in parallel under identical conditions. It is difficult to compare efficiencies of derivation between different published studies because of the inevitable inconsistencies in criteria used to assess the data. The rates for successful derivation of ESC lines presented here (Tables 2 and 3) are based on the proportion of epiblasts that were suitable for picking after a period of blastocyst outgrowth on feeder cells. Only 4 of 119 (3.4%) freshly recovered CBA/Ca blastocysts plated were discarded as being unsuitable for epiblast picking (Table 2), whereas 4 of 19 (21%) cryo-preserved C57Bl/6 blastocyst outgrowths were considered nonviable. ESC derivation was attempted from all 12 similarly cryo-preserved NOD blastocyst outgrowths, irrespective of their morphology. It is possible that the efficacy of derivation for C57Bl/6 (80%) and NOD (17%) ESCs may have been greater if freshly recovered embryos had been used. Notwithstanding the use of frozen embryos, our figure of 80% efficiency of C57Bl/6 ESC generation compares very favorably with previous studies using KSR in conjunction with 1× LIF on feeder cells, which report rates of 19%–36% [10] and 50–75% [16]. The latter study employed alternating steps of culture in medium containing either KSR or FBS, and there is some evidence that the presence of a small quantity of FBS (0.3%) in conjunction with KSR may be beneficial to ESC maintenance [36]. The use of KSR in ESC derivation is further supported by the experience of our laboratory and of others [17] that KSR is preferable to FBS for the derivation of iPSCs from mouse adult and embryonic fibroblasts.
Effect of background strain on ESC derivation
The reasons that some mouse strains lend themselves to the derivation of ESCs more than others have yet to be fully explained. There is evidence that the inhibitory effect of the hypoblast on epiblast proliferation may vary between strains. However, in a study in which 129 ESCs could be derived in N2B27 medium supplemented with LIF and BMP4, removal of the hypoblast of delayed implanted embryos was not sufficient to permit ESC derivation from C57Bl/6 and CBA/Ca epiblasts, unless Erk activity was also chemically suppressed [37]. These findings are in contrast to our results in which we observed efficient derivation of both C57Bl/6 (80%) and CBA/Ca (56%) ESCs, without recourse to elaborate embryo manipulation or the use of chemical inhibitors.
Differences between NOD and C57Bl/6 embryo outgrowth were evident by the time of epiblast picking, the NOD forming relatively large egg cylinders with a grainy opaque cytoplasm, compared with C57Bl/6 outgrowths. Picked and dissociated NOD epiblasts yielded between 1 and 5 primary colonies (mean=2.8+1.6 S.D., n=5); whereas the smaller C57Bl/6 epiblasts gave between 1 and 22 primary colonies (mean=10.75+7.5 S.D. n=12). For C57Bl/6, 80% of picked epiblasts gave primary colonies, compared with 42% of picked NOD epiblasts. We were able to obtain secondary colonies and to derive ESC lines from all C57Bl/6 primary colonies, whereas ESC lines could only be derived from 2 of the 5 NOD epiblasts that yielded colonies. NOD sub-lines, which failed to expand, were generally lost after passage 2. During routine culture, NOD ESCs were generally slower to reach confluency after cell dissociation than ESCs from other strains and benefited from being plated at a higher density during passaging.
KSR permits derivation of ESCs from refractory strains
Only 1 out of 2 blastocysts from the refractory CBA/Ca strain initially plated into KSR/5× LIF, whose epiblasts were picked and dissociated into FBS/5× LIF, subsequently gave primary colonies. Primary colonies from this embryo, when disaggregated to FBS/5× LIF, failed to yield an ESC line, thus suggesting that exposure to 15% FBS at the primary colony stage may be detrimental to ESC derivation. The beneficial effect of the biochemically defined serum-free KSR in ESC derivation may be due to the elimination of extraneous factors found in FBS, which may either induce differentiation or inhibit self-renewal. In addition, ESCs cultured in medium with KSR exhibit more pronounced dome-shape colonies than those cultured in medium containing FBS, which tends to encourage colonies that are initially flatter but subsequently become dome-like [38]. This may provide the ESC with a more advantageous microenvironment for ESC proliferation. Most ESC lines derived here could be cultured under feeder-free conditions while routinely displaying a stem-cell morphology, expressing pluripotency markers (Fig. 1), and retaining the ability to fully differentiate on withdrawal of LIF (Fig. 3).
High concentrations of LIF are required for the improved generation of ESC
For the relatively nonpermissive CBA/Ca strain, the replacement of FBS with KSR was an absolute requirement for ESC derivation (Table 2). Increasing the concentration of LIF, in conjunction with 15% KSR, further improved the efficiency of ESC generation (P=0.001). We also observed that the number and size of early passage colonies were greater with elevated LIF levels, thus suggesting increased proliferation. Earlier, studies that have succeeded in deriving CBA/Ca ESCs have required the influence of the hypoblast to be suppressed after immunosurgery to remove trophoblast cells, either by sophisticated microsurgical means [15] or by inhibition of the ERK pathway using chemical inhibitors [37]. Interestingly, we did not find it necessary to induce preimplantation developmental diapause, immunosurgically remove the trophoblast, or isolate the epiblast by microsurgery or chemical inhibition, to derive CBA/Ca ESCs using our protocol. Adopting these modifications in conjunction with the method presented here may further improve efficiency of derivation. That variation of genetic background accounts for differences in the response to LIF in ESC derivation is suggested by studies in which C57Bl/6 ESCs could be derived in 20% KSR using 103 U/mL LIF; whereas, under the same conditions, Balb/c ESC derivation required 20% KSR and 5×103 U/mL LIF. Nevertheless, increased LIF was not effective in generation of NZB ESCs [28]. A facet of our protocol that may be important in successful generation of ESCs from nonpermissive strains is the presence of LIF during the epiblast expansion stage of embryo culture. LIF maintains pluripotency in ESCs by activation of the STAT3 pathway [20], and, when added to the culture medium of zygotes cultured to the blastocyst stage, has been demonstrated to increase blastocyst total cell number through increased proliferation of the ICM, thus leading to enhanced ESC derivation [39]. By culturing preimplantation embryos through to the equivalent of the early egg cylinder stage on feeder cells in KSR/5× LIF, the ESC progenitor cell population of the epiblast is expanded. Epiblast picking at the appropriate time and, if required, primary colony picking and dissociation to fresh feeder cells in KSR/5× LIF serves to remove both trophectoderm and primitive endoderm from the expanded stem cell pool.
Novel ESCs display a stable karyotype
We have shown that NOD ESCs, derived in KSR/5×LIF, retain a normal diploid karyotype until high passage number. The male NOD ESC line (EST93) was 100% 40XY at p11 and 97% 40XY at p22; whereas the female NOD line (EST94) retained both X chromosomes in all cells scored at p17 (Supplementary Table S3). Under standard FBS/1× LIF conditions, novel female ESC lines typically lose an X chromosome during early passage (p4-p6), and the majority of cells are 39XO by p15, with increasing incidence of trisomy 8 [30]. However, with KSR/5× LIF, a sub-line of the female NOD epiblast (EST95) retained a normal 40XX karyotype in 13% of cells, even at p22, the remainder being predominantly 39XO. That both NOD embryos yielded chromosomally stable lines may reflect the fact that they were generated without chemical perturbation. In the only previous study to date that has efficiently generated germline-competent NOD ESC lines, 5 of the 11 lines (45%) whose metaphase spreads were examined were aneuploid [14]. The same study reports that 16 NOD ESC lines were derived from 30 embryos using PD0325901, a small-molecule inhibitor of MEK, and the GSK3 inhibitor, Chir99021 (collectively 2i), in conjunction with LIF in a serum- and feeder-free system. Three of 4 karyotypically normal lines injected into C57Bl/6 host blastocysts gave chimeras capable of GLT. The speculation that although small-molecule inhibitors may increase the efficiency of derivation they may also have adverse chromosomal effects on the ESCs generated through their use, is supported by observations that aneuploid cells were present in relatively high numbers in NZB ESC lines produced on feeder cells in 3i medium, which contains fibroblast growth factor receptor inhibitor as well as GSK3 and MEK inhibitors [28]. Using KSR/5× LIF, we were able to obtain extremely high rates of GLT from CBA/Ca chimeras (Tables 2 and 3). Several reports have observed improved GLT using C57Bl/6 recipient embryos, either by injecting ESCs into the 8-cell embryo rather than the blastocyst [39 –41], or by co-culture of ESCs with C57Bl/6 8-cell blastomeres [42], followed by embryo transfer. The slower proliferation of NOD ESCs compared with host ICM cells may disadvantage NOD ESCs when injected into blastocysts of a differing strain. Using a genetically marked NOD ESC line that had earlier given very low GLT (<1%) [5], Arai and colleagues reported improved transmission when NOD recipient blastocysts were used [6].
Directed differentiation of novel ESC lines
Other NOD ESC lines, derived under 3i conditions, failed to form EBs [29], or required continuous presence of small molecule inhibitors (or ectopic expression of the transcription factors Klf4 or c-Myc) to retain their pluripotent state [43]. Importantly, we did not observe these difficulties, as almost all our novel KSR/5× LIF-derived ESC lines were stable under standard culture conditions, and were able to form EBs capable of directed differentiation in vitro. Indeed, using protocols first developed by our laboratory [32], esDCs were differentiated from each of the strains of ESCs generated, as well as NOD ESCs for the first time. The NOD mouse is an important model of human type I diabetes and has been widely exploited to better understand the pathogenesis of the disease and develop approaches to its effective treatment. For example, the role played by DCs in the initiation of autoimmunity is now well established, and their exploitation for the re-establishment of immunological tolerance has been demonstrated [44]. Here, we show that NOD ESCs can be directed to differentiate into immature DCs, capable of maturation in response to TLR ligands such as LPS. This novel population of DCs up-regulates conventional co-stimulatory molecules on maturation, together with MHC class II molecules of the g7 haplotype, unique to NOD mice, and stimulate potent primary T cell responses in vitro. These findings suggest that the novel NOD ESC lines generated here will prove a useful resource for the study of type I diabetes, through both the generation of genetically modified strains of NOD mice and the directed differentiation of cell types in vitro, implicated in disease pathogenesis and its prevention.
Footnotes
Acknowledgments
The authors would like to thank Dr. Edward P. Evans for karyotyping of ESC lines, Dr. Jon Godwin for advice on thawing embryos, Mr. Richard Stillion for help with histology, and Dr. Lydia Taboul and Ms. Dee Lynch for chimera production. They are grateful to Ms. Laura Barker, Dr. Kate Silk, Ms. Naoki Ichiryu, and Mr. Simon Hackett for helpful discussion and encouragement. This work was funded by grant G0802538 from the Medical Research Council U.K.
Author Disclosure Statement
The authors declare no financial conflicts of interest. Since submission of our manuscript, the NOD ESC line EST93 has shown the capacity for germline transmission.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.