
Abstract
Activin/Nodal signaling is required for maintaining pluripotency and self-renewal of mouse epiblast stem cells and human embryonic stem cells (hESC). In this study, we investigated whether this signaling mechanism is also operative in cultured epiblasts derived from Days 10.5–12 pig embryos. Pig epiblast stem cell lines (pEpiSC) were established on mouse feeder layers and medium supplemented with basic fibroblast growth factor (bFGF). pEpiSC express the core pluripotency factors OCT4 (or POU5F1), NANOG, SOX2, and NODAL, but they do not express REX1 or alkaline phosphatase activity. Blocking leukemia inhibitory factor (LIF)/JAK/STAT3 pathway by adding the specific JAK I inhibitor 420099 and an anti-LIF antibody over 3 passages did not affect pluripotency of pEpiSC. In contrast, cells grown with the Alk-5 inhibitor SB431542, which blocks Activin/Nodal pathway, differentiated readily toward the neural lineage. pEpiSC are pluripotent, as established by their differentiation potential to ectoderm, mesoderm, and endoderm. These cells can be induced to differentiate toward trophectoderm and to germ cell precursors in response to bone morphogenetic protein 4 (BMP-4). In conclusion, our study demonstrates that pig epiblasts express the core pluripotency genes and that the capacity for maintaining self-renewal in pEpiSC depends on Activin/Nodal signaling. This study provides further evidence that maintenance of pluripotency via Activin/Nodal signal is conserved in mammals.
Introduction
M
Here we tested this hypothesis in pigs, a representative of ungulates. Attempts to derive stem cells from pig embryos have traditionally relied in the isolation and culture of ICM using protocols established for mESC [13, 14]. These strategies have not resulted in consistent results, suggesting that the blastocyst stage may not be amenable for stem cell derivation in this species. We show that pig epiblasts, however, can be isolated and cultured in vitro and that maintenance of the undifferentiated state requires bFGF and Activin A signaling instead of LIF and BMP-4. Pig epiblast stem cell lines (pEpiSC) can be cultured for extensive periods and upon growth factors withdrawal the cells can be induced to differentiate to 3 somatic germ layers, germ cell progenitors, and trophoblast. Our study supports the hypothesis that FGF/Activin/Nodal is a conserved signaling pathway for maintenance of pluripotency in mammals, and this can be exploited for developing strategies for the derivation of ESC from ungulate embryos.
Materials and Methods
Epiblast isolation, stem cell cultures, and in vitro differentiation
Pig epiblasts were isolated from in vivo-derived ovoid-early tubular stage embryos collected 10.5–12 days after insemination. Embryo development can vary greatly between individual animals; therefore, embryos showing advanced signs of development were not included in the isolations. The epiblasts used in our study correspond to late ICM-Pre-streak I/II stages, as described by Vejlsted et al. [15]. Embryos were flushed from uterine horns with phosphate-buffered saline (PBS) containing 5% fetal calf serum (FCS), and transferred to culture medium supplemented with 25 mM HEPES. Epiblasts were separated from trophoblast and underlying primitive endoderm (PE) using 21 G needles and fine forceps. Pure epiblasts were placed individually onto 4-well dishes containing mitomycin C-inactivated mouse feeder cells (50×104 cells/cm2) and were cultured in 5% CO2 at 39°C. Mouse fibroblasts were derived from E13.5 fetuses from CD1 strain, and were passaged 2–3 times before mitomycin C inactivation. Feeder layers were used 2–3 days after mitomycin C inactivation. Stem cell culture medium (SCM) consisted of Dulbecco's modified Eagle's medium (DMEM)/F12 containing 1% minimal essential medium (MEM) nonessential amino acids, 1% glutamine, 1% penicillin/streptomycin, and 0.007% β-mercaptoethanol supplemented with bFGF (R&D Systems, Minneapolis, MN) and either serum replacement (SR; Invitrogen, Carlsbad, CA) or FCS (see Table 1 for details). Plating efficiency of primary cultures was determined by scoring the number of attached colonies after 48 h. Attached epiblasts were grown initially for 5–7 days before manual splitting and transfer onto fresh feeders. These colonies were passaged another time before freezing by vitrification as described by Reubinoff et al. [16]. Survival of vitrified colonies was around 90% for 10 lines tested. Mouse LIF (1,000 IU/mL; Gibco, Grand Island, NY) was used for initial isolations. Conditioned medium was prepared from primary mouse embryonic fibroblast inactivated with mitomycin C as described by Xu et al. [17]. Medium was changed daily and cells were passaged manually every 5–6 days. When enzymatic passaging was used, a mixture of 1 mg/mL collagenase type IV and 1 mg/mL dispase (Gibco) was added to the cultures for 15 min at 39°C. Colonies were disrupted by gentle pipetting and transferred into 15-mL tubes containing fresh culture medium and left for 5 min to sediment. The medium was removed and 5 mL of fresh medium was added before washing by centrifugation. Cell pellets were resuspended by gentle pipetting and distributed onto fresh feeders. mESC were grown as described by Bian et al. [18]. For spontaneous differentiation, pEpiSC at 50%–70% confluency were cultured in DMEM/F12 + 10% FCS without bFGF for 30 days. For neural differentiation, we used the protocol described by Conti et al. [19]. For differentiation to TE and to germ cell precursors, EpiSC at Passages 6–9 were manually cut into small pieces and aggregated into embryoid bodies (EB) using hanging drops for 2–3 days. EB were transferred to low attachment dishes and kept in suspension for another 4 days, after which they were transferred to gelatinized dishes with SCM containing 10% FCS and supplemented with 10–50 ng/mL (for trophoblast; for EpiSC8 and 21) and 50–100 ng/mL (for germ cell precursors; for EpiSC8, 21, and 42) of BMP-4 (R&D Systems). This medium was changed daily for up to 14 days.
Indicates lines propagated for >12 passages.
Immunocytochemistry and karyotype
Cells grown on coverslips were washed in PBS before fixation with 4% paraformaldehyde for 20 min. After washing, the cells were permeabilized with 0.1% Triton and blocked in PBS + 5% bovine serum albumin (BSA) for 1 h. All primary antibody incubations were carried out overnight at 4°C, unless otherwise indicated. Antibodies used were: Oct4 (Santa Cruz Biotechnology, Santa Cruz, CA; 1:100), Nanog (AbCam, Cambridge, MA; 1:100), Tubulin (R&D Systems; 1:50), and MAP2 (R&D Systems; 1:300). Respective secondary antibodies were added for 1 h at room temperature. After washing with PBS + 1% BSA, the cells were mounted using Vectashield with 4′-6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA). For SSEA-1 (Hybridoma bank, Iowa City, IA; 1:50) and SSEA-4 (Hybridoma bank; 1:50), cells were treated the same except that they were not permeabilized. Fluorescent images were acquired using epifluorescence and simple PCI image analysis software. Alkaline phosphatase (AP) activity was detected using an AP detection kit (Sigma, St. Louis, MO). For karyotyping, cells were incubated in 0.05 ug/mL demecolcine for 30 min, trypsinized, and pelleted. The pellet was resuspended in a hypotonic solution (0.075 M KCl) for 30 min. After centrifugation, the cells were fixed in ice-cold ethanol for 1 h. The cell suspension was dropped onto humidified slides and air-dried. Following dehydration with 70%, 90%, and 100% ethanol, the metaphase spreads were stained with Giemsa. Metaphase spreads (20 per cell line) prepared from 3 cell lines at Passage 7 (EpiSC8, 21) and Passage 12 (EpiSC42) were visualized under phase contrast using 100× objective under immersion oil.
Signaling inhibitors
The STAT3 inhibitor 420099 (Calbiochem, San Diego, CA) was dissolved in dimethyl sulfoxide (DMSO) as 10 mM stock and was added to cells at 0.6 μM as described by Tesar et al. [6]. SB431542 (Tocris, Ellisville, MO) was dissolved in DMSO to 100 mM and stored at −20°C. Cells were treated with 20 μM SB431542 for 6 days as described previously by Tesar et al. [6]. Anti-mLIF antibody was added to the cultures (100 ng/mL; R&D Systems) for 3 passages. Control groups were exposed to an equivalent concentration of DMSO. Plating efficiency was estimated according to Tesar et al. [6] in triplicate experiments. In brief, cell clumps were randomly placed onto 12-well plates and their numbers established after attachment. When colonies reached confluency, they were passaged using collage-nase/dispase and all cells were plated onto new wells. After 3 passages, the total number of colonies was counted for each group and they were represented as percentage difference to the starting number of colonies plated.
Gene expression
RNA isolation was carried out using the RNAeasy kit (Qiagen, Valencia, CA) following the manufacturer's instructions. Dissected epiblasts, trophoblast, and pEpiSC colonies were frozen in RLT buffer containing p-mercaptoethanol at −80°C until use. DNAse-treated RNA was used for cDNA synthesis using Omniscript (Qiagen). Pig gene sequences were retrieved from TIGR project (
Statistical analysis
Comparisons of plating efficiency and cell line derivations at Passage 2 using different media were performed using Fisher's exact test. Unpaired t-test was used for comparing the effects of JAK inhibitor and LIF addition/blocking to the cultures.
Results
Isolation and establishment of pig epiblast stem cells
Pig epiblasts were dissected manually from embryos recovered at Days 10.5–12 after insemination. In pig embryos, the dark polar trophoblast cells (or Rauber's layer) shed off allowing good visualization of the epiblast under a stereomicroscope (Fig. 1A). During the dissection, the darker TE and the underlying globular PE layer were removed carefully using fine needles (Supplementary Fig. 1; Supplementary materials are available online at
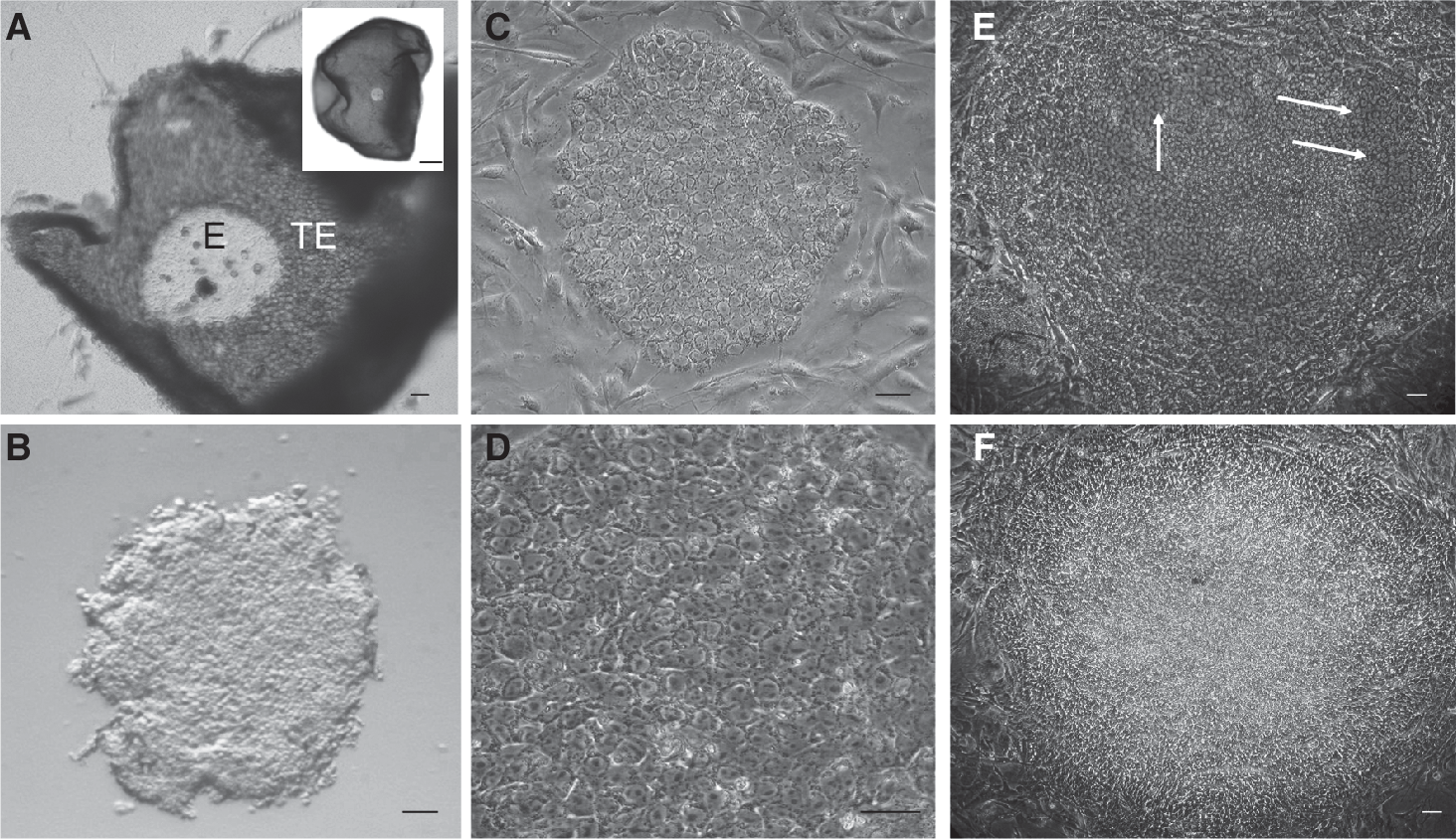
Isolation and culture of pig epiblasts.
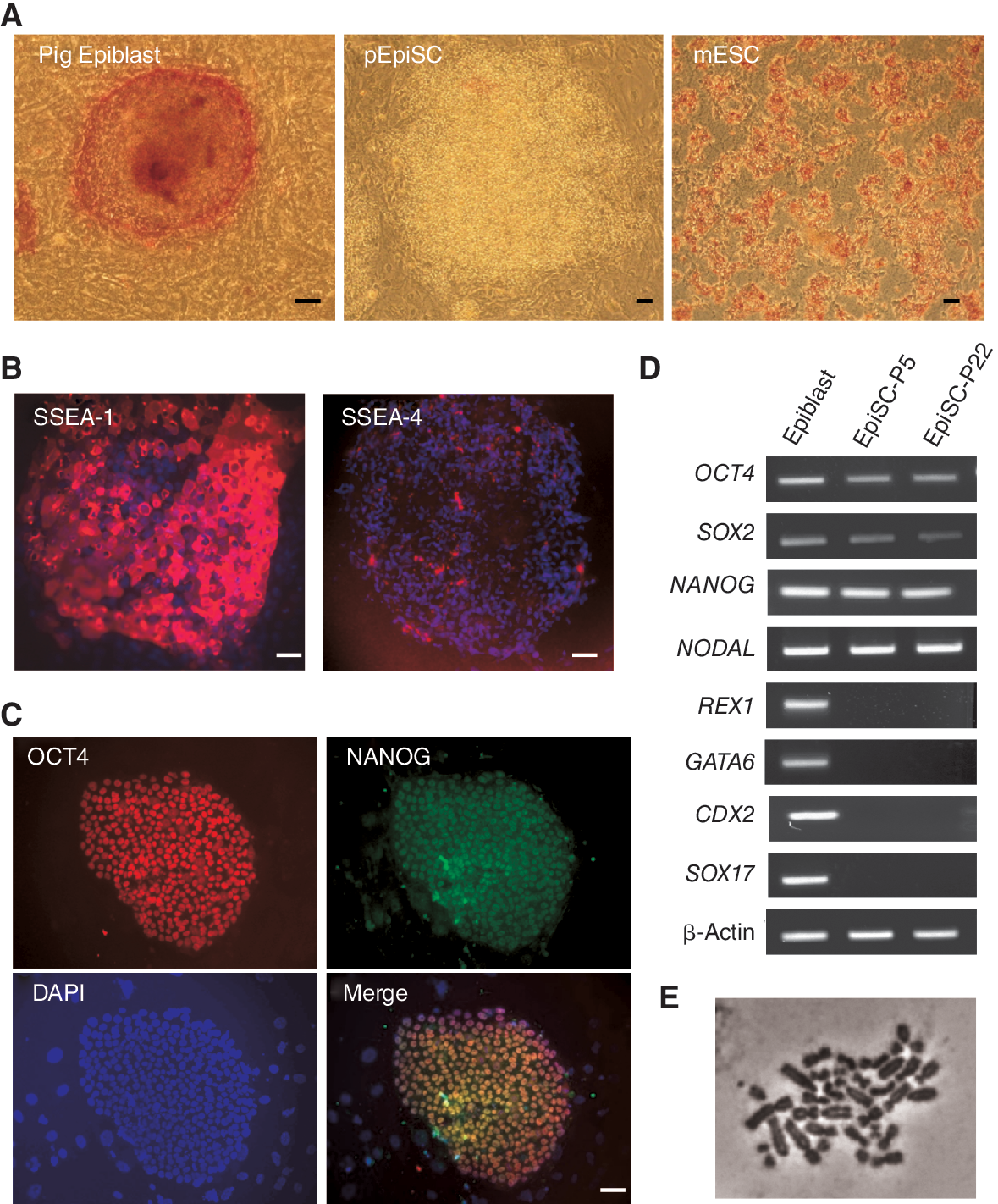
Characterization of pig epiblast stem cell lines (pEpiSC). (
Isolation in medium containing either FCS or SR supported the establishment of cell lines when bFGF was included in the medium. Increasing concentrations of FGF did not improve the derivation efficiency; therefore, long-term expansion was carried out with medium containing 5 ng/mL bFGF. Addition of exogenous Activin A or LIF did not improve the efficiency of derivations; therefore, they were not included after P2. More consistent derivations with reduced extraembryonic differentiation were obtained when conditioned medium containing FCS was used instead of conditioned medium with SR or unconditioned medium. Selected lines were cultured long term on mouse feeder layers either in medium with 20% SR or FCS supplemented with 5 ng/μL bFGF.
For initial characterization we tested AP activity, a conventional marker of mouse and hESC. Freshly isolated epiblasts and epiblasts plated for 12 h stained positive for AP. After P2, however, AP activity was no longer detectable in pEpiSC (Fig. 2A). This observation correlates with observations in mEpiSC that show no AP activity [6]. Surface marker expression shows that mEpiSC are SSEA-1-positive [6,19] and similar observations were reported for pig epiblasts [20]. We detected strong SSEA-1 expression in cultured epiblasts and weak or no expression of SSEA-4 (Fig. 2B). Expression of NANOG and SOX2 is lower in blastocysts compared with pig epiblasts [21,22]. Here we show that cultured epiblasts express OCT4, NANOG (Fig. 2C and 2D), SOX2, REX1, and NODAL (Fig. 2D), demonstrating that pig epiblasts express the core pluripotency genes. Interestingly, we also detected a number of extraembryonic markers such as GATA6, SOX17, and CDX2 in freshly isolated epiblasts. Established pEpiSC lines (Epi8, 21, and 42) also maintained expression of these pluripotency markers, except for REX1, and lost expression of the extraembryonic genes detected in isolated epiblasts (Fig. 2D). It is worth mentioning that REX1 is also a marker of PE in the pig [23], thus the expression in isolated epiblasts could be due to cells committed to this lineage.
The results from this initial characterization show that the expression profile of pEpiSC is similar to what was reported for mEpiSC [6], indicating that pEpiSC maintained epiblast-like characteristics after long-term culture. Furthermore, it suggests that under these culture conditions extraembryonic gene expression is repressed rapidly in these cultures.
Maintenance of self-renewal
Our initial cultures included LIF, although it was not clear whether these cells required this cytokine for self-renewal. pEpiSC express LIF and LIF-r (Supplementary Fig. 3; Supplementary materials are available online at
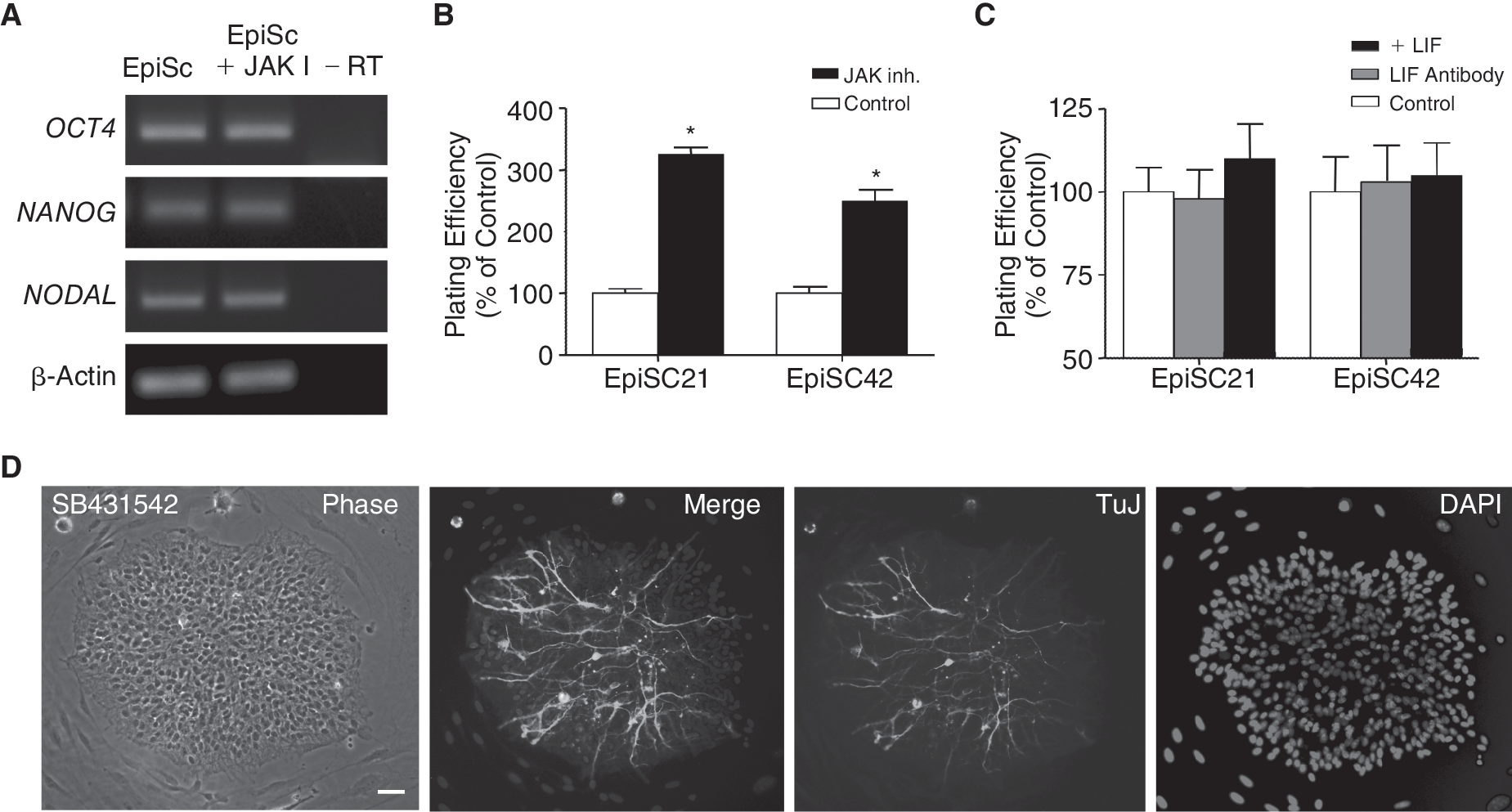
Signaling pathways required for pig epiblast stem cell lines (pEpiSC) pluripotency. (
Multilineage differentiation of pEpiSC
After having defined the growth conditions of undifferentiated pEpiSC, we tested their differentiation potential. First, we allowed spontaneous differentiation of 2 EpiSC lines (Epi21 (P7) and Epi42 (P9)) grown on feeder layers in medium without bFGF for 30 days. Cells with morphological characteristics of the 3 somatic germ layers were identified under phase contrast in different regions of the dishes (Fig. 4A). Because pEpiSC were differentiated on top of mouse feeder cells, we were not able to perform immunocytochemistry for identification of markers of somatic lineages, due to the false positives produced by the cross reactivity of the mouse cells to the specific antibodies. Instead, we extracted RNA from these cells and we analyzed the expression of a panel of markers from ectoderm, mesoderm, and endoderm using reverse transcriptase polymerase chain reaction (RT-PCR). The results show that pEpiSC can spontaneously differentiate to cells representative of the 3 somatic germ layers (Fig. 4B) and confirm the morphological characteristics of the cells (Fig. 4A). Cells with neural morphology did not express markers of other lineages; however, endoderm cells expressed low levels of PECAM, and mesoderm cells expressed low levels of KERATIN14 (data not shown), indicating that these cells formed mixed populations during differentiation. As further test for the differentiation potential of pEpiSC, we induced differentiation to the neural lineage using an established protocol [19]. pEpiSC colonies cut into small pieces were cultured in hanging drops to induce the formation of EB before plating and culturing in conditions that support differentiation of mouse and hESC to neural progenitors. Under these conditions, almost all cells arising from plated EB showed typical characteristics of neural progenitor cells and the neural markers TuJ and MAP2 were detected by immunocytochemistry (Fig. 4C).
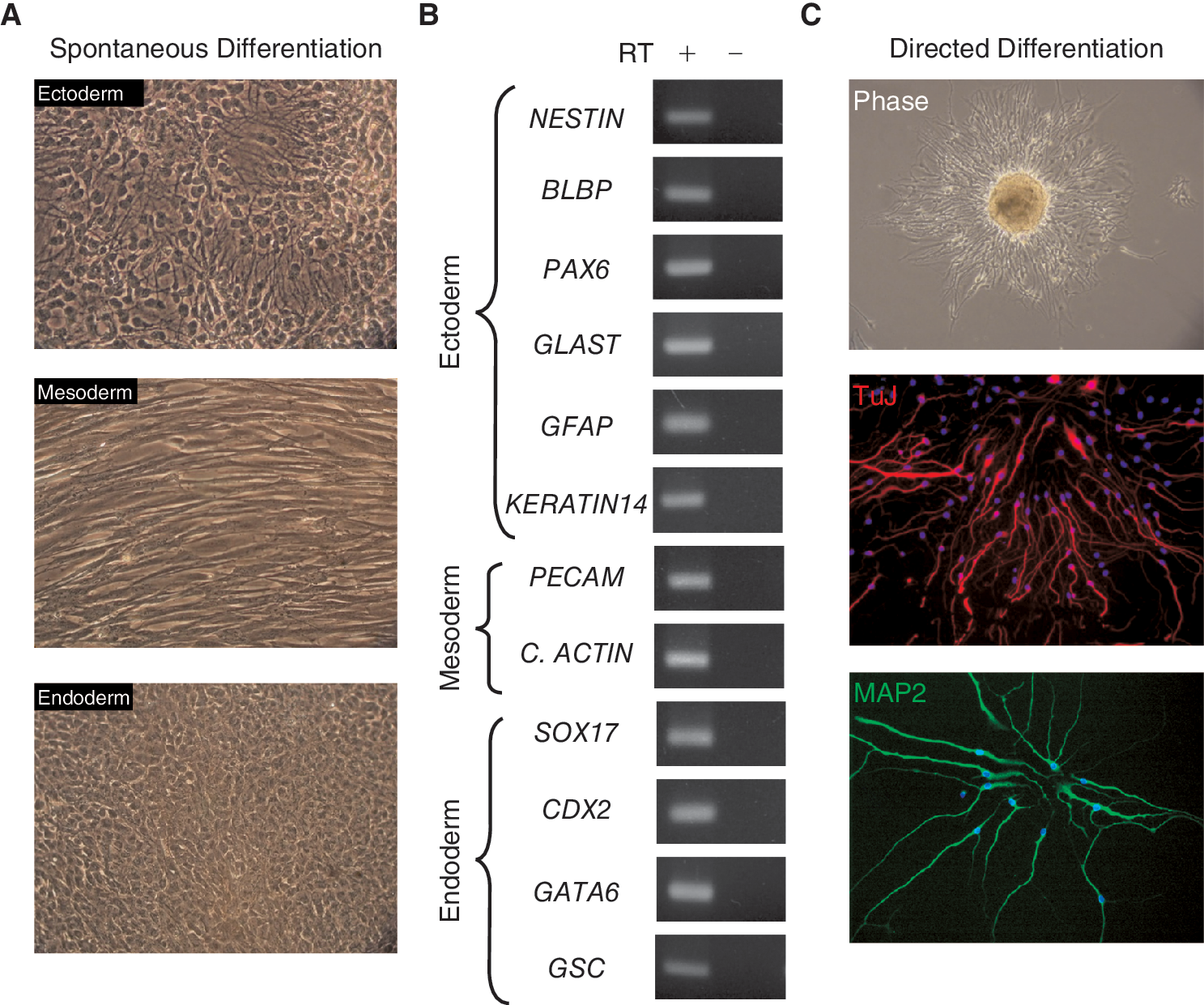
Differentiation potential of pig epiblast stem cell lines (pEpiSC).
We next investigated whether pEpiSC were able to differentiate into extraembryonic lineages following an induction protocol with BMP-4 as previously used with mEpiSC [5] and hESC [8]. EB made from pEpiSC for 6 days were plated onto gelatinized dishes and cultured for additional 8 days in medium supplemented with BMP-4 (Fig. 5A). Gene expression analysis of these cultures showed activation of the trophoblast markers EOMES, HAND1, H19, MSX2, and progesterone receptor (PGr), indicating that pEpiSC initiate trophoblast differentiation in response BMP-4 (Fig. 5B). BMP-4 is also effective in inducing differentiation of hESC to germ cell precursors [20]. Using a similar protocol to that used for hESC, we tested whether germ cell markers were induced from BMP-4-treated pEpiSC. EB kept in suspension for 6 days were plated with or without BMP-4 and cultured for 8 days before gene expression analysis (Fig. 5A). Specific germ cell markers DAZL and VASA were detected in BMP-4-treated groups (Fig. 5C). The meiotic marker SCYP3 [21], however, was not detected in these groups suggesting that extended cultures may be needed to initiate meiotic differentiation in vitro. These results indicate that pEpiSC have the potential to initiate the germ cell program upon induction with BMP-4. These results are in accord with reports in human and mESC differentiation toward germ cell lineage [9,22], indicating that the ability of BMP-4 to induce germ cells is conserved in mammals.
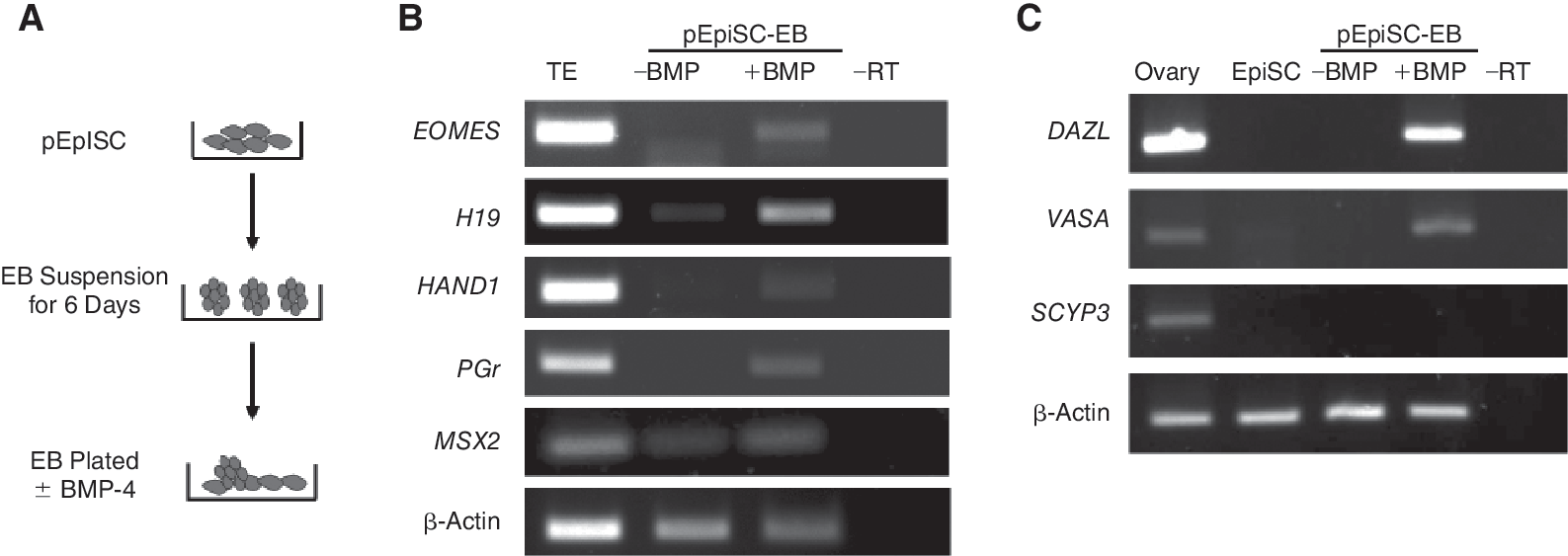
Differentiation to trophectoderm and germ cell lineage in response to BMP-4. (
Discussion
Here we report the isolation and in vitro growth requirements for pEpiSC. In contrast to the mouse and human counterpart, pig epiblast-stage embryos are not implanted and can be retrieved by flushing the uterus of pregnant animals. In the pig, like in other ungulates such as sheep and cattle, the epiblast progresses through gastrulation before implantation of the conceptus [23 –25]. The epiblast at this stage of development transitions from a round oblong disc to a developmentally advanced pear-shaped embryonic disc with a narrow posterior end [26]. Early round oblong epiblasts identified under a stereomicroscope can be easily separated from the remaining trophoblast cells on the basis of their contrasting appearance [27]. The underlying PE cells, which are transparent cells and have a globular phenotype, can be distinguished from the epithelial cells of the epiblast. Thus, the distinctive features of the bilaminar pig embryo allow the isolation of the individual components such as epiblasts, PE, and trophoblast.
Stem cells express a core group of genes that are critical for maintaining pluripotency [28]. Our analysis of pure epiblasts shows that these cells express the core genes NANOG, OCT4, and SOX2 and the epiblast marker NODAL, Previous studies have shown that pig blastocysts express OCT4; however, its binding partner SOX2 is expressed at low levels and NANOG is not expressed [29, 30]. OCT4/SOX2 proteins cooperate to activate the NANOG gene, suggesting that in pig blastocysts this interaction has not been established and therefore NANOG is not expressed. NANOG plays a critical role in establishing pluripotency in mammals [31]. In mouse blastocysts, NANOG is expressed in conjunction with OCT4 and SOX2 conferring pluripotent properties to the ICM [32, 33]. In the pig the concerted expression of these factors is delayed until the epiblast stage [27] suggesting that the ICM is a transitional stage that has not yet acquired pluripotency. The lack of pluripotency in the pig ICM may explain the difficulty in establishing ESC from blastocysts. Previous reports have demonstrated that pig ICM derived from Day 8 blastocysts transit to an epiblast phenotype after 2–3 days in culture [34] and they rapidly differentiate [27]. Here, we cultured isolated epiblasts using conditions that support hESC and mEpiSC growth. We demonstrate that pEpiSC depend on Activin/Nodal signaling for self-renewal, and that the LIF/JAK/STAT3 pathway is not essential for pluripotency. The response to the Activin/Nodal inhibition, consisting in rapid differentiation to the neural lineage, is similar to the response of mEpiSC and hESC cultured under similar conditions [5, 35]. In contrast, blocking of LIF/JAK/STAT3 pathway failed to interfere with self-renewal and pluripo-tency of pEpiSC, indicating that this pathway does not play a central role in sustaining pluripotency in pEpiSC. These results are in agreement with previous findings with mEpiSC and hESC [4 –6,10]. Interestingly, we found that pEpiSC have increased plating efficiency when grown with the specific JAK I inhibitor, similar to a previous report in mEpiSC [6], suggesting that inhibition of JAK/STAT3 signaling can have a beneficial effect in growth of EpiSC. Indeed, similar observations in hESC indicate that activation of STAT3 may promote differentiation and apoptosis in these cells [4,10], and therefore the inhibition of this pathway may be beneficial for the in vitro growth of Activin/Nodal-dependent stem cells. Blocking LIF signaling with an anti-mLIF antibody did not improve plating efficiency, suggesting that the small molecule inhibitor is more specific and robust in its inhibitory function. Conversely, addition of mLIF did not result in increased plating efficiency, suggesting that either mLIF is not biologically active in pig cells, or LIF signaling is not critical for maintaining pEpiSC. Furthermore, our functional tests confirm previous observations showing that addition of het-erologous LIF does not prevent differentiation of pig ESC [36, 37].
The surface marker SSEA-1 was readily detected in pig epiblasts, as shown previously [24, 36] and SSEA-4 was very faint, indicating that the pattern of surface markers resembles that of mEpiSC [5] and is the opposite of hESC [7]. Alkaline phosphatase activity was detected in plated epiblasts, but not in pEpiSC after a few passages, similar to the observations in mEpiSC [5]. AP activity in isolated epiblasts was stronger in peripheral cells suggesting that these cells may have a different cellular identity to the rest of the epiblast. Indeed, analysis of isolated epiblasts shows expression of the extraembryonic markers GATA6, SOX17, REX1, and CDX2, all of which are not expressed in pEpiSC. The expression of the extraembryonic genes in dissected epiblasts could either be due to the presence of few contaminating TE or PE cells left after the isolation procedure, or alternatively due to the presence of cells within the epiblast expressing these genes. Although we cannot exclude the first possibility we favor the latter, since the dissections resulted in consistently pure epiblast cells. Furthermore, during the establishment of the pEpiSC we systematically observed that plated epiblasts initially differentiated into extraembryonic cells that surrounded a center of stem cells. The epiblasts dissected from these outgrowths plated onto new feeders began to grow homogeneously, without differentiation, which suggests that the cells giving rise to these extraembryonic cells were already committed when the epiblasts were plated. The systematic differentiation of the peripheral cells of the cultured epiblasts into extraembryonic cell types supports the idea that some cells within the margins of the epiblast may be primed to contribute to extraembryonic cell types. This is supported by observations that GATA6 is detected in some cells of pig ICM, as a prelude of the segregation of the PE [29]. It is tempting to speculate that the peripheral cells of the epiblast may be contributing to the sustained growth of the trophoblast and the PE during these stages of development, explaining the expression of markers of these tissues in our assays, and the rapid differentiation of plated epiblasts. The idea of marginal epiblast cells with different identity has been proposed recently for the rabbit [38] and our findings in the pig support these observations. Ongoing experiments in our laboratory are focused at establishing the identity of the cells in the pig epiblast.
We show that pEpiSC can spontaneously differentiate to cells expressing markers of 3 somatic germ layers and they can also respond to a differentiation protocol for inducing neural differentiation. Attempts to induce mesoderm and endoderm did not achieve consistent results, although this was not totally unexpected since results with hESC shows that directed differentiation into specific lineages can prove very challenging with some cell lines [39], and therefore further work is needed to establish the proper differentiation conditions for pEpiSC into these lineages.
Differentiation to the trophoblast lineage has been demonstrated for mEpiSC [5] and hESC [8] in response to BMP-4. Our results show that pEpiSC also activate trophoblast genes in response to BMP-4, demonstrating that these cells can give rise to all somatic germ layers and trophoblast. As a last test for the in vitro differentiation potential of pEpiSC, the contribution to the germ cell lineage was studied. We show that pEpiSC can reactivate the specific germ cell markers DAZL and VASA after induction with BMP-4. The capacity of BMP-4 to induce germ cells has been shown in vivo in the mouse [40] and in vitro with hESC [20]. Thus, our results with pEpiSC show that the role of BMP-4 as an inducer of the germ cell lineage is conserved.
In conclusion, our study shows that the epiblast of the pig embryo is pluripotent, and can be isolated and cultured long term. Cultured pig epiblasts depend on Activin/Nodal signaling for self-renewal, as previously shown for mEpiSC and hESC, indicating that maintenance of pluripotency by this signaling mechanism is conserved in mammals.
Footnotes
Acknowledgments
This study was funded by the University of Nottingham and the Royal Society. Part of the work shown here was presented at the ISDB Congress, September 2009 (Edinburgh, United Kingdom).
Author Disclosure Statement
The author's declare that there is no conflict of interest that would prejudice the impartiality of this work.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.