
Abstract
We report the toxicological and pharmacokinetic properties of the synthetic, small interfering RNA I5NP following intravenous administration in rodents and nonhuman primates. I5NP is designed to act via the RNA interference (RNAi) pathway to temporarily inhibit expression of the pro-apoptotic protein p53 and is being developed to protect cells from acute ischemia/reperfusion injuries such as acute kidney injury that can occur during major cardiac surgery and delayed graft function that can occur following renal transplantation. Following intravenous administration, I5NP was very rapidly cleared from plasma was distributed predominantly to the kidney, with very low levels in liver and other tissues. Doses of 800 mg/kg I5NP in rodents, and 1,000 mg/kg I5NP in nonhuman primates, were required to elicit adverse effects, which in the monkey were isolated to direct effects on the blood that included a sub-clinical activation of complement and slightly increased clotting times. In the rat, no additional adverse effects were observed with a rat analogue of I5NP, indicating that the effects likely represent class effects of synthetic RNA duplexes rather than toxicity related to the intended pharmacologic activity of I5NP. Taken together, these data support clinical testing of intravenous administration of I5NP for the preservation of renal function following acute ischemia/reperfusion injury.
Introduction
I5NP is manufactured synthetically. Its structure consists of a blunt-ended 19-base-pair RNA duplex that is partially protected from nuclease degradation using a methoxy modification on the 2′ position of the ribose sugar (Czauderna et al., 2003). This modification occurs naturally in mammalian cells (Starr and Sells, 1969).
I5NP siRNA is designed to act via the RNA interference (RNAi) pathway to temporarily inhibit expression of the p53 protein in order to delay p53 pro-apoptotic activity following renal injury. In cells, the antisense strand of I5NP is incorporated into the RNA-induced silencing complex (RISC), and the RISC-I5NP complexes then proceed to destroy p53 mRNA, which temporarily inhibits expression of the p53 protein. The temporary inhibition of p53 expression by I5NP affords kidney cells time to repair cellular damage following reperfusion injury and thereby avoid induction of apoptosis. Cells that are irreversibly damaged, or that have accumulated deleterious mutations, are later eradicated when the effects of I5NP have subsided and p53 expression levels return to normal which, in rat kidneys, occurred two days after intravenous (i.v.) administration of a rat-specific active analogue of I5NP (Molitoris et al., 2009).
Herein, we report the results from nonclinical pharmacokinetic and toxicity studies used to support first-in-man clinical trials.
Materials and Methods
siRNAs
The molecular sequence of I5NP is:
Passenger (sense) strand 5′-GaGaAuAuUuCaCcCuUcA-3′ Guide (antisense) strand 5′-uGaAgGgUgAaAuAuUcUc-3′
Molecular sequence of the QM5 rat analogue is:
Passenger (sense) strand 5′-GaAgAaAaUuUcCgCaAaA-3′ Guide (antisense) strand 5′-uUuUgCgGaAaUuUuCuUc-3′
Uppercase letters represent unmodified RNA nucleosides and lowercase letters represent 2′-O-methyl sugar modified RNA nucleosides.
I5NP was manufactured by Avecia Biotech (Milford, MA) and QM5 was manufactured by Avecia or Agilent Technologies (Boulder, CO).
siRNAs were provided as bulk powder and formulated on the day of dosing in commercially available phosphate-buffered saline (PBS) to the nominal concentrations indicated using a factor to correct for purity and oligonucleotide content. The formulation accuracy and homogeneity of dosing solutions were verified by analysis of dose formulation samples, conducted by Pyramid Laboratories (Costa Mesa, CA).
Quantification of I5NP in plasma and tissues
A sandwich hybridization assay was developed by Charles River Laboratories Preclinical Services Montreal Inc. to specifically quantify the antisense strand of I5NP, which is considered to be directly proportional to the concentration of duplex I5NP on a molar basis since both strands of I5NP are present in equivalent amounts in the hybridized duplex. The method involves hybridization of the 3′ end of the antisense strand of I5NP to a 10-nucleotide (nt) “capture probe” tethered to the surface of a 96-well plate, and subsequent hybridization of the 5′ end of the antisense strand to a biotinylated 9-nt “detection probe.” A horseradish pyroxidase-streptavidin conjugate is then used for detection in combination with the colorimetric substrate 3,3′,5,5′ tetramethylbenzidine.
The method was fully validated in human plasma and partially validated for monkey and rat and plasma. In addition, a similar type of method was validated for rat kidney and qualified for rat liver. The rat kidney method was used to evaluate levels in all tissues except liver.
2-Photon confocal microscopic imaging
Imaging was performed by InPhoton, LLC (www.inphoton.com).
Toxicity studies
A list of studies performed (excluding genotoxicity studies) is provided in Table 1. The monkey safety pharmacology study was performed by Charles River Laboratories, Nevada. Genetic toxicity (bacterial reverse mutation and chromosomal aberration) assays were performed by BioReliance, Inc. (Rockville, MD). All other studies were performed at Charles River Laboratories, Montreal.
i.v., intravenous; NA, not applicable; PBS, phosphate buffered saline; sac, sacrificed; TK, toxicokinetic.
Safety pharmacology study in monkeys
The effects of I5NP on the cardiovascular, respiratory, and nervous systems were assessed in conscious telemeterized monkeys. Four experimentally naive male cynomolgus monkeys (Scientific Resources International, Ltd., Reno, NV), 3 to 5 years of age, and weighing 3.5 to 3.7 kg at the outset (Day –1) of the study, were assigned to a single dose group. At least 1 week prior to initiation of dosing, a telemetry transmitter (Model No. TL11M2-D70-PCT or TL11M3-D70-PCTP, Data Sciences™ International, St. Paul, MN) was surgically implanted into each animal for monitoring and recording of cardiovascular parameters. Electrocardiogram (ECG) leads were placed to collect lead-2 ECG data. In addition, a catheter was implanted into an artery and routed to a subcutaneous access port, which was used to obtain arterial samples for determination of blood gases (i.e., partial pressures of CO2 [PaCO2] and O2 [PaO2], pH, and percentage saturation of hemoglobin with oxygen [SaO2]).
The first day of dosing was designated Day 1. Each animal received 4 separate IV doses of the control article (PBS 1×) on Day 1 (at 0, 2, 6, and 14 hours) and 4 i.v. doses (at 0, 2, 6, and 14 hours) of the test article each on Days 4 (5 mg/kg/dose) and 11 (25 mg/kg/dose). The reason for administering 4 doses over 14 hours on each occasion was to provide flexibility in the dosing regimen of I5NP in the phase 1 clinical studies. The animals were observed for changes in general health (twice daily and within 10 minutes following each dose), food consumption (once daily), body weight (Days 1, 3, and 10), cardiovascular data (obtained via telemetry; prior to, during and following dosing on Days 1, 4, and 11), respiratory function (prior to and following dose administration) and neurological evaluations (prestudy and within 24 hours after the end of the telemetry recording period following the last dose).
Blood samples were collected for serum chemistry and hematology (prestudy and approximately 10 hours following the last dose on each dosing day), coagulation (prior to the first dose and 5 minutes, 1 hour and approximately 10 hours after the 4th dose on each dosing day), blood gas (prior to the 1st dose, prior to the 3rd dose and approximately 10 hours after the 4th dose on each dosing day) and complement Bb analysis (Days 1 and 11: prior to the first dose, 5 minutes after the first dose, prior to the fourth dose and 5 minutes and approximately 10 hours after the fourth dose; Day 4 prior to the 1st dose, 5 minutes after the first dose, and 5 minutes after the fourth dose). In addition, blood samples were collected for toxicokinetic analysis (plasma concentrations of I5NP) prior to the first dose, at 5 minutes after the first and last doses and at one hour after the last dose on Days 4 and 11. At the end of the study (Day 13), the animals were returned to the testing facility telemetry animal colony.
Sub-acute toxicity and pharmacokinetic study in rats
Rats (Sprague Dawley; Charles River Canada, St-Constant, Quebec, Canada) were administered PBS, I5NP (3, 15, or 60 mg/kg/dose) or the rat analogue QM5 (15 or 60 mg/kg/dose) by i.v. injection (slow push over approximately 30 seconds), via a tail vein, at 4 time points (0, 2, 6, and 14 hours) on Day 1. As described for the safety pharmacology study above, the reason for administering 4 doses over 14 hours was because to provide flexibility for the phase 1 dosing regimen. At the start of treatment, animals were 8 weeks of age and ranged in weight as follows: males 250 to 315 g; females 166 to 229 g. There were 10 animals per sex in the PBS and I5NP dose groups and 5 per sex in the QM5 dose groups. All of these animals were sacrificed on Day 3 of the study. Additional recovery animals (5 per sex) were included in the control and high-dose I5NP and QM5 groups for recovery assessment, and an additional 6 per sex were included in the I5NP groups for toxicokinetic sampling. Recovery animals were maintained untreated for a 15-day recovery period following the last dose.
The following parameters were evaluated: clinical signs (daily during the treatment/recovery phase), body weight (twice pretreatment and on Days 3, 9, and 14), food consumption (weekly), hematology (Days 3 and 17), serum chemistry (Days 3 and 17), urinalysis (Days 3 and 17), toxicokinetics (plasma and tissue drug levels; samples collected from dedicated animals), macroscopic observations at necropsy, organ weights, and histopathology. In addition, bone marrow was collected from the main study animals and evaluated for micronuclei, as an indicator of potential genotoxicity of I5NP and QM5.
Sub-acute toxicity and pharmacokinetic study in nephrectomized rats
Thirty (15 male, 15 female) non-nephrectomized and forty (20 male, 20 female) nephrectomized Sprague-Dawley rats were obtained from Taconic (Germantown, NY). Six per sex received either 10 mg/kg of I5NP or PBS, as 4 injections given at 0, 2, 6, and 14 hours. At the onset of treatment, the animals were all approximately 12 to 13 weeks old. Body weights ranged from 317 to 527 g for all treated males and from 204 to 279 g for all treated females. Nephrectomy consisted of a 2-step surgical procedure involving removal of 2/3 of one kidney, followed by complete removal of the remaining kidney. Control (“non-nephrectomized”) animals were not subjected to any surgical procedures.
The following parameters were evaluated: clinical signs, body weight, hematology, clinical biochemistry, pharmacokinetics, and tissue distribution of I5NP, gross pathology, organ weights and histopathology.
Acute toxicity and pharmacokinetic study in rats
Rats (Sprague Dawley; Charles River Canada, St-Constant, Quebec, Canada) were administered PBS, I5NP (200, 800, 1,200, or 2,000 mg/kg) or QM5 (1,200 mg/kg) by i.v. injection. There were 10 animals per sex in the PBS and I5NP dose groups and 5 per sex in the QM5 dose groups. Animals were sacrificed on Day 2 of the study. Additional recovery animals (5 per sex) were included in the control and high-dose I5NP and QM5 groups for recovery assessment, and an additional 4 per sex were included in the I5NP groups for toxicokinetic sampling. Recovery animals were maintained untreated for a 15-day recovery period.
The following parameters were evaluated: clinical signs (twice daily), body weight (randomization, the day prior to dosing, Days 2, 9, and 16), food consumption (prior to treatment, and weekly thereafter), hematology (at necropsy), serum chemistry (at necropsy), urinalysis (Days 2 and 16), toxicokinetics, macroscopic observations at necropsy, organ weights, and histopathology.
Sub-acute toxicity and pharmacokinetic study in monkeys
Monkeys (cynomolgus; Rhenos, LLC, Chestnut Hill, MA) were administered PBS or I5NP (3, 12, or 48 mg/kg/dose) by i.v. injections (slow push over approximately 30 seconds) at 4 time points (0, 2, 6, and 14 hours) into the brachial veins. At the start of treatment, the animals ranged in weight as follows: males 2.0 to 2.8 kg; females 1.8 to 2.1 kg. The age of the animals received for this study ranged from approximately 2.5 to 3 years old. After dosing, the animals were sacrificed on Day 3 (3 per sex per group) or Day 17 (2 per sex in control and high-dose groups).
The following parameters were evaluated: clinical signs (daily during the treatment/observation phase), body weight (twice pretreatment and on Days 3 and 13), appetence (daily), ophthalmology (predose and Day 3), electrocardiograms (predose and Day 3), hematology (predose, Days 3 and 17, with coagulation parameters additionally assessed 3 times on Day 1), serum chemistry (predose, Days 3 and 17), urinalysis (Days 3 and 17), complement factor analyses (3 time points on Day 1), toxicokinetics, macroscopic observations at necropsy, organ weights, and histopathology.
Acute toxicity and pharmacokinetic study in monkeys
Monkeys (cynomolgus; Charles River Laboratories Preclinical, Sparks, NV) received single i.v. injections of vehicle (PBS), 200, 500, or 1,000 mg/kg I5NP. At the start of treatment, animals were approximately 3 to 4 years old and ranged in weight as follows: males 2.3 kg to 3.2 kg; females 2.1 kg to 2.7 kg. There were 3 animals per sex in all dose groups, and 2 animals per sex in the control and 1,000 mg/kg dose groups for assessment of recovery. Main study animals were sacrificed on Day 2, and the recovery animals were maintained untreated for a 15-day recovery period.
The following parameters were evaluated: mortality and signs of ill health and reaction to treatment (twice daily), clinical signs (pretreatment and Days 1, 2, and 13), body weights (twice pretreatment and on Days 2 and 15), food evaluation (daily from Day –7 extending to the end of recovery period), ophthalmology examinations, and electrocardiography (predose and Day 2), hematology (predose and Days 2 and 16, with coagulation parameters additionally assessed 3 times on Day 1), serum chemistry (predose and Days 2 and 16), urinalysis (at termination), complement factor (C3a and Bb) analyses (pre and post dose), toxicokinetics, macroscopic observations at necropsy, organ weights, and histopathology.
Genetic toxicology studies
The potential genotoxicity of I5NP was investigated in a standard battery of in vitro and in vivo genetic toxicity studies including a bacterial reverse mutation assay in Salmonella and Escherichia coli, chromosome aberration assay in human lymphocytes, and a micronucleus study (included in the subacute rat study).
Results
Plasma clearance
I5NP elimination from the plasma was assessed in several i.v. toxicity studies in rats and monkeys (Table 1). Rats received i.v. doses ranging from 3 to 1,200 mg/kg. In the monkey studies, i.v. doses ranged from 3 to 1,000 mg/kg. One of the rat studies (460418) included an evaluation of pharmacokinetics of I5NP in renally impaired animals (5/6-nephrectomized model). Two of the rat and two of the monkey studies were subacute, repeat-dose studies where the first dose was followed by three additional administrations over a 14-hour period.
No significant gender differences were apparent for the I5NP plasma concentration data; hence, mean data are discussed in terms of the overall group averages (sexes combined).
Plasma clearance in normal animals
As expected, plasma concentrations of I5NP were highest at the first sampling time point 5 minutes post-dosing (Fig. 1). With doses of 200 mg/kg or lower, I5NP plasma concentrations declined dramatically (>90%) within 30 minutes post-dosing, and by 2 hours after the first dose, the mean concentrations were reduced by greater than 98% (in most cases by greater than 99.5%) of the 5-minute means. At the 800 and 1,200 mg/kg doses in the rat (Fig. 1A) and the 500 and 1,000 mg/kg doses in the monkey (Fig. 1B), I5NP plasmas levels declined substantially (>40%) within 15 minutes; however, the elimination was slower than occurred with the lower doses, suggesting a partial saturation of the clearance mechanisms. By 2 hours post-dosing, plasma levels in these high-dose animals were less than 10% of the respective 5-minute plasma levels. In the repeat-dose studies, no sign of accumulation of I5NP with repeat dosing was observed (data not shown), which is not surprising in view of the rapid clearance.
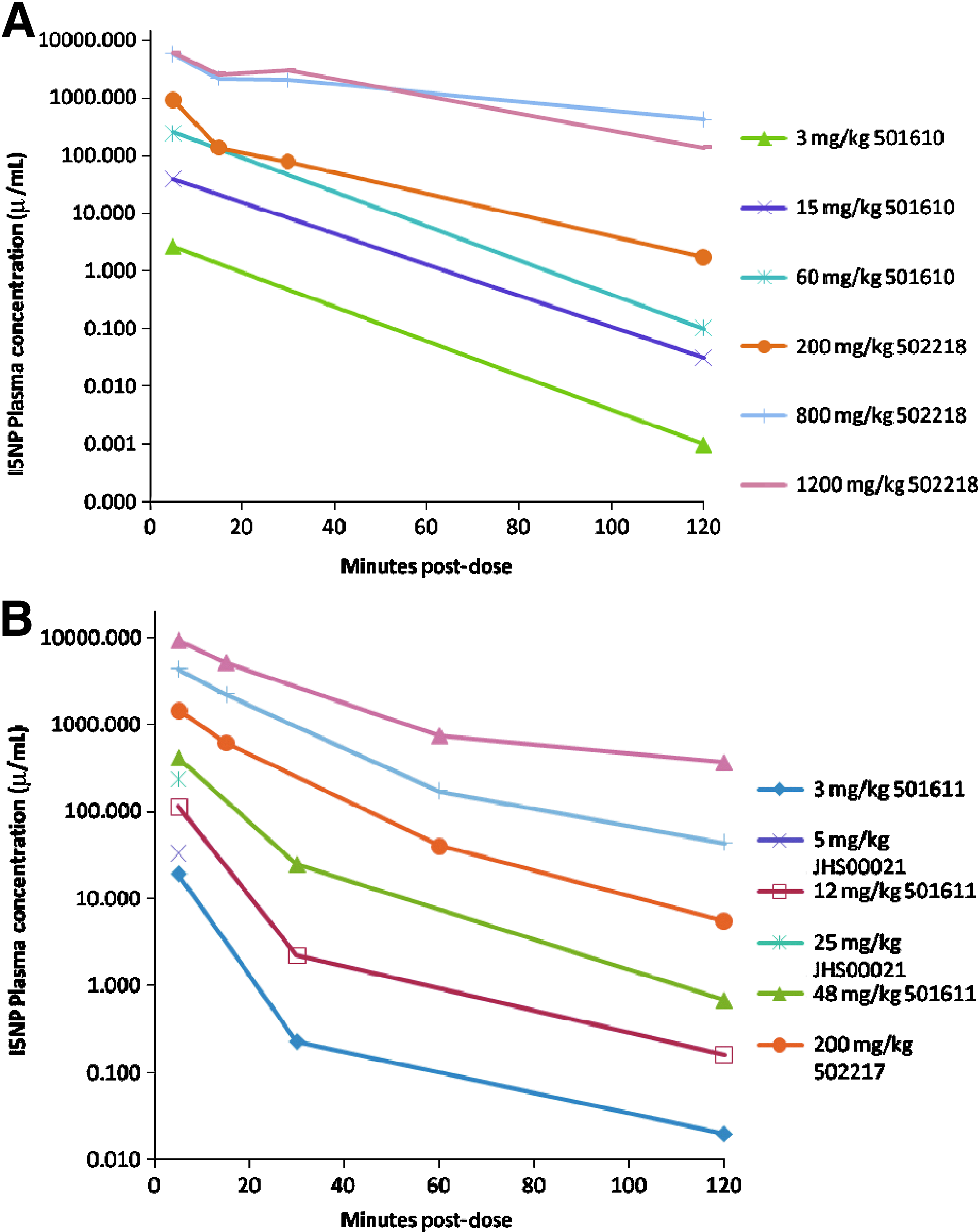
The plasma half-life for the early distribution phase in the i.v. studies was estimated to be less than 8 minutes. Clearance of I5NP from the plasma appeared to be multi-compartmental, with an apparent short initial distribution phase of approximately 15–30 minutes. A terminal (presumed elimination) phase in the rat could be identified. The percent area under the concentration time curve (AUC0-inf) extrapolated from the last observed concentration was below 1%, and the apparent terminal half life (t1/2) values for I5NP in plasma ranged between 0.4 and 0.5 hours. The overall mean clearance value (CL) was 599 mL/hr/kg for males and females combined. The mean apparent volume of distribution (Vz) was 420 mL/kg in males and females combined.
Plasma clearance in renally impaired rats
The rate of I5NP plasma clearance was slightly reduced in renally impaired vs. normal animals (Fig. 2). Between 5 and 15 minutes after the last injection, mean I5NP concentration (sexes combined) fell by approximately 98% and 89% relative to the 5-minute mean values for the non-nephrectomized and nephrectomized groups, respectively. It is noteworthy that the rate of decline of I5NP plasma concentration during this initial phase was still remarkably fast in 5/6-nephrectomized animals. While there was a clear quantitative impact of nephrectomy on this initial clearance compartment, removal of 5/6 of the kidney mass did not preclude a rapid decline in plasma levels, and it is reasonable to conclude that there was no important qualitative difference in the overall pharmacokinetic profile induced by partial nephrectomy.
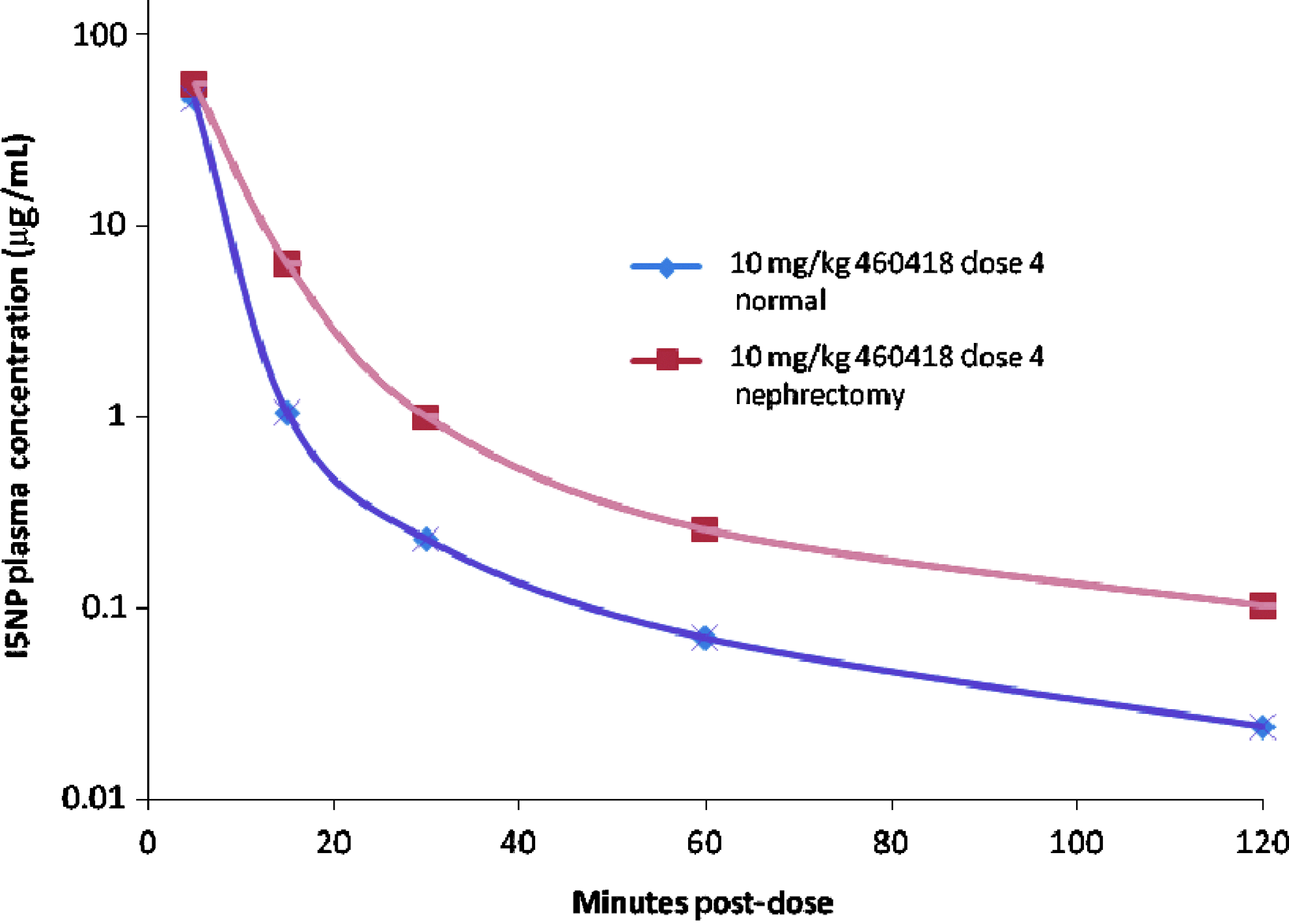
I5NP plasma concentrations in normal and partially (5/6) nephrectomized rats following the fourth subacute i.v. dose of I5NP.
Pharmacokinetic parameters were calculated for the data obtained from samples collected after the fourth dose from Study 460418. The mean calculated area under the concentration versus time curves (AUC0-tlast) for normal animals was 18,341 ng•h/mL and 18,381 ng•h/mL for males and females respectively, and 16,746 ng•h/mL and 16,132 ng•h/mL for nephrectomized males and females respectively. The apparent lower value of mean AUC in nephrectomized animals is partly due to lower exposure in 2 rats, combined with a small number of animals used to calculate the mean. Therefore, it was considered that the difference in AUC between nephrectomized and non-nephrectomized groups was not biologically significant. A terminal (presumed elimination) phase could be identified, and the percent AUC0-inf extrapolated from the last observed concentration was below 1%. The apparent terminal t1/2 values for I5NP in plasma ranged between 0.4 and 0.5 hours for both males and females. Mean overall CL values were similar in normal and nephrectomized rats (599 to 654 mL/hr/kg, respectively). The mean apparent volume of distribution was also similar in normal and nephrectomized rats (420 to 483 mL/kg, respectively).
Tissue distribution of I5NP
Tissue distribution in normal rats
Tissue distribution was characterized following i.v. injection of I5NP in the subacute repeat-dose rat study, wherein rats received four administrations of 3, 15, or 60 mg/kg of I5NP over 14 hours. In this study, tissues for analysis of I5NP concentrations were collected upon sacrifice at 2 hours after the first dose, 4 hours after the second dose (i.e., immediately prior to the third dose), and 2 hours after the fourth (last) dose. No consistent gender differences in I5NP concentration in tissues were apparent; hence, the results for the sexes were combined.
The mean I5NP levels measured in kidney samples were much higher than in other tissues (Table 2). At 60 mg/kg/dose, the mean concentration of I5NP in kidney was disproportionately greater than would be predicted from a linear dose relationship. However, the inter-animal variability was quite large (individual animal values at 2 hours after the first or fourth doses varied from ∼1,000 ng/g to ∼66,000 ng/g), which raised some uncertainty about the accuracy of the group mean values.
LLOQ=lower limit of quantification (generally 7 ng/g; 10 or 20 ng/g for some tissues).
The majority of the kidney data were derived from assays where quality control or dilution control samples failed to meet acceptance criteria; hence, the values are considered to be qualitative.
Group means in data sets containing values below the limit of quantification were calculated by assuming a value identical to the LLOQ, unless more than half of the samples for males or females were less than the LLOQ, in which case the group mean was not calculated (NC).
For the groups that received 3 mg/kg/dose and 15 mg/kg/dose, the I5NP concentrations measured in the kidney tended to be greater 2 hours after the first or fourth doses, as compared to 4 hours after the second dose. There was no indication of accumulation of I5NP in kidney tissue with repeated doses, and the relatively lower levels at 4 hours after the second dose (as compared to 2 hours after the first dose) suggest that I5NP deposited in the kidney was undergoing clearance between 2 and 4 hours post-dose and that the levels may have been higher at an earlier time point following dosing. The greater uptake by kidney tissue is consistent with previous findings about the tissue distribution of oligonucleotides (Geary et al., 1997a).
For all other tissues, I5NP levels were quite low at all sample collection time points and across all dose levels. There was no indication of accumulation in lung, heart, brain, testes, ovaries, colon, and bone marrow with 4 successive doses (i.e., mean values that were somewhat higher after the fourth dose occurred randomly and are considered to reflect individual animal variation rather than accumulation; Table 2). For the liver, I5NP concentration was generally below the limit of quantification, except for the high dose (60 mg/kg/dose) group at 2 hours after the fourth dose, for which the mean was low (37 ng/g) and only marginally above the limit of quantification (7 ng/g).
The significant kidney tissue levels described above are also consistent with 2-photon confocal microscopy results observed following i.v. administration of a Cy3-labeled siRNA to rats. Confocal microscopy enables high resolution, 3-dimensional imaging of cells, tissues and organs in living animals. In these studies, siRNAs comprising the same chemical composition as I5NP were labeled with the Cy3 fluorescent dye and injected intravenously into rats. Following administration, labeled siRNA was found to be filtered readily from the blood into the glomerular space (Fig. 3A–C), and then rapidly re-absorbed by proximal tubule cells (Figs. 3D, 4). siRNA accumulated almost exclusively in the proximal tubule cells, reaching peak levels between 5 and 30 minutes after administration, diminishing rapidly after 2 hours, and becoming undetectable by 24 hours post-administration (Fig. 4, Molitoris et al., 2009). These data demonstrating rapid uptake and clearance of siRNA from the kidney are in good agreement with the persistence of the pharmacodynamic effect of the rat analogue on reduction of p53 mRNA in the rat kidney, where p53 mRNA levels were reduced within 3–6 hours after i.v. administration and returned to basal levels within 48 hours after injection (Molitoris et al., 2009).
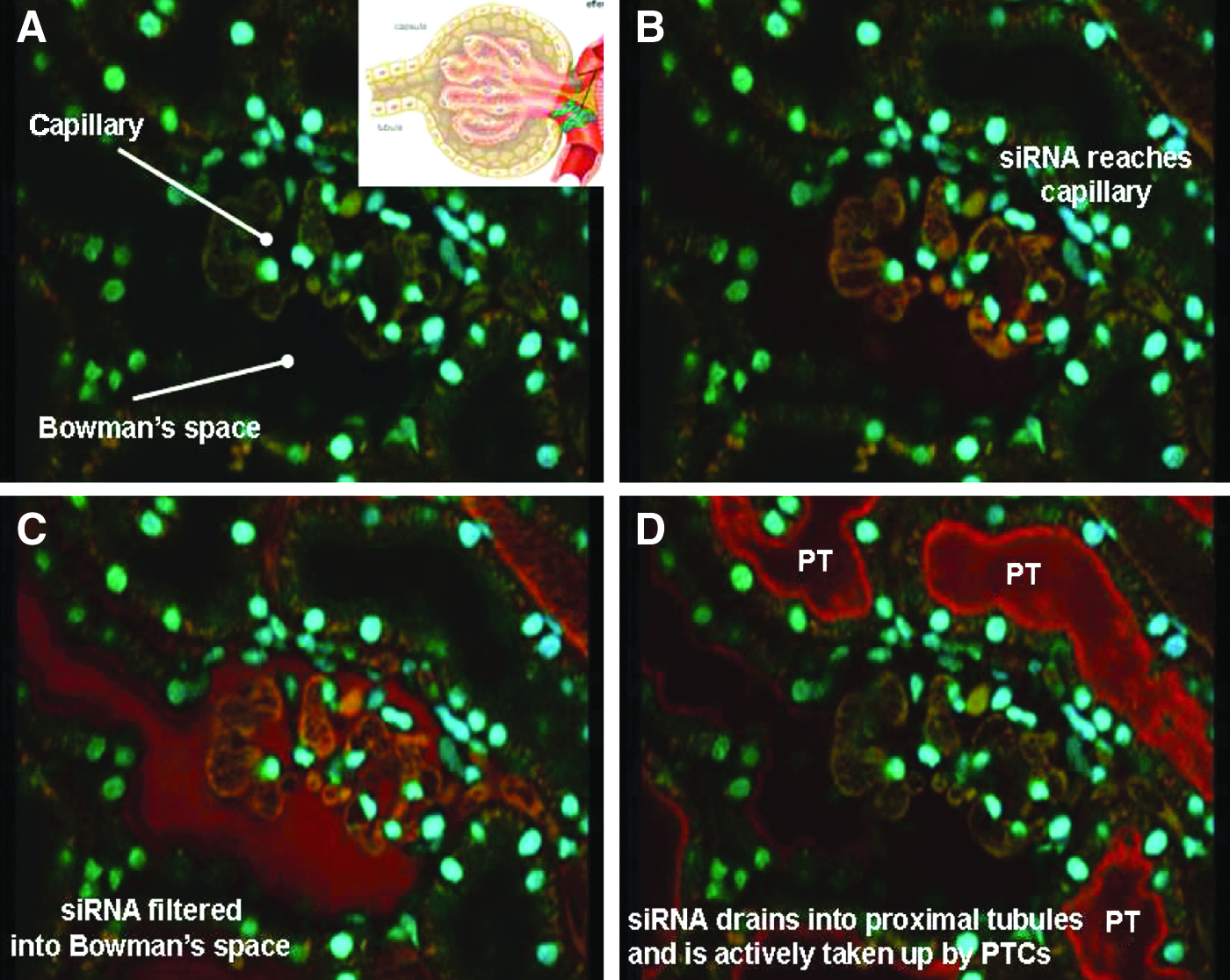
In vivo (2-photon microscopy) imaging of uptake of fluorescent-labeled small interfering RNA (siRNA) in the rat kidney following i.v. administration. Fluorescent-labeled siRNA was administered intravenously (1 mg/kg) via a 3-second infusion into the tail vein.
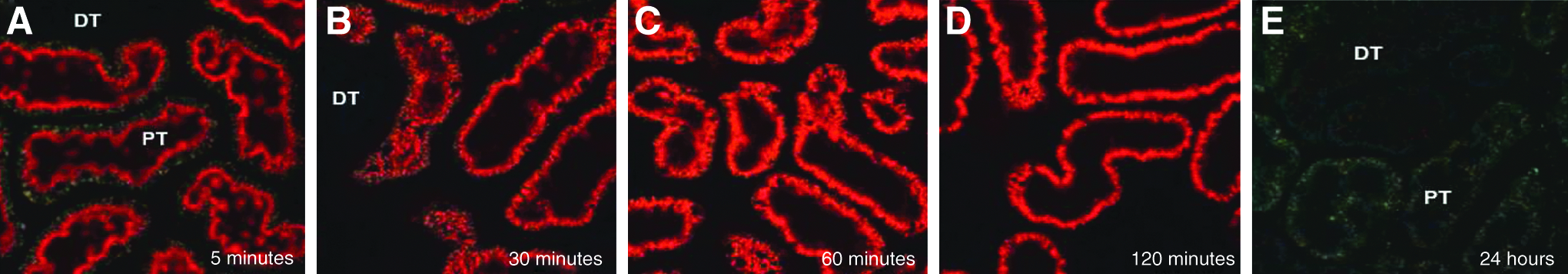
In vivo (2-photon microscopy) imaging of fluorescent-siRNA distribution in the rat kidney following i.v. administration.
These data demonstrate that I5NP is distributed rapidly to the kidneys via glomerular filtration following i.v. administration, and then is actively reabsorbed from the tubular lumen by the proximal tubular epithelial cells.
Tissue distribution in renally impaired rats
I5NP was administered to rats that underwent a 5/6 nephrectomy to examine the effect of renal impairment on I5NP tissue distribution (Table 3 and Fig. 5). Nephrectomy entailed a surgical removal of 2/3 of one kidney, followed by removal of the remaining kidney. The mean I5NP concentration in residual kidney tissue of nephrectomized animals at 3 hours after the last dose was slightly greater than the mean for normal animals on a ng/g of tissue basis (Fig. 5A), but the total percent deposition in kidneys was substantially decreased in nephrectomized animals due to the reduction in kidney mass. However, this reduced total deposition of I5NP in the kidneys of nephrectomized animals did not result in appreciably greater uptake into other organs. This was reflected by the marginally higher means for other tissues in nephrectomized versus normal animals at 3 hours following the fourth dose, all of which were much lower than the I5NP concentrations measured in kidney at this time. Most likely, the bulk of the administered dose of I5NP was excreted via the kidneys in both nephrectomized and non-nephrectomized animals, and significant redistribution to other tissues apparently did not occur as a result of partial nephrectomy. By 30 hours after the fourth dose, kidney levels in both I5NP-treated groups had declined to very low levels, with means of 12 and 55 ng/g for normal and 5/6-nephrectomized groups respectively (Fig. 5B) reflecting extensive clearance of I5NP from kidney tissue.
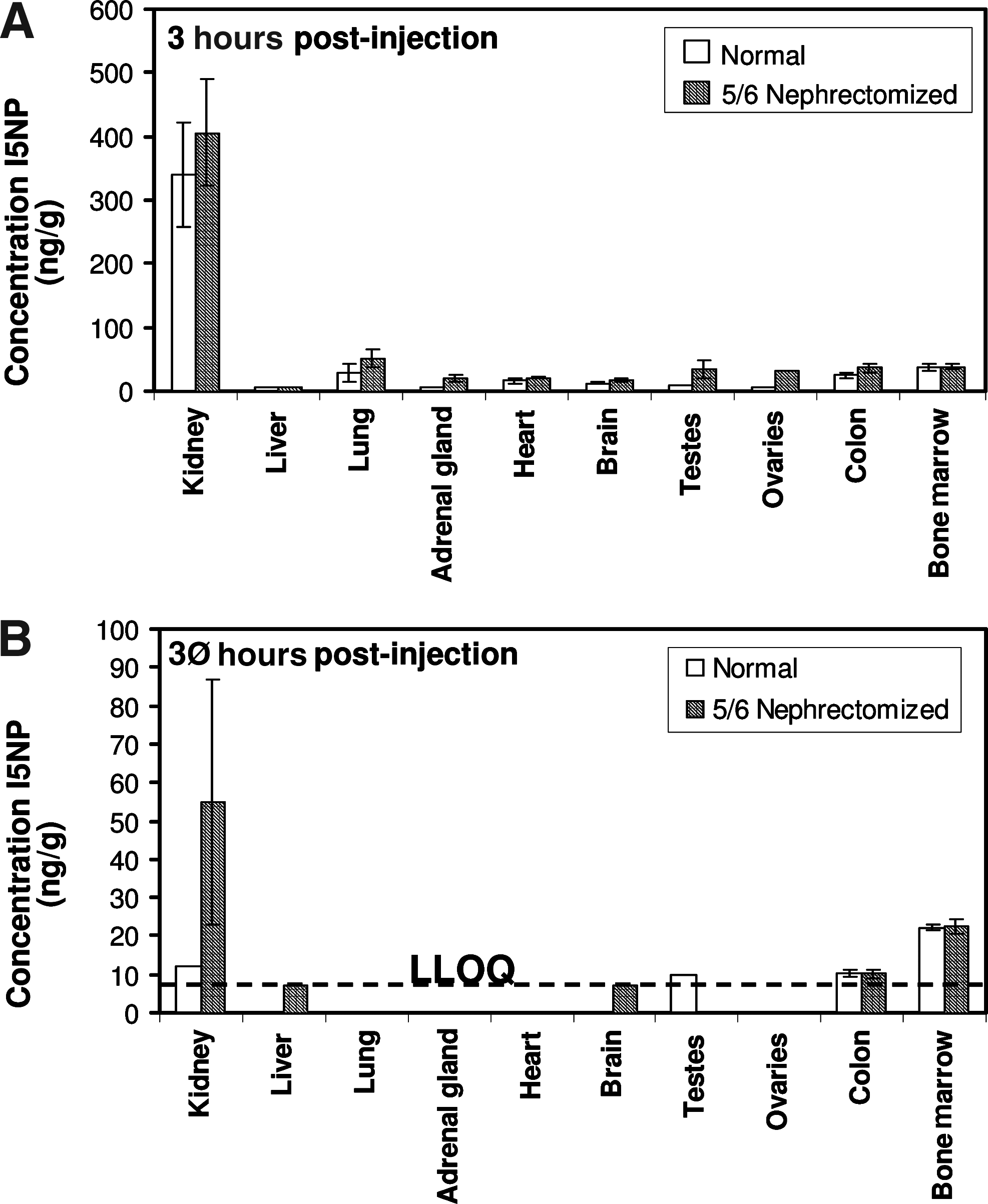
Tissue levels of I5NP after i.v. injection in normal and partially (5/6) nephrectomized rats.
LLOQ=lower limit of quantification (7 ng/g or 2 ng/g for kidney only).
Group means in data sets containing values below the limit of quantification were calculated by assuming a value identical to the LLOQ, unless more than half of the samples for males or females were less than the LLOQ, in which case the group mean was not calculated (NC).
The LLOQ for assays of some samples was set at 20 ng/g.
Results from a rejected assay; insufficient sample was available to repeat analyses; values are provided for information purposes.
Toxicology studies
The nonclinical toxicology program for I5NP was designed to support the safety of i.v. administration of I5NP. Subacute repeat-dose general toxicity studies were conducted in rats and monkeys to enable single and multiple administrations of I5NP within a 1-day period.
Safety pharmacology studies
A safety pharmacology study was performed to determine the potential effects of I5NP on the cardiovascular, respiratory, and central nervous systems following repeated i.v. bolus administration to conscious, unrestrained cynomolgus monkeys (Table 1). There were no I5NP-related effects on cardiovascular (blood pressure, heart rate, and ECGs), respiratory, or central nervous system parameters in the safety pharmacology study, nor were there any effects on other parameters assessed in this study.
Toxicity studies
Systemic toxicity of I5NP was initially evaluated in sub-acute i.v. studies in rats and monkeys to support i.v. administration of I5NP (Table 1). Despite the relatively high doses used (60 mg/kg i.v.×4 doses in the rat and 48 mg/kg i.v.×4 doses in the monkey), dose-limiting toxicities or target organs could not be determined in these studies. Therefore, single-dose i.v. toxicity studies in rats and monkeys were performed, using the maximum feasible doses of 2,000 mg/kg in rats and 1,000 mg/kg in monkeys, to identify dose-limiting toxicities/target organs (Table 1). The genotoxic potential of I5NP was evaluated in a battery of studies consisting of in vitro and in vivo assay systems.
Possible adverse effects stemming from exaggerated pharmacologic activity were assessed in the rat studies by including an analogue of I5NP (referred to as QM5) that is active in this species (Molitoris et al., 2009). The region of the p53 mRNA to which I5NP is targeted is identical in monkeys and humans (100% sequence homology); hence, no analogue of I5NP was needed to assess exaggerated pharmacology in the monkey studies.
The toxicity studies in rats and monkeys and the genotoxicity studies were conducted in accordance with the Good Laboratory Practice Regulations (21 CFR Part 58) of the United States Food and Drug Administration.
Summary of findings in the rat
In the rat single-dose i.v. study (Study 502218, Table 1), a dose of 200 mg/kg I5NP was considered the no observable adverse effect level (NOAEL). The highest dose administered (2,000 mg/kg—the maximum feasible dose) resulted in appearance of a moribund state of the first 2 male rats immediately following dosing, and subsequent high-dose animals were dosed with 1,200 mg/kg. One female also succumbed after receiving a dose of 1,200 mg/kg. The cause of death of these animals could not be determined from macroscopic or microscopic observations. Dose-dependent erosion/ulcers of the glandular stomach were observed in animals administered 200, 800, or 1,200 mg/kg I5NP or 1,200 mg/kg QM5, and subacute inflammation and edema of the pancreas was observed in animals given 800 mg/kg or 1,200 mg/kg I5NP. The changes in the stomach mucosa were graded as minimal and were only observed in 3 of 20 animals sacrificed on Day 2 in the 200 mg/kg dose group, and were not observed at all in the subacute study where animals were dosed with up to 60 mg/kg i.v.×4 doses in a 14-hour period. In the high-dose groups of the single-dose i.v. study (1,200 mg/kg), all of the clinical and anatomic macroscopic and microscopic changes were shown to be completely or partially reversed during a 2-week recovery period.
Summary of findings in the monkey
In the monkey single-dose i.v. study (Study 502217, Table 1), a dose of 500 mg/kg was considered the NOAEL. Transient clinical signs of salivation and decreased activity were observed in the mid- and high-dose groups (500 and 1,000 mg/kg—the maximum feasible dose). Slight dose-related increases in activated partial thromboplastin time (APTT) and prothrombin time (PT) were seen 5 minutes following dosing but resolved within 2 hours. The magnitude of the prolongation in APTT was marginally clinically significant (increased approximately 80%) at the maximum feasible dose of 1,000 mg/kg. There was also a small increase in mean complement C3a split product concentration in plasma at 5 minutes post-dose in all female dose groups and in males in the 500 and 1,000 mg/kg groups. There were no clinical signs or other indications of any sequelae stemming from complement activation, which is consistent with the low levels of the C3a split product observed.
I5NP was non-genotoxic in the three genetic toxicity assays conducted, and the QM5 rat analogue of I5NP was non-genotoxic in the micronucleus assay.
Discussion
I5NP is an siRNA that acts via RNA interference to temporarily inhibit the expression of the pro-apoptotic protein, p53. I5NP is being developed for diseases of the kidney related to ischemia/reperfusion injury and is currently being evaluated for the prevention of acute kidney injury in patients undergoing major cardiovascular surgery, as well as for the prophylaxis of delayed graft function following renal transplantation.
The clearance of I5NP from rat and monkey plasma following i.v. administration appeared to be multi-compartmental, with a very fast initial clearance component (initial plasma half-life on the order of minutes) that dominated the plasma concentration versus time curve. This rapid plasma clearance is likely related to the probable low binding of I5NP to plasma proteins, thereby facilitating renal filtration (Brown et al., 1994; Crooke 1998).
The distribution of I5NP in rats exhibited a predictable pattern based on the known propensity of oligonucleotides to accumulate in the kidneys (Geary et al., 1997b; Parry et al., 2000; van de Water et al., 2006). The concentration of I5NP in the kidneys of rats following i.v. injection was far greater than in other tissues, which is consistent with the clinical objective of delivering this anti-p53 siRNA to the kidneys in patients following renal injury. Two-photon microscopy data indicate that uptake in the kidney occurs rapidly via reabsorption of the siRNA from the glomerular filtrate selectively by proximal tubule epithelium (Molitoris et al., 2009). In these studies, the residence time of siRNA in proximal tubule cells was less than 24 hours. The rapid uptake and short residence time in proximal tubule epithelium as measured by 2-photon microscopy is in good agreement with the duration of the pharmacodynamic activity of p53 mRNA knockdown observed in rat kidney following i.v. injection of the rat analogue, where reduction in p53 mRNA could be detected quickly (within 3–-6 hours) after administration which persisted for less than 48 hours (Molitoris et al., 2009).
Renal impairment had little impact on the overall plasma clearance profile, as concentrations of I5NP still diminished quite rapidly in partially (5/6) nephrectomized rats and were vanishingly low by 2 hours post-injection. There was also no clear shift in tissue distribution associated with partial nephrectomy in the rat. These results are in good agreement with data obtained with a phosphorothioate-modified oligonucleotide in rats with either tubular (cisplatin-induced) or glomerular (anti-Thy1.1 antibody-induced) damage, where urinary elimination and tissue deposition were not markedly altered compared to normal animals (Masarjian et al., 2004). However, in quantitative terms, the tissue deposition of I5NP was substantially lower than that reported for phosphorothioate-modified oligonucleotides, particular for non-target tissues (i.e., those other than kidney). Peak tissue levels of I5NP suggest that only 1%–2% of the administered dose was absorbed by tissues, with very little uptake in extra-renal tissues, and the rest likely being eliminated in the urine. In contrast, up to 40% of the administered dose of phosphorothioate-modified oligonucleotides was reportedly taken up by the kidney, liver, and spleen following i.v. administration (Geary et al., 1997a; Phillips et al., 1997; Levin et al., 1998; Monteith and Levin, 1999). This difference in tissue distribution probably derives from the relatively low protein binding of phosphodiester-backbone oligonucleotides (an siRNA, in this case), versus phosphorothioate-modified antisense molecules (Brown et al., 1994; Crooke 1998). Greater uptake by extra-renal tissues has been reported for synthetic RNAs via other parenteral routes of administration, such as subcutaneous injection (Sandberg et al., 1999; Lee et al., 2000; Parry et al., 2000).
The NOAEL for I5NP in the rat was 200 mg/kg. Clinical signs in the rat, which resolved the same day, primarily consisted of decreased activity, labored breathing, and altered breathing sounds, decreased muscle tone, and pallor/blue skin. Erosion and/or ulcers of the glandular stomach and subacute inflammation and edema of the pancreas were identified as the primary treatment-related tissue pathologies in rats. Thymic hemorrhages were also observed in the rat at doses of 800 and 1,200 mg/kg but were of uncertain toxicologic significance. All of the clinical and anatomic pathology changes in the rat were shown to be completely or partially reversible during a 2-week recovery period. Similar findings were observed with the rat analogue of I5NP, indicating no additional effects associated with RNAi-mediated down-regulation of p53 protein expression. It is surprising that no histopathologic changes were observed in the organ exhibiting the greatest uptake (i.e., the kidney). It is possible that the hemorrhaging observed in the extra-renal tissues may be due to the viscosity of the test article, leading to micro-emboli, since high concentrations (150 mg/mL of siRNA) and large dosing volumes (up to 20 mL/kg) of the siRNA were required to achieve the high dose levels evaluated in these studies.
The NOAEL in the monkey was 500 mg/kg. No effects on cardiovascular, respiratory, and neurologic parameters were observed in monkeys following i.v. administration at dose levels up to 25 mg/kg. Clinical signs in the monkey, which resolved the same day, primarily consisted of salivation and decreased activity. Slight dose-related increases in APTT and PT were seen 5 minutes following dosing but resolved within 2 hours. The magnitude of the prolongation in APTT was marginally clinically significant (increased approximately 80%) only at the highest dose tested of 1,000 mg/kg. There was a small increase in mean complement C3a split product concentration in plasma at 5 minutes post-dose in females that received 200 mg/kg and higher, and in males at doses of 500 mg/kg and higher. There were no clinical signs or other indications of any sequelae stemming from complement activation, which is consistent with the low levels of the C3a split product. All other parameters measured were unaffected by treatment with I5NP, and the acute changes in coagulation and complement parameters were no longer present during or at the end of a 15-day recovery period.
The effects on complement and coagulation are consistent with the known blood-level-dependent class effects of phosphorothioate-modified oligonucleotides (PS ONs), although the dose levels required to elicit such changes with I5NP were much greater than those reported for PS ONs (Henry et al., 1997). An extensive body of published information, primarily derived from monkey studies, has established that phosphorothioate-modified oligonucleotides can prolong coagulation times, activate the alternate complement pathway, accumulate in organs rich in macrophages, primarily the liver and spleen, and nonspecifically activate the innate immune system, resulting in liver, spleen and lymph node enlargement (largely attributable to an increase in the resident macrophage and or lymphocyte population, as well as mononuclear infiltration in these organs). These effects are due to the phosphorothioate backbone and are not sequence-specific, although they can be tempered by the introduction of secondary chemical modifications in the ribose units (Henry et al., 2000). Of these various class effects observed with PS ONs, only the blood-level-dependent effects (i.e., minor prolongation of APTT and activation of complement) were observed with I5NP, and only at very high dose levels in monkeys (500 and 1,000 mg/kg), presumably because of the absence of a phosphorothioate backbone.
In the monkey studies reported herein, we observed no increase in the Bb fragment and only a modest increase of ≤3-fold in C3a. This can be compared to a 25-fold increase in Bb and a similar 3-fold increase in C3a caused by first-generation phosphorothioate antisense oligonucleotides (Henry et al., 1997). The plasma levels of phosphorothioate that resulted in the 25-fold increase in Bb were 146 μg/mL. These levels are approximately 10- and 60-fold lower than the plasma concentrations produced by the lowest (200 mg/kg) and highest (1,000 mg/kg) doses, respectively, of I5NP given to monkeys (Table 1). The lower range of C3a elevation (≤3-fold) observed with 500 and 1,000 mg/kg of I5NP can reportedly be produced by a dose of PS ON as low as 20 mg/kg (i.e., 25- and 50-fold lower). Hence, the propensity for I5NP to elicit complement activation is remarkably diminished, relative to the dose-response for PS ONs. Moreover, the response to I5NP appears quantitatively different than that produced by PS ONs, as the latter class of molecules primarily elicits alternative pathway activation (reflected by elevations in plasma levels of the Bb split product), whereas no increase in Bb was observed in monkeys treated with very large doses of I5NP. Thus, it may be surmised that the limited complement activation produced by I5NP was secondary to stress, as it has been shown that the complement system can be activated by stress (Burns et al., 2008).
Overall, I5NP was well tolerated in rodents and nonhuman primates. The favorable safety profile of I5NP relative to that of other published oligonucleotides likely relates to the reduced distribution to extra-renal tissues, and lack of non-natural modifications such as phosphorothioate backbone substitutions typically introduced into single-stranded oligonucleotides to confer resistance to nuclease degradation. Taken together, these data support clinical testing of i.v. administration of I5NP for the preservation of renal function following acute kidney injury and renal transplantation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.