
Abstract
Stromal cell-derived factor-2 (SDF-2) is reportedly involved in multiple endoplasmic reticulum (ER) functions, including the misfolded protein catabolic process, protein glycosylation, and ER protein quality control. However, the precise molecular and cellular functions of SDF-2 remain unknown. Previously, we discovered that SDF-2 mediates acquired resistance to oxaliplatin in human gastric cancer cells. In this study, we have generated SDF-2-specific monoclonal antibodies (mAbs), using the rat medial iliac lymph node method, as a tool to explore novel mechanisms of oxaliplatin resistance. The antibodies detected endogenous human SDF-2 in immunoblotting analyses. In addition, immunoprecipitation analyses revealed the availability of these antibodies for human SDF-2. Thus, these mAbs will be available to elucidate molecular and cellular functions of SDF-2 in cancer cells.
Introduction
Stromal cell-derived factor-2 (SDF-2) was first isolated and cloned as complementary DNA (cDNA) encoding a novel secretory protein from the endoplasmic reticulum (ER) and Golgi apparatus of a mouse stromal cell line, ST-2.(1) The human SDF2 gene maps on chromosome 17 at q11.2 and encodes a protein of 211 amino acids, including three MIR (protein O-mannosyltransferase [PMT], inositol 1,4,5-trisphosphate receptor [IP3R], and ryanodine receptor [RyR]) domains.(1–3)
SDF-2 also contains an N-terminal hydrophobic signal sequence and a C-terminal ER retention-like motif, HAEL. Fujimori et al. confirmed that the HAEL motif functions as an ER retention signal; therefore, SDF-2 is localized in the ER.(4) They have also suggested that SDF-2 associates with chaperones in the ER, which contributes to preventing the aggregation of misfolded proteins.(4) In human placental cells, SDF-2 may contribute to cell survival during the unfolded protein response by interfering with ER stress proteins such as spliced XBP1 (XBP1s) and CHOP.(5)
Similarly, other studies in mice and in plants such as Arabidopsis thaliana and Oryza sativa L. have shown that SDF-2 acts as a component of the ER chaperone and is related to quality control of newly synthesized proteins.(3,6–9)
Our previous study has shown that SDF-2 mediates acquired resistance to oxaliplatin, a platinum-based chemotherapeutic agent, in the human gastric cancer cell line OCUM-2M.(10) However, since the exact molecular functions of SDF-2 are not known, it is unclear by which mechanism SDF-2 confers oxaliplatin resistance to OCUM-2M cells. Therefore, in this study, we used the rat medial iliac lymph node method to generate monoclonal antibodies (mAbs) against human SDF-2. The generated SDF-2 mAbs will be useful tools in the immunoblotting and immunoprecipitation of human SDF-2 proteins in cancer cells.
Materials and Methods
Cell culture
The human gastric cancer cell line, OCUM-2M, and the oxaliplatin-resistant subcell line, OCUM-2M/OXA, were cultured in Dulbecco's modified Eagle's medium (DMEM) containing high glucose (Fujifilm Wako Pure Chemical Corporation, Osaka, Japan) and supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 0.1 mg/mL streptomycin (Meiji Seika Pharma Co., Tokyo, Japan) in a humidified 5% CO2 incubator at 37°C. The cell lines were derived from a diffuse-type human gastric cancer patient, and the subline was established from parental cells by stepwise exposure to oxaliplatin.(11)
Construction of SDF-2-knockout cells by CRISPR/Cas9
To generate SDF2-knockout OCUM-2M/OXA cells, cells were cotransfected with the human codon-optimized SpCas9 and chimeric guide RNA (gRNA) expressing pX330 vector (pX330-U6-Chimeric_BB-CBh-hSpCas9, No. 42230; Addgene) targeting 5′-ggggggtttgtggagcgctgtgg-3′ in the human SDF2 coding sequence and pX459 vector expressing SpCas9 and puromycin resistance gene (pSpCas9[BB]-2A-Puro [PX459] V2.0, No. 62988; Addgene) using Lipofectamine LTX reagent (Thermo Fisher Scientific, Waltham, MA). Transfected cells were subjected to puromycin selection for 96 hours and then cloned by limiting dilution to obtain single cell clones. Individual clones were validated by sequencing with polymerase chain reaction-amplified fragments for frameshift indel mutations and by immunoblotting analysis with an SDF-2-specific antibody (Santa Cruz Biotechnology, Dallas, TX).
Rat immunization and mAb production
Anti-SDF-2 rat mAbs were generated using the rat lymph node method established by Sado et al.(12) A 9-week-old female WKY/Izm rat (Japan SLC, Hamamatsu, Japan) was injected in the hind footpads with 200 μL of an emulsion containing 266 μg of a synthetic peptide of 20 amino acids (GIFMKPSELLKAEAHHAELC), which corresponded to residues 193–211 of human SDF-2, and complete Freund's adjuvant. After 19 days, the cells from the medial iliac lymph nodes from the immunized rat were harvested and then fused with mouse myeloma SP2 cells at a ratio of 5:1 in a 50% polyethylene glycol (PEG 1500; Roche, Basel, Switzerland) solution. The resulting hybridoma cells were plated into 96-well plates and cultured in HAT selection medium (Hybridoma-SFM [Thermo Fisher Scientific]; 10% FBS; 5% BM-condimed H1 [Roche]; 100 mM hypoxanthine; 0.4 mM aminopterin; and16 mM thymidine). Eight days postfusion, the hybridoma supernatants were screened using an enzyme-linked immunosorbent assay (ELISA) against the human SDF-2 synthetic peptide. Positive clones were subcloned and rescreened by ELISA, immunoblotting, and immunoprecipitation.
Enzyme-linked immunosorbent assay
The synthetic human SDF-2 peptide (0.92 μg/mL) was diluted in ELISA buffer (20 mM sodium phosphate, pH 7.2) and adsorbed on the surface of Serocluster 96-well Nunc-Immuno™ Plate II (Thermo Fisher Scientific) by incubating overnight at 4°C. The plates were then blocked with 1% bovine serum albumin in TBS-T to avoid nonspecific binding. Next, the hybridoma supernatants were added and the plates were again incubated overnight at 4°C. After washing three times with TBS-T, the plates were incubated for 30 minutes at room temperature with horseradish peroxidase (HRP)-conjugated anti-rat IgG antibody (GE Healthcare, Buckinghamshire, United Kingdom) at a dilution of 1:5000. After washing with TBS-T, immunoreactivity was visualized using a pNPP phosphatase substrate system (KPL, Gaithersburg, MD), and the absorbance was measured at 450 nm using the Varioskan™ LUX (Thermo Fisher Scientific).
Viral infection
Lentiviral packaging was performed by cotransfection of pLenti6-PUbc-mSlc7a1-HygR (No. 17224; Addgene), HIV-based vectors including a cDNA expression cassette, and the pPACK packaging plasmids (System Biosciences, Palo Alto, CA) as previously described.(13) Plasmid DNAs were transfected into 293T cells by the calcium phosphate transfection method. At 32 hours post-transfection, the culture supernatants were harvested and stored as lentivirus stock. OCUM-2M cells were plated at a density of 1 × 105 cells/well in 12-well plates, and then infected with fivefold diluted lentiviral stock solution. After 24 hours, the virus solution was replaced with fresh culture media. OCUM-2M/mSlc7a1 cells were then selected by 800 μg/mL hygromycin (Fujifilm Wako Pure Chemical Corporation) for 5 days. Plat-E cells were transfected with SDF2-FLAG/pMXs-Puro (RTV-012; Cell Biolabs, San Diego, CA) by the PEI (PolyScience, Niles, IL) transfection method for retrovirus production. At 32 hours post-transfection, the culture supernatants were harvested and stored as retrovirus stock. OCUM-2M/mSlc7a1 cells were plated at a density of 1 × 105 cells/well in 12-well plates, followed by infection with fivefold diluted retroviral stock solution. OCUM-2M/SDF-2-FLAG cells were selected by 1 μg/mL puromycin (Fujifilm Wako Pure Chemical Corporation) for 5 days.
Preparation of the microsomal fraction
OCUM-2M/SDF-2-FLAG cells were homogenized with 0.25 M sucrose buffer containing 5 mM Tris-HCl (pH 7.5) and 1 mM EDTA in a Potter-type homogenizer. The homogenate was centrifuged at 1,000 × g for 15 minutes at 4°C to remove the nuclei and cellular debris. The supernatant was further centrifuged at 12,000 × g for 10 minutes, and followed by ultracentrifugation using the SW 41 rotor in the Optima™ L-90K ultracentrifuge (Beckman Coulter, Inc., Pasadena, CA) at 150,000 × g for 20 minutes at 4°C to collect the microsomes. The microsomal pellets were lysed with lysis buffer containing 50 mM HEPES-NaOH (pH 7.5), 150 mM NaCl, 1% NP-40, 5 mM EDTA, and protease inhibitor cocktail (Nacalai Tesque, Kyoto, Japan).
Immunoprecipitation
The microsomal fraction of OCUM-2M/SDF-2-FLAG cells was lysed in immunoprecipitation buffer containing 50 mM HEPES-NaOH (pH 7.5), 150 mM NaCl, 0.5% NP-40, 5 mM EDTA, and protease inhibitor cocktail (Nacalai Tesque) for 15 minutes on ice. The lysates were incubated with recombinant protein G agarose (Thermo Fisher Scientific) for 2 hours at room temperature to preclear any nonspecific binding. The precleared lysates were then subjected to immunoprecipitation with rat anti-SDF-2 mAbs and incubated overnight at 4°C. The precipitates were washed three times with immunoprecipitation buffer, followed by elution with sodium dodecyl sulfate (SDS) sample buffer for 5 minutes at 95°C. The eluants were subjected to immunoblot analysis.
Immunoblotting
The microsomal fraction of OCUM-2M cells was washed twice with phosphate-buffered saline and lysed in lysis buffer containing 50 mM HEPES-NaOH (pH 7.5), 150 mM NaCl, 1% NP-40, 5 mM EDTA, and protease inhibitor cocktail (Nacalai Tesque). The samples were subjected to sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) using 12.5% acrylamide gels and transferred to Immobilon-P transfer membrane (Merck KGaA, Darmstadt, Germany). The membranes were blocked for 1 hour at room temperature with 1% skim milk in TBS-T (0.1% Tween-20), and then incubated overnight at 4°C with rat anti-SDF-2 mAbs diluted to 1:1,000 in Can Get Signal Immunoreaction Enhancer Solution 1 (Toyobo, Osaka, Japan). After washing in TBS-T, the membranes were incubated for 1 hour at room temperature with goat anti-rat IgG-HRP (GE Healthcare) diluted to 1:10,000 in Can Get Signal Immunoreaction Enhancer Solution 2 (Toyobo). After washing in TBS-T, chemiluminescence detection was performed with Immobilon ECL Ultra Western HRP Substrate or Immobilon Western HRP Substrate (Merck KGaA) and visualized directly using a ChemiDoc XRS+ system (Bio-Rad Laboratories, Berkeley, CA). Additional antibodies were purchased from the following sources: anti-calreticulin antibody from Cell Signaling Technology (Danvers, MA); anti-FLAG-HRP antibody from Merck KGaA; anti-SDF-2 antibody from Santa Cruz Biotechnology; and anti-rabbit IgG HRP linked F(ab′)2 fragment donkey from GE Healthcare. The membranes were stripped in boiled Milli-Q water and then stained with Quick-CBB (Fujifilm Wako Pure Chemical Corporation) for visualizing amounts of loaded proteins.
Results and Discussion
The full-length human SDF-2 protein is a 211-amino acid sequence. To generate an mAb specific for the human SDF-2 protein, we utilized a peptide of 20 amino acids, which corresponded to residues 193–211 of human SDF-2 as an antigen. Following the rat lymph node method for generating antibodies, lymphocytes were collected from the enlarged lymph nodes of the immunized rats 19 days postimmunization. The hybridomas, which were obtained after fusing the lymphocytes with mouse myeloma SP2 cells, were tested for the production of mAbs that react with the synthetic peptide antigen by ELISA. The positive clones were examined by immunoblotting to evaluate their reactivity against human SDF-2 in total extracts of PANC-1 cells. One clone, designated as mAb 3A1, reacted with a 23 kDa protein band. We therefore subcloned and rescreened by ELISA to validate the specificity of the mAb for SDF-2 using SDF2-knockout cells. Because SDF-2 is localized in the ER, we performed immunoblotting for SDF-2 using the microsomes, which are heterogeneous vesicle-like fragments formed from pieces of the ER of SDF2-knockout OCUM-2M/OXA cells. All the clones were purified using protein G affinity resins. Using a rat isotyping kit, the immunoglobulin class of both mAbs, 3A1-1D6 and 3A1-1H10, was determined to be rat IgG2ακ. Both clones gave specific signals as a 23 kDa band in immunoblotting (Fig. 1). Commercially available anti-SDF-2 antibody, sc-165443, also gave the same signal. We next examined the availability of both clones in immunoprecipitation analysis. SDF-2 and FLAG were detected slightly lower than the rat IgG light chains in both precipitants of the 3A1 clone, whereas subclones of 3B5 (negative clones) and control rat IgG did not; this indicated that the 3A1 clone efficiently immunoprecipitated the human SDF-2-FLAG fusion proteins (Fig. 2). From these findings, we concluded that mAb 3A1 specifically recognized endogenous human SDF-2. This novel tool will be beneficial for immunoblotting and immunoprecipitation. The use of this antibody will allow for further elucidation of the molecular and cellular functions of SDF-2 in the ER functions of cancer cells.
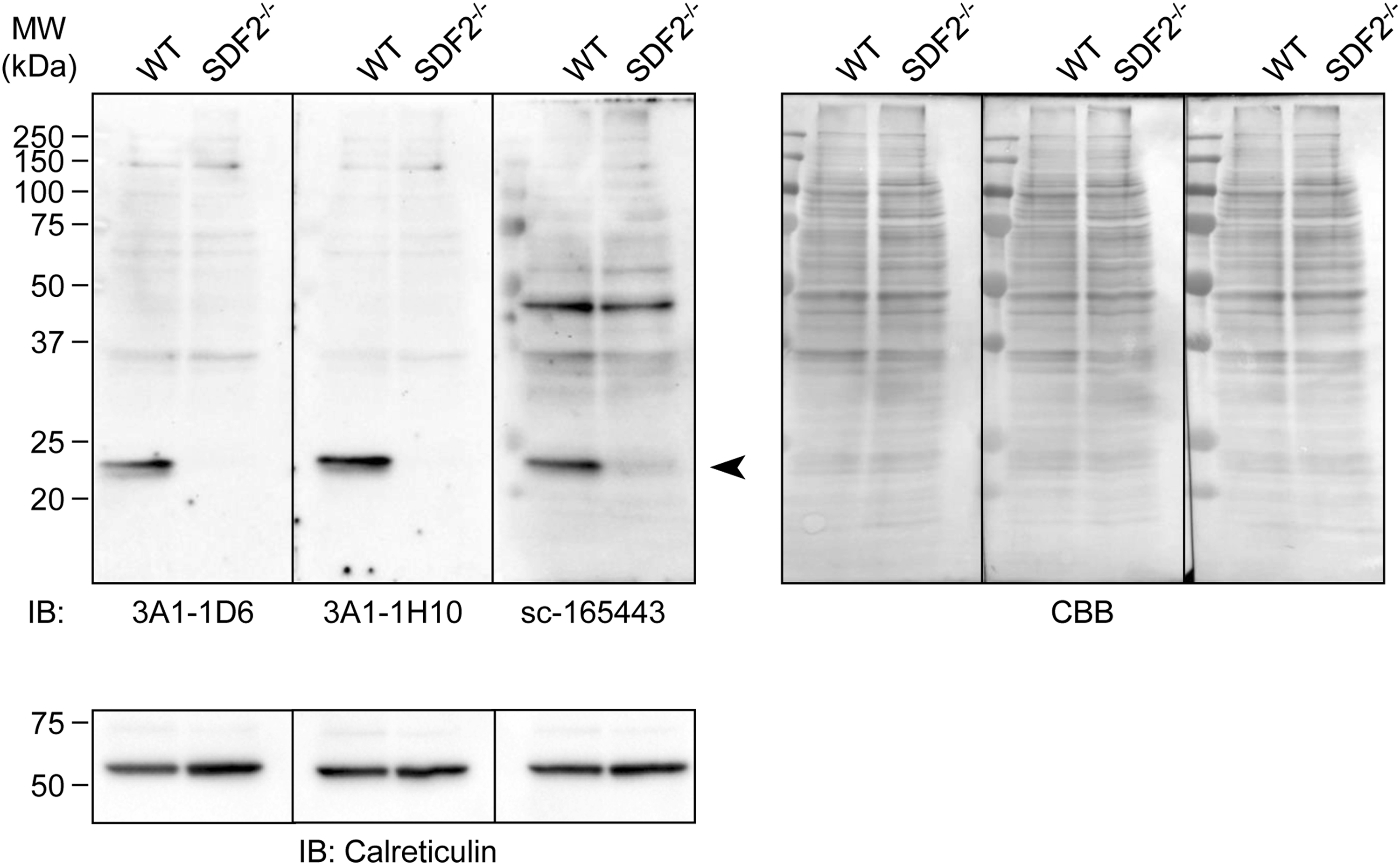
Specificity of anti-SDF-2 mAbs. Immunoblotting analysis of the microsomal fraction from WT or SDF2-knockout (SDF2−/−) OCUM-2M/OXA cells, using anti-SDF-2 antibodies. Commercially available anti-SDF-2 antibody, sc-165443, was used as a positive control, and blots were reprobed with an anti-calreticulin antibody as a loading control. All blots were striped and restained with CBB R-250 staining to visualize the amounts of loaded proteins. Arrowhead, SDF-2. CBB, coomassie brilliant blue; mAbs, monoclonal antibodies; SDF-2, stromal cell-derived factor-2; WT, wild type.
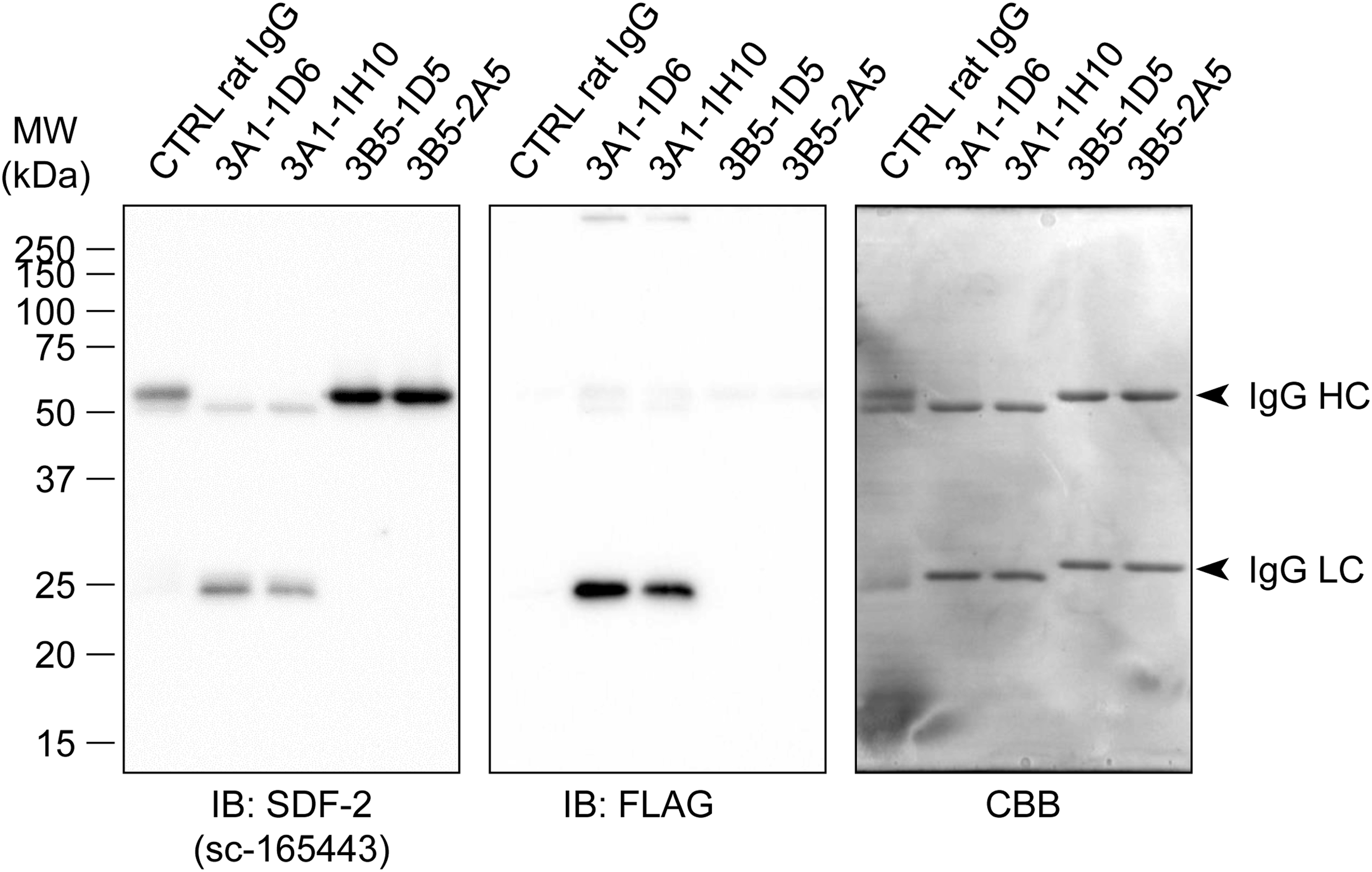
Immunoprecipitation using anti-SDF-2 mAbs with the microsomal fraction from SDF-2-FLAG expressing OCUM-2M cells. Control rat IgG and negative clones of generated mAbs (3B5-1D5 and 3B5-2A5) were used as negative controls. All blots were striped and restained with CBB staining to visualize the amounts of loaded proteins. Arrowheads, IgG heavy and light chains.
Footnotes
Acknowledgment
The authors thank the Research Support Platform of Osaka City University Graduate School of Medicine for technical support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was partly supported by JSPS KAKENHI Grant, number 17K15757, and the Uehara Memorial Foundation.