
Abstract
CD28 superagonist (CD28SA), a therapeutic immunomodulatory monoclonal antibody triggered rapid and exaggerated activation of CD4+ effector memory T cells (TEMs) in humans with unwanted serious adverse effects. It is well known that distinct metabolic programs determine the fate and responses of immune cells. In this study, we show that human CD4+ TEMs stimulated with CD28SA adopt a metabolic program similar to those of tumor cells with enhanced glucose utilization, lipid biosynthesis, and proliferation in hypoxic conditions. Identification of metabolic profiles underlying hyperactive T cell activation would provide a platform to test safety of immunostimulatory antibodies.
Introduction
CD28 superagonists (CD28SAs) are monoclonal antibodies (mAbs) that engage CD28 costimulatory receptors resulting in potent T cell activation independent of concomitant T cell receptor (TCR) engagement.(1) TGN1412, a CD28SA, triggered an excessive and adverse cytokine release (cytokine storm) when administered to volunteers in a human Phase 1 clinical trial.(2) We and others have shown that CD28SA induces exaggerated activation, polyclonal expansion, and migration of CD4+ effector memory T cells (TEMs).(2–4) The basis for this hyperactive, dysregulated phenotype is subject of much research and the lack of inhibitory inputs(4) has been suggested as one potential mechanism. T cells need to undergo metabolic reprogramming to adapt to their changing metabolic needs as they progress from resting T cells to fully differentiated effectors and memory cells.(5,6)
T cells utilize glucose as the primary nutrient source for energy generation and biomass production, although amino acids, such as glutamine, are also utilized.(5) Before activation, naive T cells have low metabolic requirements and rely on mitochondrial oxidative pathways for basal energy generation. Upon activation, T cells adopt a metabolic profile typified by aerobic glycolysis and basal oxidative phosphorylation (OXPHOS).(5,7) Rapidly proliferating T cells require lipids to support membrane biogenesis and depending on the T cell subset, lipids may be acquired or synthesized (lipogenesis).(8) T cells use distinct metabolic programs according to their differentiation state and immunological role. Studies have shown that CD4+ T helper (Th)1, Th2, and Th17 cells are highly glycolytic, whereas CD4+ regulatory T cells have high lipid oxidation rates.(9) The metabolic phenotype of the TGN1412-target cell, CD4+ TEMs, is yet to be fully characterized. Importantly, the metabolic program that supports the dramatic hyperactivation and proliferation induced by CD28SA is currently unknown.
Tumor cells exhibit metabolic abnormalities such as elevated aerobic glycolysis and de novo fatty acid biosynthesis to generate the energy required to support rapid cell division.(10) In this report, we demonstrate that superagonistic activation programs CD4+ TEMs toward a tumor cell-like metabolic profile that favors enhanced glycolysis and lipogenesis. We also define ATP-citrate lyase (ACL) and acetyl-Coenzyme A (ACC) as key molecular indicators of the CD28SA-induced lipogenic phenotype.
Materials and Methods
Reagents
All reagents were obtained from Sigma-Aldrich (United Kingdom) unless otherwise stated.
Effector memory T cell isolation
Ethics approval for the use of human peripheral blood mononuclear cells (PBMCs) from healthy donors was given by the local Ethics Committee and all subjects provided informed consent. PBMCs were isolated from heparinized venous blood by density gradient separation (LymphoPrep, O7811; Axis-Shield). The CD4+ TEMs isolation kit (130–094–125; Miltenyi Biotec) was used to purify TEMs from PBMCs according to the manufacturer's instructions.
Stimulating antibodies
Humanized superagonistic anti-CD28 antibody, NIB1412, a human IgG4 sharing the H chain V region and L chain sequences of TGN1412, was generated at the National Institute for Biological Standards and Control (NIBSC, United Kingdom). Murine anti-human CD3 (clone: UCHT1, Cat No. 16–0038–85) antibody was purchased from eBioscience (United Kingdom).
Proliferation assays
Plate-bound or solid-phase PBMC in vitro systems have been previously shown to support robust T cell activation by anti-CD3 and CD28SA,(4,11) and therefore this system was chosen to study metabolic reprogramming of TEM cells. Ninety-six-well round-bottom non tissue culture treated plates were coated with stimulating antibodies at 37°C for 2 hours. Plates were washed twice to remove unbound antibody before addition of T cells. The T cells were cultured in complete media (RPMI 1640 supplemented with 15% fetal calf serum (Life Technologies, United Kingdom), 2 mM
Cell viability assay
Briefly, CD4+ TEMs were plated in base media with
Flow cytometric analysis
CD4+ TEMs were activated with plate-bound anti-CD3 or NIB1412 for 48 hours. For the quantification of mitochondria, cells were stained with MitoTracker® Deep Red FM (M22426; Molecular Probes) at 20 nM during the last 30 minutes of treatment. Cells were washed with phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde and stained with HCS LipidTOX™ Green Neutral Lipid Stain (H34475; Invitrogen) at 1:500. To quantify mitochondrial superoxide production, cells were incubated with MitoSOX™ Red (Cat No. M36008; Invitrogen) at 37°C for 10 minutes, washed and fixed in 2% paraformaldehyde. MitoSOX Red was excited at 488 nm and fluorescence emission at 575 nm was measured.
For the determination of glucose uptake and cell surface expression of glucose transporters, cells were incubated with 2-NBDG (N13195; Molecular Probes) for 30 minutes, washed three times, and then stained with anti-Glut1-PE (MAB1418; R&D Systems) for 20 minutes. Cells were then washed and fixed with 4% paraformaldehyde.
Staining and incubations were performed at 37°C. Untreated cells were used as controls. Fluorescent signals from cells were acquired on BD FACS Canto II flow cytometer and data were analyzed using Cyflogic software v. 1.2.1.
Immunofluorescence microscopy
Imaging of mitochondria and lipid droplets was performed by washing preactivated cells after 48 hours and plating them on poly-
Gel electrophoresis and western immunoblotting
Cells were lysed with RIPA buffer with 1 mM PMSF and protease inhibitor cocktail. Twenty micrograms of protein lysate was resolved by 10%–12% SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis), transferred to PVDF membranes (Bio-Rad), blocked, and probed with the primary antibodies: anti-ACL (phospho S455) (#4331; CST) and anti-Acetyl Coenzyme A Carboxylase (phospho S79) (ab68191; Abcam, United Kingdom) followed by appropriate horseradish peroxidase-conjugated secondary antibodies (Cell Signaling Technology, United Kingdom) and visualized using the ECL system (PerkinElmer Life Sciences).
Bioenergetics of T cells
CD4+ TEMs were stimulated with plate-bound anti-CD3 mAb or NIB1412 for 48 hours in complete media and then resuspended in serum-free base media (102353-100; Seahorse Bioscience) with 2 mM
Acetyl-Coenzyme A measurement
Concentration of acetyl-CoA in whole-cell lysates from preactivated CD4+ TEMs was measured by fluorescence assay using a kit according to the manufacturer's instructions (The PicoProbe Acetyl CoA Assay Kit: ab87546; Abcam). Fluorescence was quantified using a Varioskan™ Flash Multimode Plate Reader (Thermo Scientific, United Kingdom).
Citrate and alpha-ketoglutarate measurement
Concentration of citrate and alpha-ketoglutarate (α-KG) in whole-cell lysates from preactivated CD4+ TEMs was measured by fluorescence assay using a kit according to the manufacturer's instructions (The Citrate Assay Kit: ab83396, α-KG Assay Kit: ab83431; Abcam). Relative fluorescence units/intensity were quantified using a Varioskan Flash Multimode Plate Reader (Thermo Scientific).
Statistical analysis
Unpaired, two-tailed Student's t-test was used to analyze data and results were presented as mean ± standard deviation. p < 0.05 was considered to be statistically significant.
Results
CD28SA-activated TEMs cells display a hyperactive phenotype
T cell activation triggers metabolic programs that promote biomass generation, which is evident by an increase in cell size, termed blasting. We used a TGN1412-like CD28SA mAb, termed NIB1412 to stimulate CD4+ TEMs and demonstrate that the percentage of blasting T cells in the CD28SA-stimulated condition is about 4-fold (59% compared to 14% with anti-CD3) greater than in the CD3-activated condition (Fig. 1A). We also confirmed the hyperactive phenotype of CD28SA-activated T cells as evidenced by their enhanced proliferative response (Fig. 1B).
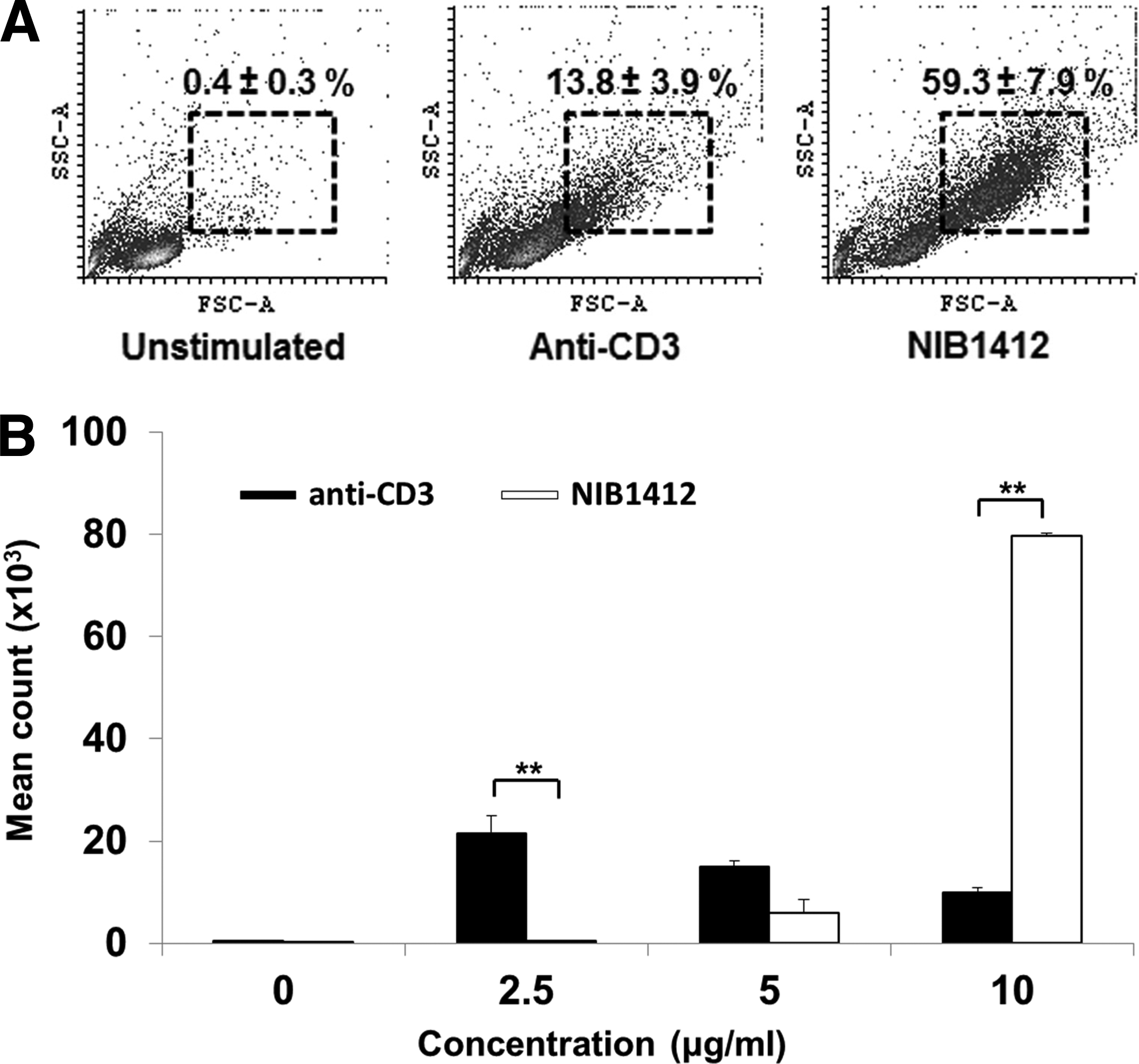
CD28SA-activated CD4+ TEMs display a hyperactive phenotype.
CD28SA stimulation maximizes OXPHOS potential
To assess the contribution of OXPHOS to meet the energy requirements during hyperactivation of CD4+ TEMs upon CD28SA stimulation, we used extracellular flux analysis to measure oxygen consumption rates (OCR), basal respiration, ATP production, maximal respiration and spare respiratory capacity (SRC). CD28SA-stimulated T cells had higher basal respiration and ATP production when compared with anti-CD3-activated CD4+ TEMs (Fig. 2A). To determine whether CD28SA-stimulated T cells had more active mitochondrial mass, we stained CD4+ TEMs with MitoTracker Deep Red and found that CD28SA stimulation led to a higher quantity of actively respiring mitochondria than when activated with anti-CD3 mAbs (Fig. 2B, C). CD28SA-stimulated T cells also produced more mitochondrial reactive oxygen species (mitoROS) than anti-CD3-activated CD4+ TEMs (Fig. 2D), which is reflective of enhanced mitochondrial electron transport activity. Collectively, these results indicate that CD28SA-activated cells maximize their OXPHOS potential, which is supported by their greater active mitochondrial mass. To test whether CD28SA-stimulated cells are reliant on OXPHOS for their hyperproliferation, we cultured activated CD4+ TEMs under hypoxic or normoxic conditions. While, CD3-stimulated cells proliferated more in normoxic than in hypoxic condition, CD28SA-stimulated cells proliferated in both the conditions with higher rate of proliferation in hypoxic condition (Fig. 2E). This suggests that CD28SA-activated T cells have a flexible metabolic program that reduces the reliance on OXPHOS for proliferation.
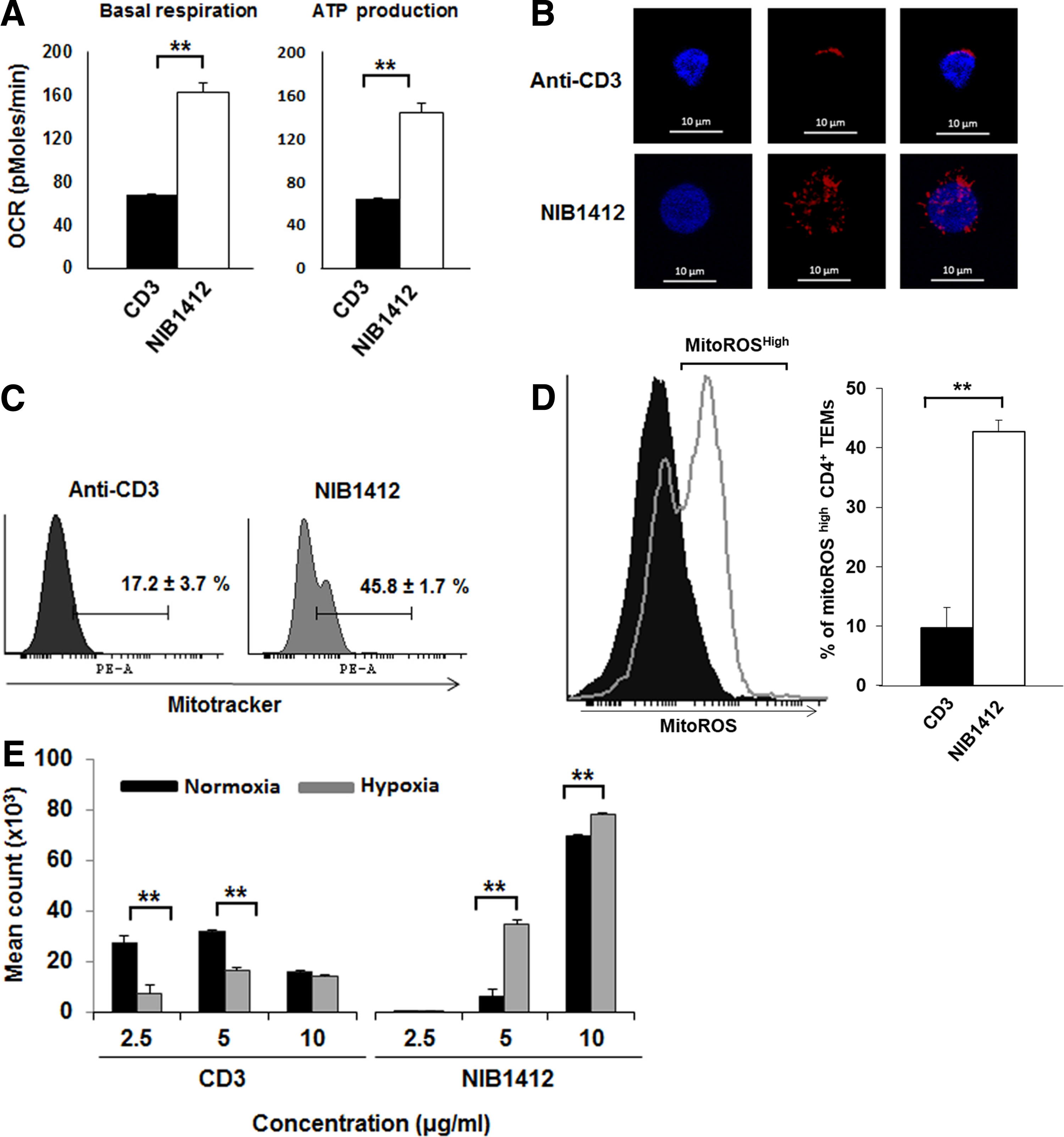
CD28SA stimulation maximizes OXPHOS potential.
CD28SA stimulation maximizes glucose utilization
To assess glucose handling during CD28SA stimulation, we measured ECAR, a marker of glycolysis, and found that CD28SA-stimulated CD4+ TEMs displayed significantly higher glycolysis and lacked glycolytic reserve compared with anti-CD3-stimulated cells (Fig. 3A). This result indicates that CD28SA-activated cells adopt a metabolic program that maximizes basal glycolytic rate. We tested whether the high glycolytic rate was associated with altered glycolytic machinery in CD4+ TEMs. The enzyme GAPDH is a critical component of the glycolytic pathway that generates pyruvate from glucose (Supplementary Fig. S1) with cytoplasmic distribution of GAPDH being indicative of glycolytic activity. We examined the cellular distribution of GAPDH by immunofluorescence and observed that the distribution of GAPDH staining was coincident with nuclear contours in anti-CD3-stimulated CD4+ TEMs (Fig. 3B). In contrast, CD28SA-stimulated CD4+ TEMs had more dispersed cytosolic distribution of GAPDH (Fig. 3B).
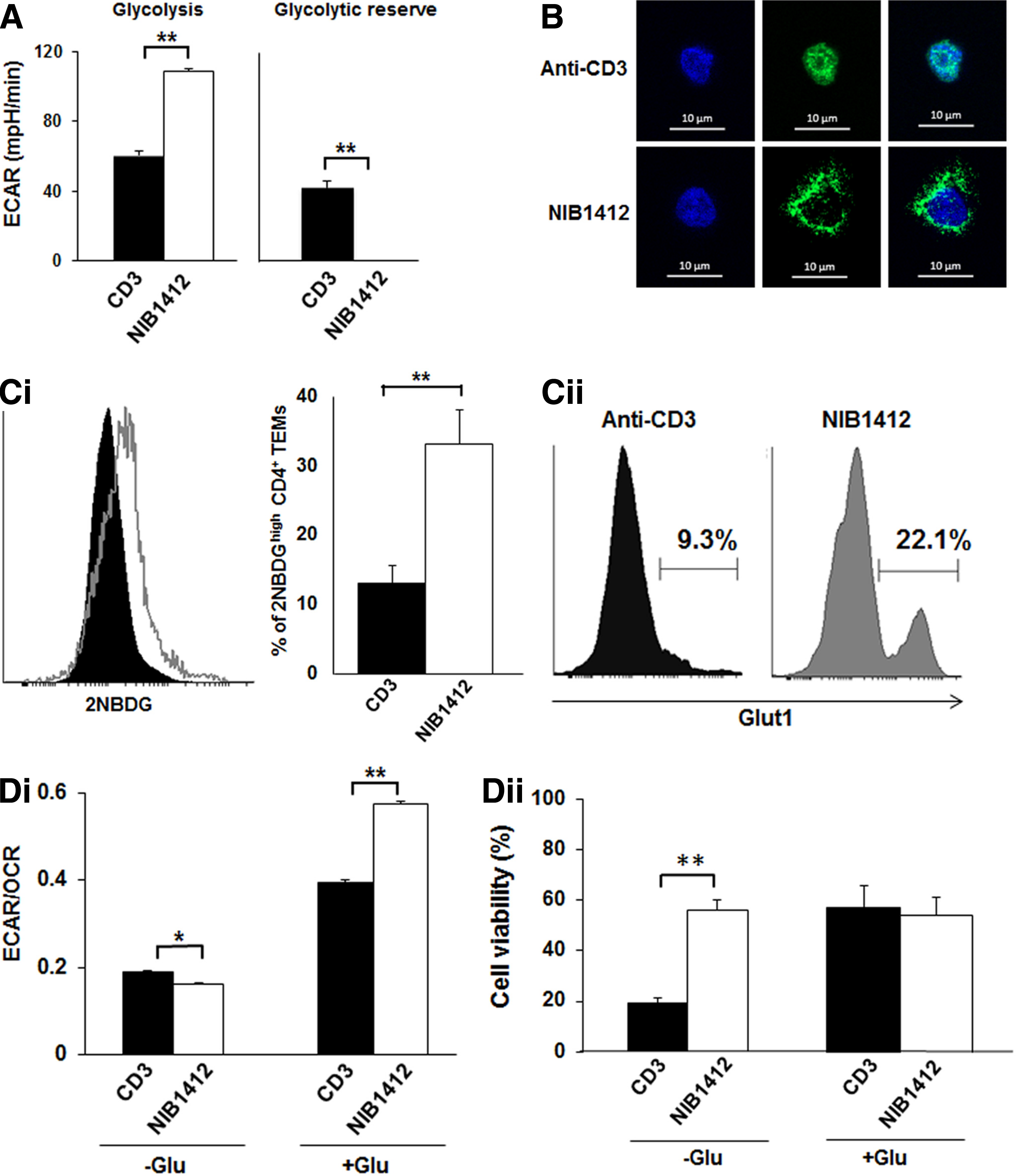
CD28SA stimulation maximizes glucose utilization.
Increased glycolytic rates need to be fuelled by a commensurate increase in the uptake of glucose, which is predominantly mediated by the glucose transporter, GLUT1 in T cells. The uptake of the fluorescent glucose analog 2-NBDG (Fig. 3C-i) and cell surface expression of GLUT1 (Fig. 3C-ii) were both higher in CD28SA-stimulated CD4+ TEMs than in CD3-stimulated cells. The ratio of the basal ECAR to OCR indicates the cellular preference for glycolysis versus OXPHOS metabolic programs.(12) Figure 3D-i shows the ECAR/OCR ratio of CD28SA- and anti-CD3-stimulated T cells cultured (in base media with glutamine only) in the absence of glucose (−Glu) or in the presence of glucose (+Glu). In the −Glu condition the ECAR/OCR ratio was significantly lower in CD28SA-stimulated cells than anti-CD3-stimulated cells. In the +Glu condition CD28SA-stimulated cells had a significantly higher ECAR/OCR ratio than anti-CD3-stimulated cells. These plots demonstrate that in the presence of glucose, CD28SA-stimulated cells preferentially drive glycolysis, whereas anti-CD3-stimulated cells also drive OXPHOS. Collectively these results show that CD28SA stimulation drives an adaptable metabolic program that maximizes glucose utilization to drive both OXPHOS and glycolysis depending on glucose availability. This is further demonstrated by cell viability in glucose-deficient conditions wherein viability of CD28SA-stimulated cells was unaffected after 24 hours while more than 50% of the CD3-stimulated cells died (Fig. 3D-ii).
CD28SA-activated T cells are metabolically programmed to favor lipogenesis
To determine whether mitochondrial fatty acid oxidation (FAO) contributes to OXPHOS, we measured changes in SRC in the presence of the Carnitine palmitoyl transferase (CPT1a) inhibitor, etomoxir (CPT1a is involved in transfer of fatty acids from cytosol to the mitochondria). In both +Glu and −Glu conditions, around 80% of FAO contributed to the SRC of CD3-stimulated CD4+ TEMs. In contrast, only about 30% of FAO contributed to the SRC of CD28SA-stimulated cells in −Glu while there was no contribution of FAO to the SRC in the +Glu conditions (Fig. 4A). The low rate of FAO in CD28SA-activated CD4+ TEMs suggested the possibility that these cells are potentially increasing their lipid reserve. LipidTox staining revealed that CD28SA stimulation induces a significantly greater accumulation of neutral lipids in CD4+ TEMs than when activated with anti-CD3 mAbs (Fig. 4B-i). This was confirmed and quantified by flow cytometry (Fig. 4B-ii). We next measured the acetyl-CoA content and demonstrate that total acetyl-CoA levels were significantly higher in CD28SA-stimulated CD4+ TEMs than in CD3-stimulated cells (Fig. 4C-i). The increased lipogenesis in CD28SA-stimulated cells is reflected by the hyperactivity of the lipogenic enzymes, ACL and ACC, as determined by their phosphorylation status (Fig. 4C-ii). Hyperactivation of ACL and ACC has been reported in tumor cells and we confirmed this in several types of tumor cell lines (Fig. 4C-iv).
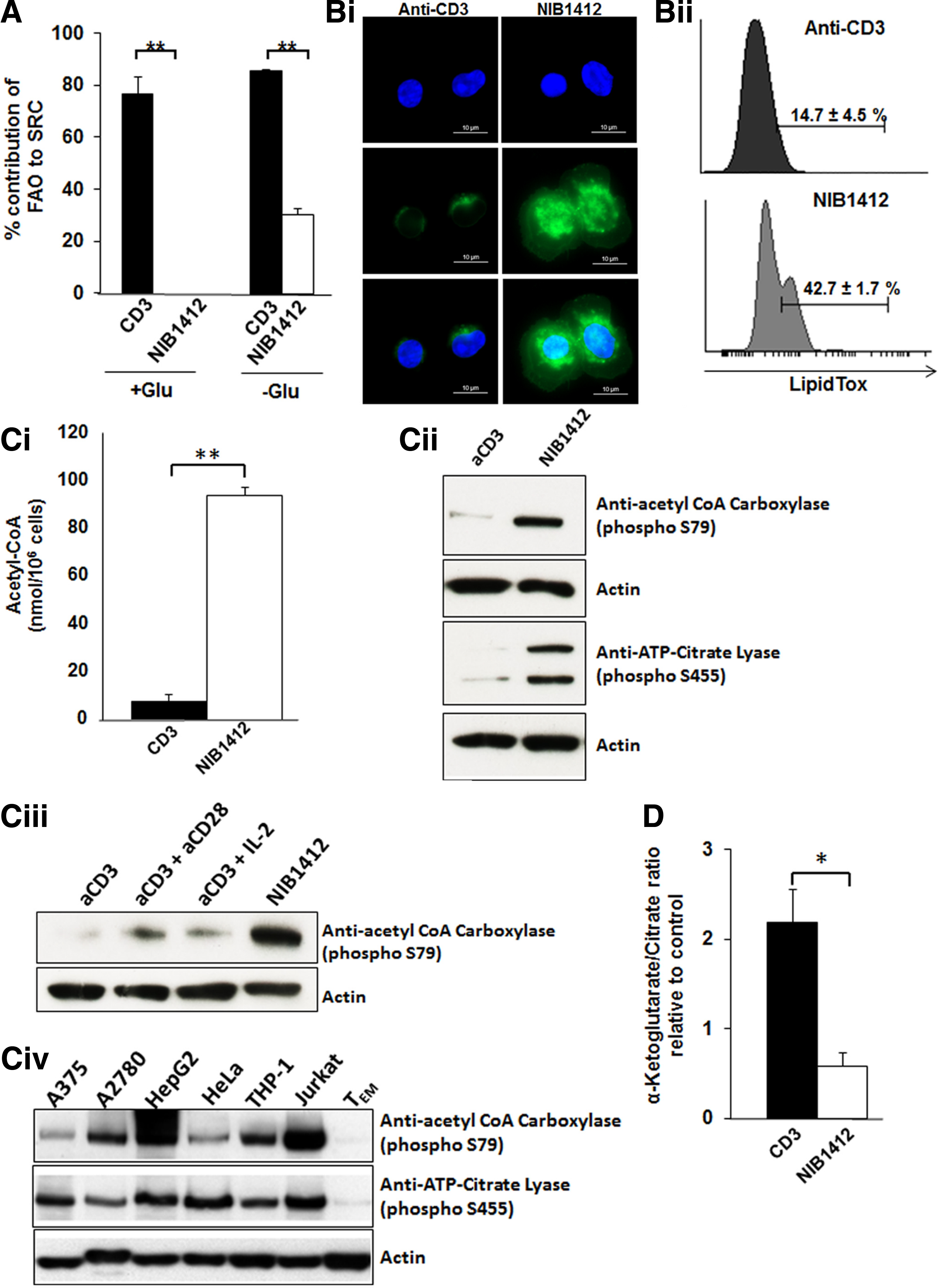
CD28SA activation induces de novo lipogenesis.
To rule out the influence of the CD28SA-secreted IL-2 on the de novo lipogenesis, we also activated CD4+ TEMs with anti-CD3 in combination with IL-2 (aCD3+IL-2), and with anti-CD3 in combination with conventional anti-CD28 (aCD3+aCD28). As can be seen in the results, CD28SA-stimulated cells had significantly greater ACC phosphorylation than any of the other conditions (Fig. 4C-iii). This indicates that the enhanced lipogenic program in CD28SA-activated CD4+ TEMs is not secondary to IL-2 effects. We observed that CD3-stimulated CD4+ TEMs possessed a high relative α-KG/citrate ratio of 2.2 ± 0.3, suggesting ongoing reductive glutamine metabolism (Fig. 4D). In contrast, CD28SA-stimulated cells had a low relative α-KG/citrate ratio of 0.6 ± 0.1 indicative of an absence of reductive glutamine carboxylation (Fig. 4D).
Collectively, these results show that CD28SA-stimulated cells have a dominant program of endogenous biosynthesis of fatty acids, which was not accompanied by preferential reductive glutamine metabolism. Our results show that CD28SA stimulation induces a tumor cell-like metabolic phenotype, where increased glycolytic flux correlates with elevated de novo lipogenesis and reduced FAO.
Discussion
The metabolic programming of T cells is an area of intense investigation with implications for therapeutic intervention in many disease areas.(13–16) We and others have previously shown that CD28SA activation results in a hyperactive T cell phenotype.(4,17) Activated immune cells prefer glycolysis over OXPHOS as it is around two orders of magnitude faster for biomass accumulation and proliferation.(18) Nevertheless, OXPHOS is necessary for cell-surface expression of the IL-2 receptor and thus vital for lymphocyte proliferation.(19)
In the current study, we find that CD28SA-stimulated TEMs cells had higher levels of ATP production, basal respiration, and actively respiring mitochondria compared with anti-CD3-activated TEMs. The high oxygen consumption and ATP production is indicative of enhanced rates of OXPHOS, which occurs primarily in the mitochondria.(20) Studies have shown that an increase in mitochondrial mass results in an increase in overall mitochondrial function, with a boost in residual OXPHOS capacity and an increase in overall ATP-generating capacity.(21,22) Furthermore, our observation of increased mitoROS in CD28SA-stimulated TEMs cells is reflective of enhanced mitochondrial electron transport activity.(23) MitoROS production is regulated by mitochondrial membrane potential (Δψm), metabolic state of mitochondria, and oxygen concentration.(24) Mitochondrial mass and Δψm are known to be significantly increased in cells during cell cycle, particularly when they are in S phase(25) and we have shown in a previous study(4) that a high proportion of CD28SA-stimulated cells are in S-phase at 48 hours postactivation and onward. Thus, the elevated mitochondrial mass and mitoROS might be due to the rapid cell proliferation induced by CD28SA activation. We also observed that CD28SA-stimulated cells proliferated in both normoxic and hypoxic conditions with higher rate of proliferation in the latter. These observations indicate that CD28SA-activated T cells have a flexible metabolic program that reduces the reliance on OXPHOS for proliferation.
We observed that CD28SA stimulation maximizes glucose utilization with higher glycolysis and a deficient glycolytic reserve. The T cell machinery has to be configured to support the higher glycolytic rate. Studies have shown that GAPDH shuttles between nuclear and cytoplasmic compartments, with cytoplasmic GAPDH being active in glycolysis,(26) while nuclear GAPDH is thought to be involved in the initiation of apoptotic cascades(27) and promoting gene transcription.(28,29) We observed a predominant cytosolic distribution of GAPDH in CD28SA-stimulated TEMs as well as increased cell surface GLUT1 expression, supportive of enhanced glucose uptake and utilization.(30) In keeping with a role for CD28 in this process, CD28 costimulation (in addition to TCR activation), acting through the PI3K/Akt pathway is an important component in increasing glycolytic flux essential for T cell proliferation.(31) Interestingly, the adaptable metabolic program induced by CD28SA in T cells (which is accompanied by supportive cellular changes) is reminiscent of the metabolic and cellular changes seen in tumor cells. For example, a variety of tumor cells show upregulation of GAPDH together with its accumulation in the cytoplasm,(29) exhibit elevated levels of glucose uptake and glucose transporters, and(32) can survive in vitro in the complete absence of glucose if supplied with glutamine.(33)
Mitochondrial FAO is a source of acetyl CoA that is used in the TCA cycle for energy generation through OXPHOS.(34) CPT1a is a metabolic enzyme that controls the rate-limiting step in mitochondrial FAO.(35) IL-2-secreting CD8+ effector memory T cells store exogenous long-chain fatty acids in lipid droplets, which are then mobilized for mitochondrial FAO.(8) In contrast, CD8+ memory T cells were shown to use extracellular glucose to synthesize fatty acids for neutral lipid stores,(8) which then undergo lipolysis to supply fatty acids for FAO as well as for incorporation into cellular membrane structures.
The lipid reserve of T cells is determined by the balance between lipogenesis and rate of FAO.(8) De novo fatty acid biosynthesis is not only required to synthesize new membranes for cell proliferation but also to facilitate the formation of lipid rafts for increased signaling of cell surface receptors.(36) Accumulated neutral lipid stores are then mobilized to provide free fatty acids to promote production of signaling lipids, for incorporation into cellular membranes or for the production of ATP through β-oxidation.(37) Although the dynamics of fatty acid utilization for FAO has not been shown in CD4+ TEMs, in this report we propose that CD28SA stimulation enables CD4+ TEMs to promote endogenous biosynthesis of fatty acids similar to cancer cells, which are known to undergo a metabolic shift toward pronounced increase in de novo lipid biosynthesis.(38)
Lipogenesis requires the availability of acetyl-CoA, which is an essential substrate for the endogenous biosynthesis of fatty acids. Increased glycolysis can shunt glucose-derived pyruvate into the mitochondria to be decarboxylated to acetyl-CoA, and then condensed with oxaloacetate to form citrate.(39) Citrate is then exported from the mitochondria through the malate citrate shuttle system and used as a substrate for ACL.(40) ACL is a key cytosolic enzyme that catalyses the generation of acetyl-coenzyme A (CoA) from mitochondria-derived citrate. Acetyl-CoA is then carboxylated to malonyl-CoA by ACC, which subsequently results in fatty acid synthesis. The activity of ACL and ACC are increased by serine phosphorylation.(41,42) We report that total acetyl-CoA levels were significantly higher along with increased activity of ACL and ACC in CD28SA-stimulated TEMs than in CD3-stimulated cells. We also show that IL-2 derived from CD28SA-stimulated T cells(43) did not contribute to the enhanced lipogenic program observed in CD28SA-activated TEMs. The rapid proliferation of several types of cancer cells is dependent on the enhanced activity of ACL, with elevated acetyl-coA levels and increased activity of lipid-metabolizing enzymes being increasingly recognized as a feature of cancer cells.(44,45) This metabolic program is now observed in CD28SA-stimulated TEMs.
ACL and ACC represent committed steps in channeling glucose-derived metabolites toward a lipid biosynthetic fate, which is regulated by PI3K/Akt signaling.(46) This is consistent with the activation of the PI3K pathway by engagement of CD28 by the superagonistic mAb.(47) The acetyl CoA that feeds lipogenesis is primarily derived from citrate.(48) The citrate can be produced from glucose-derived pyruvate that enters the TCA cycle. A recent study has shown that in cancer cells, glutamine can enter the TCA cycle and also become a source of citrate for de novo lipogenesis through reductive carboxylation of glutamine-derived α-KG(49) (Supplementary Fig. S1). An elevated α-KG/citrate ratio was shown to be the principal driving force for reductive glutamine metabolism(49) and we observed a low relative α-KG/citrate ratio in CD28SA-stimulated TEMs cells indicative of an absence of reductive carboxylation, unlike anti-CD3-stimulated TEMs. Elevated glycolytic flux is associated with de novo lipogenesis and reduced FAO. CD28SA-stimulated TEMs exhibited increased glycolytic flux similar to cancer cells where intermediate metabolites are channeled toward acetyl-CoA generation for use in de novo fatty acid biosynthesis.(36)
The tumor microenvironment (TME) is rendered hostile to the survival and function of tumor infiltrating effector T cells by the remodeling effects of tumor cells. Tumor cells achieve this through a number of mechanisms, including establishing a hypoxic milieu, depleting extracellular glucose, secreting metabolites that impair immunity and by enhancing expression of ligands for immune checkpoints such as PD-1 to maintain T cell exhaustion.(50) Current immunotherapies based on checkpoint inhibitors target limited pathway/s involved in immune evasion. A comprehensive immunotherapeutic strategy should be aimed at increasing T cell fitness through countering multiple aspects of tumor cell remodeling of TME. Our results suggest that T cells that have been metabolically reprogrammed through the CD28SA pathway will have superior fitness in the TME owing to their increased tolerance to hypoxia and a flexible metabolic program, much alike tumor cells. Furthermore, we have also previously shown that CD28SA-activated T cells can escape inhibition through the PD-1 pathway.(4) Our findings indicate that immunotherapies that induce T cell metabolic reprogramming similar to CD28SA-activated T cells will enhance the efficacy of tumor-infiltrating T cells. For example, it can be envisaged that chimeric antigen receptor T cells that are modified to exploit the metabolic reprogramming of the CD28SA pathway will have superior efficacy and persistence in the TME.
In summary, we have shown that CD28SA activation induces a tumor cell-like T cell metabolic program that is geared toward maximal glucose utilization, endogenous lipid synthesis, and less dependence on OXPHOS. Tumor cells frequently exhibit similar alterations in metabolic activity to support the increased production of metabolic intermediates for the synthesis of proteins, nucleic acids, and lipids. We have also defined ACL and ACC as the likely molecular components that support operation of this metabolic program. Our study provides insights into the remarkable capacity of T cells to adopt a variety of distinct metabolic programming depending on the pathways that are engaged.
Our results may also have implications and utility in drug development, as dysregulated metabolic patterns in immune cells treated with therapeutic mAbs could serve as novel biomarkers of hazard identification. For example, aberrant nutrient uptake and utilization, together with increased de novo lipid biosynthesis could be indicative of excessive immunostimulatory potential of the therapeutic mAb. Metabolic studies that compare a number of immunostimulatory therapeutic mAbs are required to examine this further.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.