
Abstract
This study evaluated the effects of long-term intake of nutritive sweeteners (NSs) and non-nutritive sweeteners (NNSs) on body weight, food and energy intake, blood pressure, metabolic parameters, and memory retention in rats. Sixty male Sprague–Dawley rats were randomly divided into six groups (n = 10 per group): control (water),10% sucrose (SUC), aspartame (ASP), sucralose (SCA), stevia (STV), and 5% xylitol (XYL). Pure NSs (SUC and XYL) and NNSs were added to the drinking water for 18 weeks. ASP, SCA, and STV dosage was based on the estimated daily intake limit: 4.1, 2.0, and 3.4 mg/kg/day, respectively. Chronic access to NNSs did not result in any difference in total weight gain of the rats, while it was significantly elevated in the SUC group compared with the control and NNSs groups. Food intake was significantly lower in all NNSs groups compared with SUC and control groups. Sweetened beverage intake volumes were significantly diminished in all NNSs groups compared with intake in SUC and control groups. Total calories consumed were lower for the STV and XYL groups compared with all other groups. Blood pressure and glucose metabolism did not differ significantly between the groups. All sweeteners increased total cholesterol, low-density lipoprotein, and triglyceride levels. Short-term memory was significantly impaired in the ASP group in the novel object recognition task, while long-term memory was impaired in SUC and STV groups. These metabolic and behavioral results suggest that the long-term intake of NSs or NNSs can be associated with peripheral and central effects.
INTRODUCTION
The high consumption of added sugars has become a health problem worldwide. Excessive intake of sugary foods and beverages can cause metabolic and hormonal changes and promote the accumulation of body fat. 1 Non-nutritive sweeteners (NNSs) can be a helpful strategy to reduce energy intake and body weight, thus reducing the risk of type 2 diabetes and metabolic syndrome. 2 Considering the prevalence of these metabolic diseases, NNSs are becoming progressively more popular as an alternative to sugar.
Consumption of NNSs, also called low-calorie sweeteners/intense sweeteners, makes it possible to reduce the energy density of foods and beverages while maintaining high palatability. 2 The NNSs currently approved by the authority of the U.S. Food and Drug Administration (FDA) as safe for human consumption (GRAS category) are saccharin, aspartame (ASP), sucralose (SCA), neotame, acesulfame potassium, stevia (STV), advantame, and Luo Han Guo fruit extracts. 3
The FDA has established an estimated daily intake (EDI, expressed in mg/kg body weight) limit for each NNSs; this value is a calculated level of exposure that the consumer is unlikely to exceed on a chronic basis and it is below the acceptable daily intake level. 4 Before the FDA-granted GRAS status to NNSs, clinical and toxicological studies were conducted in various species, including humans, to demonstrate that NNSs are safe and well tolerated.
However, accumulative evidence suggests a positive association between NNSs consumption and weight gain, metabolic syndrome, and type 2 diabetes. 5 –7 Therefore, it has been suggested that NNSs are not innocuous compounds because they can affect several neurohormonal and biochemical processes involved in energy regulation and glucose homeostasis.
NNSs have different chemical structures, but they all have in common the ability to activate some of the multiple ligand binding sites of the heterodimeric sweet taste receptor, T1R1+T1R3. 5 Therefore, one of the proposed mechanisms underlying the metabolic effects of NNSs is their interaction with the T1R family of sweet taste receptors through α-gustducin (receptor-associated subunit of the G-protein complex) 8 ; this associated G-protein-coupled receptor signaling can explain in what way NNSs contribute to developing metabolic syndrome and insulin resistance. Likewise, NNSs have been associated with impaired incretin and insulin secretion, which may eventually affect learned responses to sweetness. 9 Furthermore, NNSs also interfere with the composition and function of the gut microbiota. 10,11
The possibility of the development of metabolic syndrome, and the mechanisms underlying this effect, could indirectly relate NNSs to deleterious effects on the brain and cognitive processes; however, this association has been less studied. In this sense, ASP and potassium acesulfame have been the NNSs most widely studied for their possible effects on neurobehavioral health. For example, several studies link ASP with alteration in monoamine neurotransmitter synthesis, which might generate neurophysiological symptoms, including cognitive deficit, headache, seizure, migraines, mood disorders, and insomnia. 12,13 Likewise, potassium acesulfame has been related to alterations in neurometabolic functions that could affect cognitive functions. 14,15
Despite all the above, no studies have compared the nutritive sweeteners (NSs) and NNSs effects on memory and metabolic variables. Therefore, the present study investigated the effects of chronic oral administration of ASP, SCA, STV (steviol glycosides), xylitol (XYL), or sucrose (SUC) on food and caloric intake, water consumption, body weight, blood pressure, metabolic parameters, and memory retention in rats.
MATERIALS AND METHODS
Sixty male Sprague–Dawley rats (200–250 g) were purchased from the Institute of Neurobiology, National Autonomous University of Mexico, Campus Juriquilla. Before beginning the treatments, the animals were housed in groups (n = 5) in polycarbonate cages for 2 weeks under an inverted 12-h light–12-h dark cycle (lights on at 9 p.m.) in a temperature-controlled (22°C) room. All animals had free access to food (LABDIET 5012; 3.43 kcal/g [27% protein, 13% fat, and 60% carbohydrates]) and water throughout the experiments.
All procedures were conducted in accordance with the Guidelines for the Use and Care of Laboratory Animals (NOM-062-ZOO-1999, Ministry of Agriculture and Livestock, SAGARPA, Mexico), and the protocol was approved by the Institutional Committee for the Care and Use of Laboratory Animals. We took special care to minimize animal suffering and the number of animals used. All experiments were performed between 10:00 and 15:00 h, and independent groups were used.
Treatment groups
Rats were randomly divided into six groups (n = 10 per group): control (water), SUC, ASP, SCA, STV, and XYL. SUC, XYL, and pure NNSs were kindly donated by a local company (CYTECSA S.A. de C.V.) and were added to the drinking water for 18 weeks. Ten percent SUC and 5% XYL solutions were used; ASP, SCA, and STV dosage was based on the FDA-approved upper level of the EDI limit: 4.1, 2.0, and 3.4 mg/kg/day, respectively. 4
Measuring food and water intake and body weight gain
For each of the 18 weeks of treatment, food and caloric intake and water consumption were measured every 24 h (10:00–12:00 h). Weight gain was calculated by subtracting the basal weight from the weight obtained every week and expressed in grams. Sweetened water intake was determined by monitoring the water/sweetener solution volumes. The dose for each animal based on the average daily fluid consumption per day and body weight was calculated using an Excel fact sheet.
For the majority of the experimental period, the required doses were reached, except for ASP in the last four weeks, where a modest increase was observed (doses per week on average: ASP, 7.09 ± 1.95; SCA, 2.96 ± 0.86; and STV, 4.67 ± 1.29). Total caloric intake (including sweetener and chow) was calculated by the sum of calories ingested each week, corrected by the corresponding rat weight at the end of each week, and expressed as kcal/g.
All rats were euthanized using decapitation at the end of the experimental period and after overnight fasting. 16 After decapitation, 1 mL of trunk blood was collected in Barricor™ lithium heparinized tubes and centrifuged at 2000 g for 10 min at 4°C; plasma was removed, and aliquots were used to measure blood lipid levels.
Blood pressure measurement
Arterial blood pressure (systolic, diastolic, and mean pressure) was assessed using a computerized tail-cuff method (CODA 4; Kent Scientific Corporation, CT, USA) at the end of the study. 17 The rats were trained for 5 consecutive days by placing them on a plastic rack mounted on a thermostatically controlled hot plate maintained at 35°C. The habituation period was followed by a test session consisting of 20 blood pressure and heart rate measurements. All blood pressure measurements were made between 10:00 and 14:00 h.
Intraperitoneal glucose tolerance test
The intraperitoneal glucose tolerance test (IPGTT) was performed on overnight-fasted rats to evaluate glucose tolerance at the end of the study. Rats were fasted for 8 h and then given a glucose load (1.5 mg/kg body weight, intraperitoneal), and blood samples were collected from the tail vein at 0, 15, 30, 60, 90, and 120 min after the glucose was given. The glucose levels were determined using a CardioChek™ analyzer (Polymer Technology Systems, Inc., IN, USA).
Determination of blood lipid parameters
Blood lipid parameters were measured at the end of the study. Total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), and triglyceride (TG) levels were determined using the LABGEO PT10 clinical chemistry analyzer (Samsung Electronics Co. Ltd., South Korea).
Novel object recognition task
The novel object recognition task (NORT) is used to study short- and long-term memory and is based on the natural tendency of rodents to explore new objects and compare them with familiar ones. 18 The rats were habituated to the arena (60 × 60 × 45 cm), without any objects, for 5 min at 24 h before testing. The test involves two 5-min trials separated by 1 h (short-term memory) or 24 h (long-term memory). 19
During the familiarization trial (FT), two identical objects (A1 and A2) were placed in opposite corners, ∼5 cm from the walls of the open field. In the retention trial, one of the objects was replaced with a novel object (A = familiar and B = novel). The animals were returned to the home cage after the FT. Exploring an object is defined as directing the nose toward the object at a distance less than or equal to 2 cm. The behavior of rats was recorded using a camera placed above the arena and connected to the ANY-maze® tracking system (Stoelting Co., IL, USA).
In addition, the distance traveled was automatically measured using the ANY-maze tracking system. Based on the exploration time (ET) of the two objects, a recognition index (RI) was calculated as RI = (ETB − ETA)/(ETA + ETB). An RI value above 0.5 indicates a novel object preference and can be interpreted as normal cognitive performance.
Statistical analyses
Data are expressed as mean ± SEM and were analyzed with a one-way ANOVA or Kruskal–Wallis analysis (depending on whether populations were normally distributed or not), followed by a post hoc Tukey multiple comparison test. Other statistical analyses were performed using unpaired t-test or linear regression. A P value <.05 was considered statistically significant. The area under the curve (AUC) was calculated using the trapezoidal method.
RESULTS
Physiological effects of chronic NNSs ingestion
Data for physiological parameters of different experimental groups at the end of the 18-week experimental period are shown in Table 1. Body weight gain and food, caloric, and water intake of the rats were monitored during the experiment. Chronic access to NNSs did not result in any difference in total weight gain of the rats, while it was significantly elevated in the SUC group compared with the control and NNSs groups (F[5, 49] = 4.09, P < .01; P < .05 for post hoc comparison).
Data for Physiological Parameters of Different Experimental Groups at the End of the 18-Week Experimental Period
Values are mean ± SEM for each group of rats.
P < .05 versus the control group.
P < .05 versus all others groups.
ASP, aspartame; CON, control; SCA, sucralose; STV, stevia; SUC, sucrose; XYL, xylitol.
Food intake was significantly lower in all NNSs groups compared with SUC and control groups (overall effect, F[5, 49] = 26.60, P < .01; P < .01 for each post hoc comparison). Unlike for the ASP group, sweetened beverage intake volumes were significantly diminished in NNSs groups compared with intake in SUC and control groups (overall effect, F[5, 49] = 14.80, P < .01; P < .05 for each post hoc comparison). Total calories consumed were lower for the STV and XYL groups than all other groups (overall effect, F[5, 49] = 14.20; P < .01; P < .01 for each post hoc comparison).
Our data indicate that the parameters of arterial pressure do not differ significantly between the groups (systolic: F[5, 49] = 1.56, P = .18; diastolic: F[5, 49] = 1.79, P = .12; and mean: F[5, 49] = 1.67, P = .15). However, when comparing only the control group (135.74 ± 4.50) and STV group (149.16 ± 4.23), we found a significant increase in systolic pressure (t[0.05, 8] = −2.17, P < .04).
Effects of chronic NNSs ingestion on biochemical parameters
The effects of SUC and NNSs on biochemical markers after their administration for 18 weeks are shown in Table 2. Plasma glucose concentrations did not differ significantly between the groups (F[5, 29] = 1.91, P = .13). The AUC of plasma glucose concentrations during the IPGTT showed no significant differences (F[5, 29] = 1.89, P = .13), although in the sucrose group, the AUC was markedly higher than that of the control group (t[0.05, 8] = −2.735, P < .05).
Data for Serum Biochemical Parameters of Different Experimental Groups at the End of the 18-Week Experimental Period
Values are mean ± SEM for each group of rats.
P < .05 versus the control group.
AUC, area under the curve; HDL, high-density lipoprotein; LDL, low-density lipoprotein.
Unlike HDL-C, chronic consumption of all sweeteners increased plasma TC (overall effect, F[5, 29] = 6.08, P < .01; P < .01 for each post hoc comparison); LDL-C (overall effect, F[5, 29] = 4.91, P < .01; P < .05 for each post hoc comparison), and TG (overall effect, F[5, 29] = 4.05, P < .05; P < .05 for each post hoc comparison) levels.
Chronic NNSs ingestion in rats impairs cognitive function
Episodic memory was assessed using the RI in NORT (Fig. 1). In the FT, there were no significant differences in total time spent exploring the objects (H = 3.209, P = .668). Short-term memory was significantly impaired in the ASP group (H = 11.121, P < .05), while long-term memory was impaired in SUC and STV groups (H = 13.268, P < .05). Additionally, no significant differences were found in other measures of exploration (distance traveled), indicating that memory impairment cannot be attributed to alterations in motor activity (F[5, 42] = 0.328, P = .891).
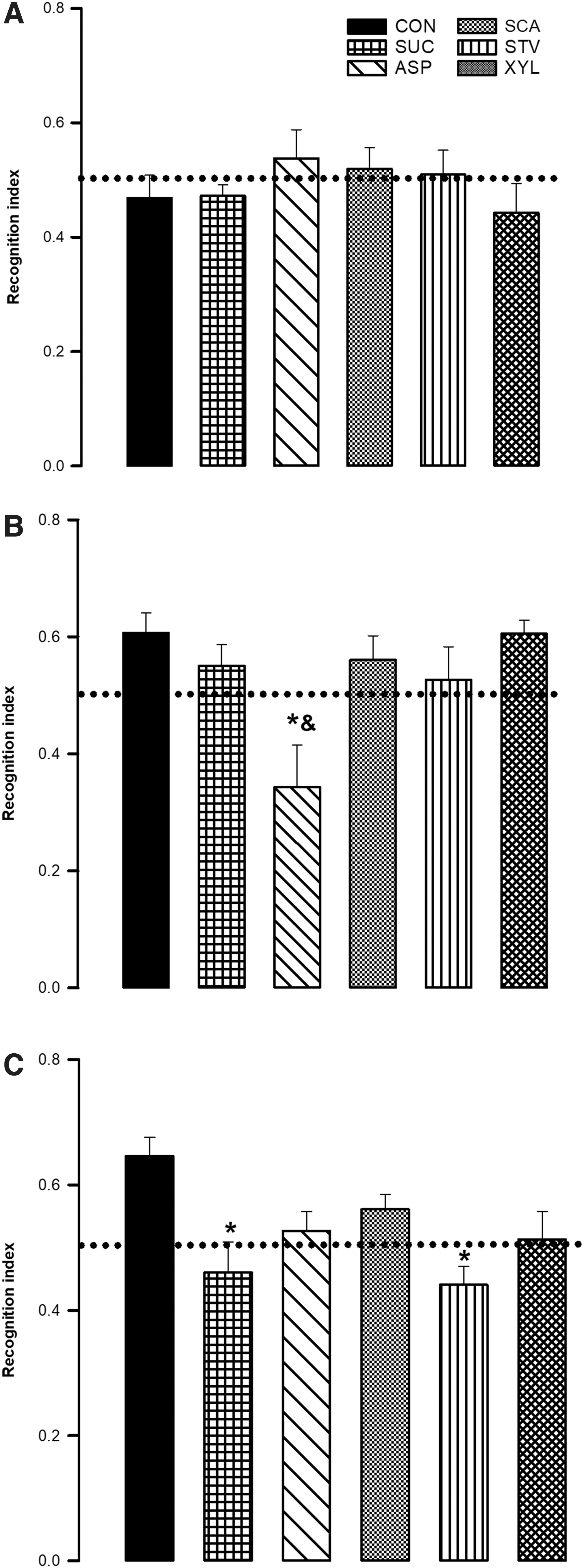
Effects of long-term intake of nutritive and non-nutritive sweeteners on recognition memory in adult male rats. Values are expressed as the mean ± SEM for each group of rats.
Considering the metabolic and behavioral results, to find out if there was an association between the plasma values of the lipid profile and the cognitive deficit found with ASP, STV, and SUC, a linear regression was performed between the RI and values of plasma lipids. No significant association was found in any of the cases.
DISCUSSION
This study investigated both peripheral and central effects of the long-term intake of NSs and NNSs. The main findings from this work include (1) long-term intake of NSs and NNSs increased plasma TC, LDL-C, and TG levels; (2) chronic consumption of ASP impaired short-term memory; and (3) chronic intake of STV and SUC impaired long-term memory.
The negative impact of added sugar consumption on weight and other health outcomes is well recognized; therefore, many people have changed to high-intensity sweeteners such as ASP, SCA, and STV to reduce the risk of potential consequences. However, accumulating data suggest that frequent consumption of these sugar substitutes may also lead to an increased risk of cardiometabolic diseases. 7,11
It has been suggested that the use of NNSs can lead to weight gain, but evidence regarding their true effect on body weight and satiety is still lacking. The present study compares the effects of NNSs with NSs on body weight gain and food, caloric, and water intake using a rat model. Despite the slight decrease in food intake in this study, no changes were observed in total weight gain after long-term intake of NNSs and XYL. These results are consistent with several articles that reported no effects of ASP, 20 –22 SCA, 21,23 STV, 21,24 –26 and XYL 27,28 on weight gain.
Contrarily, the SUC group showed that a reduction in food intake was accompanied by an increase in fluid intake and body weight gain; these findings of SUC have been well documented and confirm the hypothesis that animals adjust for calories consumed on one occasion by reducing their caloric intake on subsequent opportunities to eat. 29
Regarding blood pressure, no effect was found with NNSs or NSs consumption; when reviewing studies available in the literature, no effect has been reported with NNSs administration in normal rats. Specifically, in the case of STV, it has been reported that it reduces blood pressure levels in rat models 30 and hypertensive humans. 31 The finding of the possible hypertensive effect was not expected, and more studies are needed to investigate if it is reproducible and the possible underlying mechanisms.
In this study, an evaluation of the glycemic response was performed through IPGTT. Neither at minute 0 after glucose injection (fasted) nor in the AUC of IPGTT was any significant difference observed between the NNSs and NSs groups. These findings agree with those reported in the literature for healthy subjects and rats. 27,28,30,32,33 In addition, SUC elicited a higher postprandial glycemic profile that agrees with the tolerance reported in rodents. 34
Regarding the lipid profile, it was found that all the sweeteners used increased plasma levels of TC, LDL-C, and TG. In the case of SUC, 33,34 ASP, 35 and SCA, 33 the effects correspond to the data already published. ASP is related to liver damage and may influence various metabolic pathways, thereby enhancing the levels of plasma lipids.
Different studies have reported that STV has no adverse effects on the lipid profile; some have even shown its beneficial effect. 36 However, our findings agree with Farid et al., who reported that long-term consumption of STV elevated levels of liver function enzymes (ALT and AST), TC, LDL-C, and proinflammatory cytokines (IL-6 and -8) and led to oxidative stress. 37 On the other hand, intestinal bacteria hydrolyze steviol glycosides to steviol. It is absorbed through the portal vein and reaches the liver, where it is metabolized to steviol glucuronide, and is excreted in the urine.
Thus, our data and those of Farid et al. could be related to stress on the liver and the possibility of intestinal flora reshaping. 38 About XYL, its metabolite, xylulose 5-phosphate, through activation of the carbohydrate response element-binding protein (ChREBP), stimulates the genes associated with a lipogenic enzyme such as acetyl-coenzyme A carboxylase and fatty acid synthase. Thus, this increased lipogenesis in the liver could alter plasma lipid levels. 39 Despite the findings described above, the long-term effects of NNSs or NSs on lipid metabolism cannot be completely understood, and more studies are needed.
The effect of chronic intake of sweeteners on memory retention was evaluated using the NORT. This study shows that chronic intake of ASP has adverse effects on short-term memory. In this sense, it is well documented that ASP may cause brain oxidative stress, decreased brain glucose availability, and dose-dependent inhibition of brain serotonin, noradrenaline, and dopamine. 39 –41 It caused significant deleterious effects on cognitive responses and memory retention in rats. 12,13,40 –42
A negative effect of SUC on long-term memory has been reported only with a high-SUC diet, and it has been related to alterations in glucose metabolism 43 and maladaptive neuroplasticity. 44 Despite having carried out an exhaustive search on the consumption of STV and its relationship with cognitive deficit, there were no results; instead, it has been linked to preventive effects on high-fructose diet-induced neurocognitive damage. 45
Altogether, our results suggest that cognitive deficit was independent of increased body weight, food and caloric intake, liquid consumption, blood pressure, glucose metabolism, or plasma lipids. On the other hand, our behavioral results support the evidence that suggests that neuronal sweet taste receptors play a key role in synaptic function and memory acquisition. 46 –48
Despite our findings, it is also critical to recognize that NNSs and NSs are compounds with different chemical structures. Due to this difference, it is essential to consider their absorption profiles, metabolism, or excretion pathways to determine their biological effects on human health. However, the possibility that their chronic consumption may increase cardiovascular risk and cognitive alterations reinforces the need for new recommendations on their use.
In summary, our metabolic and behavioral results suggest that the long-term intake of NSs or NNSs can be associated with both peripheral and central effects.
Footnotes
ACKNOWLEDGMENT
The authors thank Ramón Maruri-Gómez for the language revision.
AUTHORs' CONTRIBUTIONS
All authors contributed equally to the writing of the article. In addition, all authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was partially supported by COFAA, SIP-IPN (grants 20171727 and 20200400), and the PAPIIT IA210420 program, DGAPA, UNAM. CONACyT awarded a student grant to E.I.M.R.