
Abstract
Background:
As the COVID-19 pandemic has progressed, numerous variants of SARS-CoV-2 have arisen, with several displaying increased transmissibility.
Methods:
The present study compared dose–response relationships and disease presentation in nonhuman primates infected with aerosols containing an isolate of the Gamma variant of SARS-CoV-2 to the results of our previous study with the earlier WA-1 isolate of SARS-CoV-2.
Results:
Disease in Gamma-infected animals was mild, characterized by dose-dependent fever and oronasal shedding of virus. Differences were observed in shedding in the upper respiratory tract between Gamma- and WA-1-infected animals that have the potential to influence disease transmission. Specifically, the estimated median doses for shedding of viral RNA or infectious virus in nasal swabs were approximately 10-fold lower for the Gamma variant than the WA-1 isolate. Given that the median doses for fever were similar, this suggests that there is a greater difference between the median doses for viral shedding and fever for Gamma than for WA-1 and potentially an increased range of doses for Gamma over which asymptomatic shedding and disease transmission are possible.
Conclusions:
These results complement those of previous studies, which suggested that differences in exposure dose may help to explain the range of clinical disease presentations observed in individuals with COVID-19, highlighting the importance of public health measures designed to limit exposure dose, such as masking and social distancing. The dose–response data provided by this study are important to inform disease transmission and hazard modeling, as well as to inform dose selection in future studies examining the efficacy of therapeutics and vaccines in animal models of inhalational COVID-19.
Introduction
Numerous studies have detected infectious SARS-CoV-2 or viral RNA in air samples collected in the vicinity of infected individuals,1–10 suggesting that respiratory aerosols play a role in transmission of COVID-19. As the pandemic has progressed, numerous variants of SARS-CoV-2 have emerged, with several displaying increased transmissibility.11–15 Several studies have hypothesized that increased viral shedding from infected individuals may be an important factor associated with the increased transmissibility observed with more recent variants of SARS-CoV-2.3,13,16–19 Other potential explanations include enhanced survival of virus during transport through the environment, and/or a lower infectious dose in susceptible individuals, although the relative contributions of these potential mechanisms remain unclear.
A recent study from our laboratory found that the probability of infection following inhalation of an early isolate of SARS-CoV-2, hCoV-19/USA/WA-1/2020 (WA-1), was dose dependent in a nonhuman primate (NHP) model of inhalational COVID-19. Furthermore, the median dose necessary to elicit an immune response was significantly lower than that needed to cause fever, suggesting that exposure dose may be a factor influencing disease presentation. 20 These results are in agreement with several other studies that have similarly demonstrated dose dependence of disease presentation in rodent models of COVID-19.21,22 However, no studies were identified, which compared dose–infectivity relationships for different variants of SARS-CoV-2.
To assess whether changes in the infectious dose or viral shedding contribute to the increased transmissibility associated with more recent variants of SARS-CoV-2, the present study compared dose–response relationships and disease presentation in NHPs infected with an isolate of the Gamma variant of SARS-CoV-2, which has multiple mutations in the S protein, which have been shown to contribute to increased viral fitness,23–25 to the results of our previous study with the earlier WA-1 isolate.
Materials and Methods
Ethics statement
All research was conducted in compliance with the Animal Welfare Act and other federal statutes and regulations relating to animals and experiments involving animals and adheres to principles stated in the Guide for the Care and Use of Laboratory Animals, 26 and was conducted in accordance with a Protocol approved by both the National Biodefense Analysis and Countermeasures Center Institutional Animal Care and Use Committee and the U.S. Department of Homeland Security Compliance and Assurance Program Office. The facility where this research was conducted is fully accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International and maintains a Public Health Service Humane Care and Use of Laboratory Animals Policy assurance. All work was conducted in accredited Biosafety Level 3 (BSL-3)/Animal Biosafety Level 3 (ABSL-3) laboratories.
Virus
An isolate of SARS-CoV-2 Gamma (hCoV-19/Japan/TY7-503/2021; NR-54984, Lot#70043069, BEI Resources; GISAID: EPI_ISL_792683) was provided by National Institute of Allergy and Infectious Diseases (NIAID) and was stored at −80°C until use. The Gamma variant contains numerous mutations in the S protein relative to the WA-1 isolate, some of which have been shown to result in changes in binding affinity and viral replication that potentially contribute to increased viral fitness.23–25 Additional details on this isolate are provided in the Supplementary Data.
Animals
Previous studies have demonstrated that cynomolgus macaques are susceptible to infection with aerosolized SARS-CoV-2 through inhalation, including a previous study from our laboratory that reported dose–response relationships for the earlier WA-1 isolate of SARS-CoV-2.20,27,28 Therefore, in the present study, 21 healthy, adult cynomolgus macaques (Macaca fascicularis; 10 male and 11 female; 4–8 years old; NIH Animal Center, Poolesville, MD), weighing 2.9–6.5 kg at the time of exposure, were utilized to assess dose–response relationships for SARS-CoV-2 Gamma for comparison to our previously reported data with the WA-1 isolate. All animals were serologically naive for SARS-CoV-2 before study as measured using the Euroimmun SARS-CoV-2 S1 ELISA Kit (El 2606-9601 G; Euroimmun Inc.) and negative by reverse transcription–real-time PCR (RT-qPCR) in nasal swabs. Animals were housed in high efficiency particulate air (HEPA)-filtered isolator cages (37324; Carter2 Systems, Inc.) throughout the study to minimize the potential for disease transmission among animals housed in the same room. The mean (±standard deviation) relative humidity and temperature in the housing room over the course of the study were 50.8 ± 2.9% and 23.9 ± 0.7°C, respectively.
Inhalation exposures
Inhalation exposures were conducted identically to those performed as part of our previous study with the WA-1 isolate. 20 The number median aerodynamic diameter of the generated aerosols was 0.78 ± 0.01 μm with a geometric standard deviation (GSD) of 1.44 ± 0.02, similar to the size distribution reported previously for particles exhaled from the human respiratory tract during breathing. 29 The corresponding mass median aerodynamic diameter was 1.51 ± 0.08 μm with a GSD of 1.56 ± 0.02. Additional details on the exposure methodology, as well as a summary of exposure parameters and doses for each animal are included in the Supplementary Data.
Three groups of 6–8 animals were exposed iteratively to aerosolized SARS-CoV-2. Doses for the initial iteration were based on the results of our previous study with the WA-1 isolate. The results from each iteration were utilized to inform dose selection for the subsequent iteration, with the goal of minimizing the confidence interval for the median doses.
Postexposure observations and sample collection
Animals were observed twice daily for clinical signs of illness for 21 days postexposure. Body temperature was monitored continuously utilizing implanted telemetry transmitters (M00; Data Sciences International). An average daily temperature profile (15-minute resolution) for each animal was calculated based on at least 3 days of temperature data from the preexposure period. During the postexposure period, fever was defined as an increase of 1°C above the time-matched average from the preexposure period for ≥2 hours.
Animals were anesthetized (Telazol; 3 mg/kg i.m.) on days 2, 4, 6, 8, 10, 14, and 21 postexposure for collection of blood/serum samples, nasopharyngeal and oropharyngeal swabs, and exhaled breath samples.
Serum samples were utilized to assess seroconversion using a plaque reduction neutralization test (PRNT) and serum chemistries, as described previously 20 and in the Supplementary Data.
Nasopharyngeal and oropharyngeal samples were collected using sterile specimen collection swabs (VF105-80; Vare Health), placed into 15-mL conical tubes containing 3 mL of viral culture medium, and vortexed for ∼5 seconds. For nasopharyngeal swabs, both nares were swabbed using a single swab. Samples were then assayed for infectious virus by microtitration or for the presence of viral RNA using RT-qPCR (see Supplementary Data for details).
Exhaled breath collection utilized an anesthesia mask (Surgivet mask 32393B1; Patterson Veterinary) supplied with HEPA-filtered, dry (relative humidity <10%), compressed air. The mask exhaust airflow was sampled using an Optical Particle Sizer (OPS) (1 L/min; Model 3330, TSI Inc.), and a polytetrafluoroethylene (PTFE) Filter (5 L/min). At the end of the collection period, PTFE filters were resuspended in minimum essential medium for growth (gMEM) and analyzed by microtitration and RT-qPCR. A schematic of the system and additional details on the exhaled breath collection system are provided in the Supplementary Data.
Data analysis
Dose–response relationships for viral shedding, fever, and seroconversion were modeled separately using a univariate GLM approach, with data for each endpoint used to construct a binary variable that reflected whether a subject was positive for the endpoint of interest at any point during the postexposure observation period. Binary data were fit with a probit dose–response model with viral variant treated as a covariate (JMP 16.1; SAS Institute, Inc.). Bivariate analyses similar to those utilized in our previous study were also performed. 20 Other comparisons were performed using Prism (v. 9.2.0; GraphPad Software LLC.). The specific analyses performed for each comparison are detailed in the Results section.
Results
Six of the 21 animals exposed to SARS-CoV-2 Gamma developed transient periods of fever postexposure (Table 1). The median deposited dose for development of fever, as well as the median time to fever onset and median fever duration were similar to those observed in our previous study with WA-1 (Tables 1 and 2). The results of probit analysis suggest that viral variant is not a significant factor influencing the dose–response relationships for fever (p = 0.19 through likelihood ratio test [LRT]). Temperature profiles are shown in Figure 1 and in the Supplementary Data. Gamma-infected animals with fever also displayed a transient lymphopenia that resolved by 4 days postexposure, as well as a borderline increase in interleukin-6 (IL-6) (Supplementary Data).
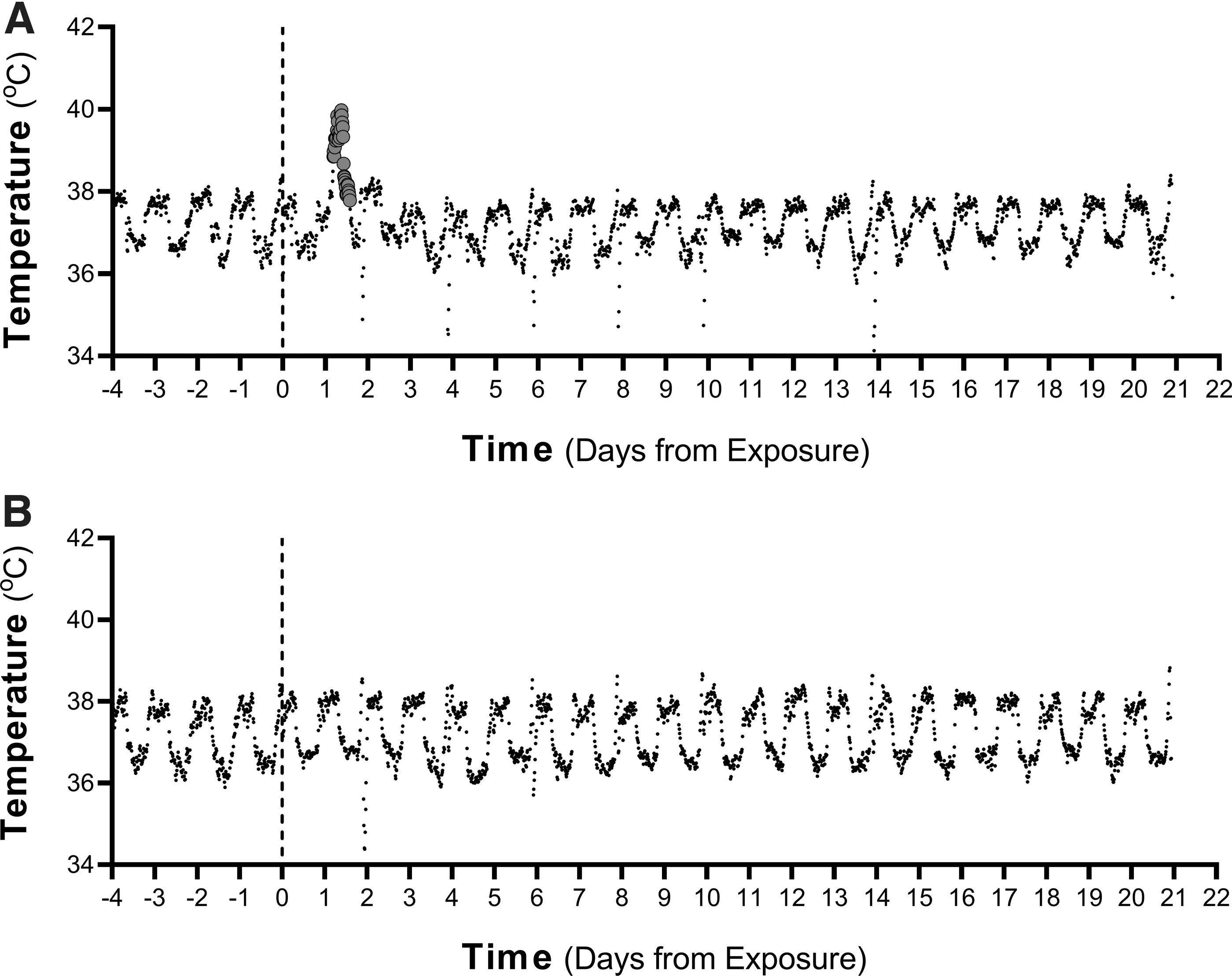
Representative temperature profiles. Temperature profiles are shown for an animal, which developed a transient fever at 29 hours postexposure
Comparison of Fever Metrics for Animals Exposed to SARS-CoV-2 Gamma and WA-1
Median doses for fever were compared through LRT, while the median times for fever onset and fever duration were compared through unpaired t-tests.
CI, confidence interval; LRT, likelihood ratio test; NA, not applicable; NHPs, nonhuman primates; TCID50, median tissue culture infectious doses.
Comparison of Median Doses for Fever, Shedding, and Development of Neutralizing Titers
Median doses for each endpoint are presented with 85% CIs, as comparison of their overlap has been shown to be a reliable indicator of significance for independent dose–response models. Probit fits were also compared through LRT.
PRNT, plaque reduction neutralization test.
Viral RNA and/or infectious virus was detected intermittently in swabs from 14/21 Gamma-exposed animals, including all six that developed fevers (Fig. 2), and in swabs from 10/16 WA-1-exposed animals, including all five that developed fevers (Fig. 3). The results of probit analysis suggest that viral variant is a significant factor influencing the dose–response relationships for shedding of viral RNA (p = 0.03 through LRT) and infectious virus (p = 0.004 through LRT) in nasal swabs. The median dose for shedding of viral RNA or infectious virus in nasal swabs was significantly lower for the Gamma variant than for the WA-1 isolate, when assessed by overlap of 85% confidence intervals, which has been shown to be a reliable indicator of significance for independent dose–response models (Table 2). 30 While shedding occurred at lower doses for Gamma relative to WA-1, neither the peak levels nor total amount of infectious virus or viral RNA detected in swabs during the postexposure observation period were greater for Gamma relative to WA-1 (p > 0.19; Table 3).
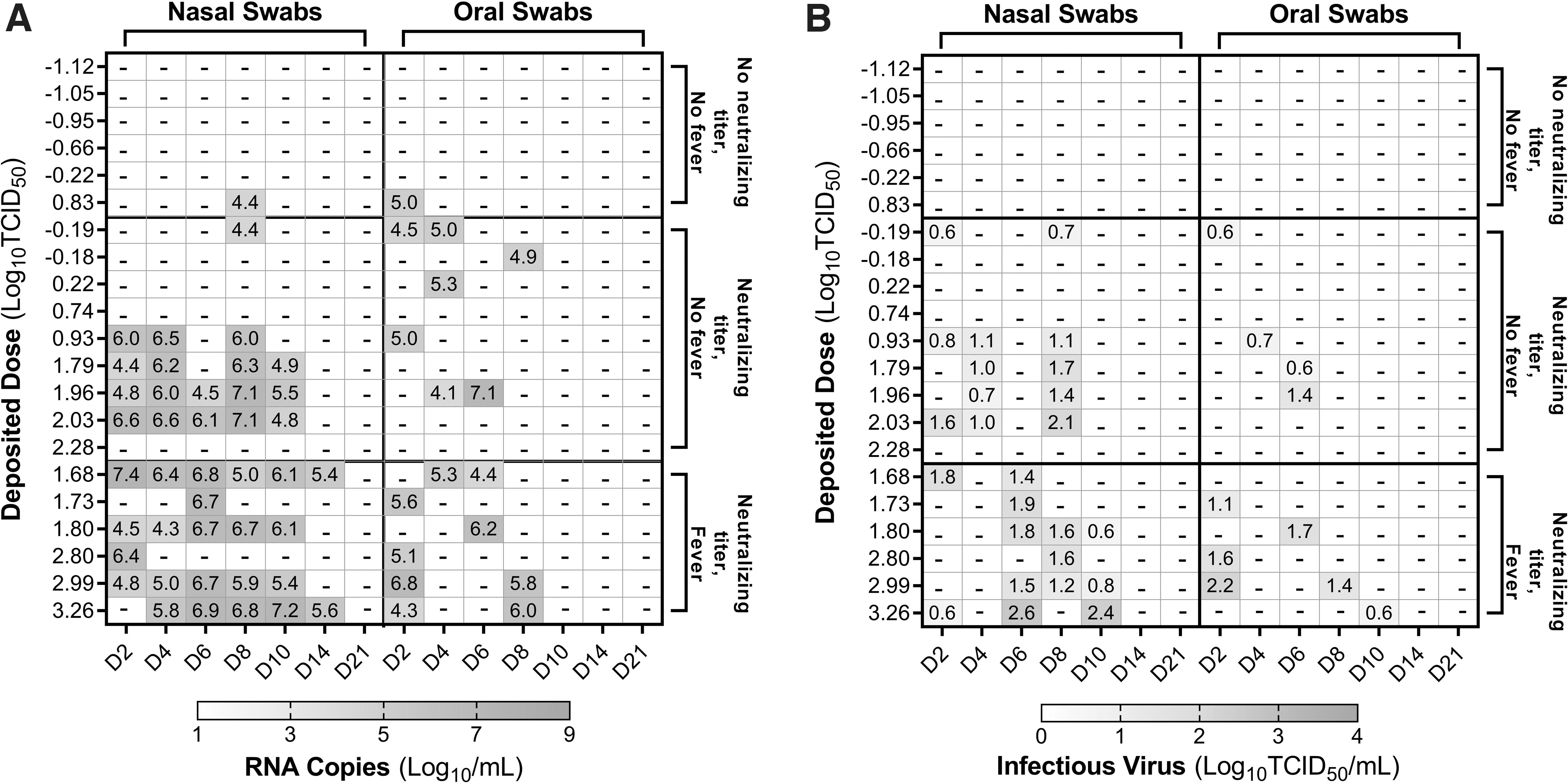
Shedding of viral RNA and infectious virus in nasal and oral swabs from animals exposed to SARS-CoV-2 Gamma. Levels of viral RNA
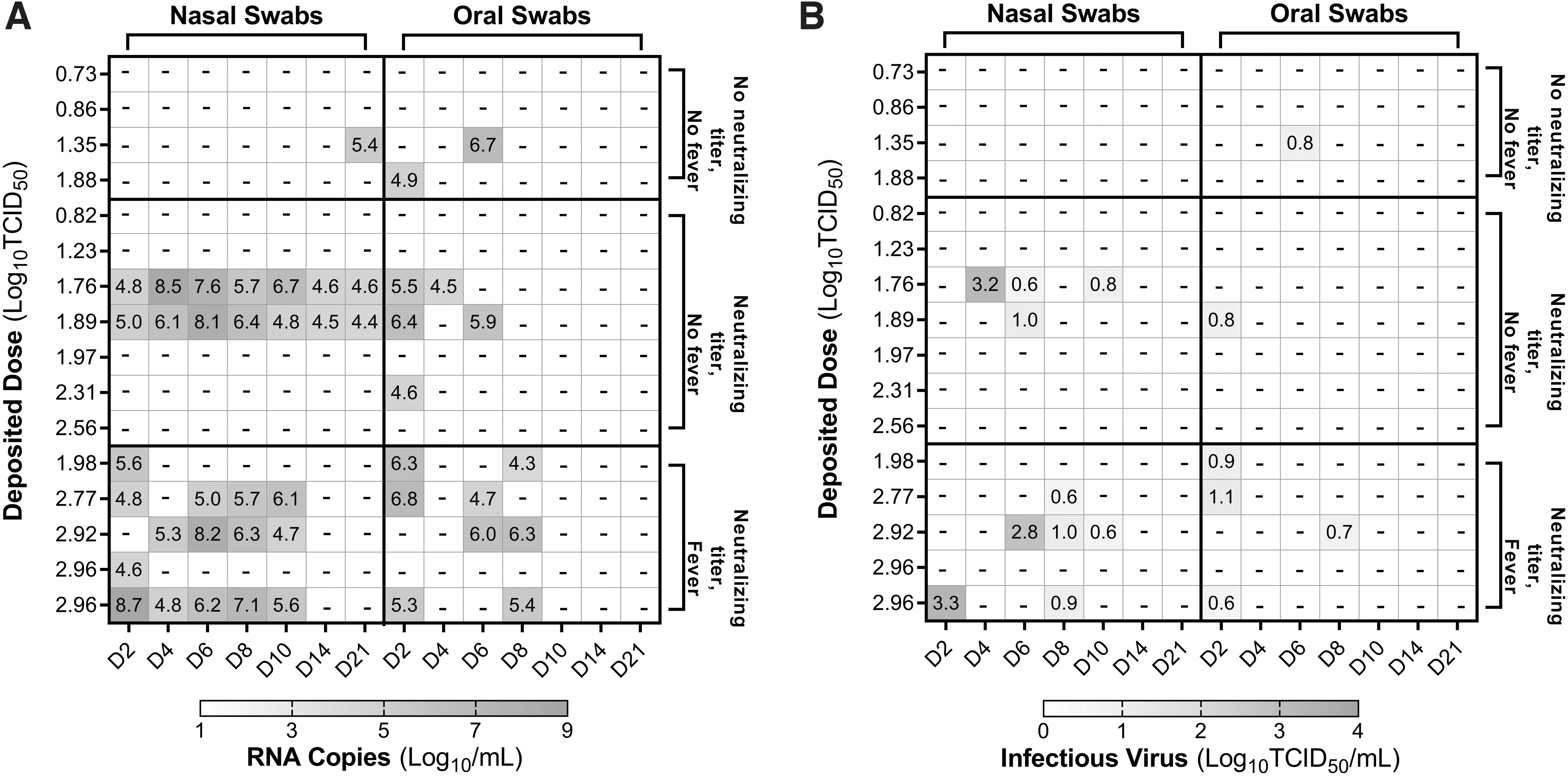
Shedding of viral RNA and infectious virus in nasal and oral swabs from animals exposed to SARS-CoV-2 WA-1. Levels of viral RNA
Comparison of Viral Shedding Metrics for Animals Exposed to SARS-CoV-2 Gamma and WA-1
Mean values compared using unpaired t-tests; Median values were compared using a Kruskal–Wallis test with a Dunn's multiple comparison's posttest.
The median times to peak shedding of infectious virus or viral RNA in swabs were also similar for Gamma and WA-1 (p > 0.40; Table 3). Finally, the median number of days on which infectious virus or viral RNA was detected in swabs postexposure were similar for Gamma and WA-1 (p > 0.99; Table 3).
On day 21 postexposure, 15 of 21 animals exposed to SARS-CoV-2 Gamma had detectable neutralizing titers against the Gamma isolate by PRNT (Table 4). In animals exposed to SARS-CoV-2 WA-1, 12 of 16 had measurable neutralizing titers against WA-1 on day 21 postexposure (Table 5). Probit analysis suggests that viral variant is a significant factor influencing the dose–response relationships for seroconversion (p = 0.033 through LRT). Similarly, the median dose for development of a neutralizing titer was significantly lower for Gamma relative to WA-1 when overlap of 85% confidence intervals was assessed (Table 2).
Summary of Fever and Plaque Reduction Neutralization Test50 Results for Animals Exposed to SARS-CoV-2 Gamma
—, denotes not applicable; HPE, hours postexposure.
Summary of Fever and Plaque Reduction Neutralization Test50 Results for Animals Exposed to SARS-CoV-2 WA-1
Data partially reproduced from Dabisch et al. 20
ND, no data due to malfunctioning telemetry device.
For exhaled breath measurements, the median baseline particle emission rate from all anesthetized animals was 5.6 particles/min, with an interquartile range of 1.4–21.2 particles/min (n = 37). Occasional events, such as a sneezes or movement of mouth, produced higher numbers of particles that were detectable in the system, demonstrating that the system is capable of detecting higher concentrations (Fig. 4). Most exhaled particles measured during quiet breathing had diameters <2 μm, although occasional particles as large as 8 μm were detected.
Exhaled particle concentration profile and particle size distribution. The particle concentration profile during a single collection period is shown. Particle concentrations were low for the majority of the recording period. However, the animal sneezed ∼3.5 minutes into the collection period, resulting in a large spike in particle concentration. The particle size distribution associated with the sneeze measured by an APS is also shown. Unfortunately, this event occurred during the preexposure period, and so no virus was detected. However, these data provide confidence that higher particle emission rates are able to be detected in the system. APS, aerodynamic particle sizer.
Viral RNA was detected on filters sampling the exhaled breath at a single postexposure time point in four animals exposed to the Gamma variant, with estimated emission rates ranging from 4.7 to 5.0 log10 RNA copies per minute (Supplementary Data). No infectious virus was detected in the exhaled breath at any time point postexposure. Three of these animals shed viral RNA in the exhaled breath on day 2 postexposure, and viral RNA was also detected in the oral swab on the same day. Viral RNA was detected in the exhaled breath of the final animal on day 6 postexposure, but no viral RNA was detected in swabs on the same day. The particle emission rates measured in these four animals with the OPS did not significantly change over the postexposure period (p = 0.97 using a Kruskal–Wallis test and a Dunn's multiple comparisons posttest; Fig. 5), and were similar to those measured in NHPs that did not develop signs of infection (p = 0.41). Similarly, no changes were observed in the postexposure particle emission rates if the analysis was restricted to animals with fever (p = 0.79; n = 6) or animals that developed neutralizing titers (p = 0.31; n = 15). As with the baseline period, most exhaled particles had diameters <2 μm.
Exhaled particle emission rates.
A previous study suggested a relationship between exhaled particle count and viral RNA load in nasal swabs of infected NHPs. 31 In the present study, a similar relationship was not observed between exhaled particle counts and the viral RNA load detected in temporally paired nasal swab samples across all animals (r 2 = 0.008; n = 39 samples; Fig. 6). Similarly, no relationship was observed if this analysis is restricted only to samples from animals exposed to higher doses that developed transient fevers (r 2 = 0.008; n = 21 samples; Fig. 6).
Exhaled particle emission rates as a function of viral RNA load in nasal swabs for animals infected with SARS-CoV-2 Gamma. No relationship was observed between exhaled particle counts and the viral RNA load detected in nasal swab samples across all animals (black and gray circles; r2 = 0.006; n = 39 samples). Similarly, no relationship is observed if this analysis is restricted only to samples from animals with fever (black circles; r2 = 0.008; n = 21 samples).
For animals exposed to the WA-1 isolate, viral RNA was detected on a filter sampling the exhaled breath in one animal at a single time point 14 days postexposure, with an estimated viral RNA emission rate of 4.8 log10 RNA copies per minute. No viral RNA was detected in swabs from the same day on which viral RNA was detected in the exhaled breath, although significant levels were found in swabs on other days [Fig. 3; Dose: 2.77 log10 median tissue culture infectious doses (TCID50)]. No infectious virus was detected in the exhaled breath at any time point postexposure. Similar to Gamma-infected animals, no changes were observed in postexposure particle emission rates in animals that developed fever (p = 0.9286; n = 5) or in animals that developed neutralizing antibody titers (p = 0.8208; n = 12) following exposure to the WA-1 isolate. Most particles exhaled by WA-1-infected animals had diameters less than 3 μm, although occasional particles with diameters as large as 8 μm were detected.
Discussion
The present study demonstrates that the median deposited doses for viral shedding in nasal swabs were approximately 10-fold lower for an isolate of the Gamma variant of SARS-CoV-2 than the earlier WA-1 isolate. Given that the median deposited doses for fever were similar, this suggests that there is a greater difference between the median doses for viral shedding and fever for Gamma than for WA-1 and potentially an increased range of doses for Gamma over which asymptomatic shedding and disease transmission are possible. These results also complement those of previous studies, which suggested that differences in exposure dose may help to explain the range of clinical disease presentations observed in individuals with COVID-19,20,32,33 highlighting the importance of public health measures designed to limit exposure dose, such as masking and social distancing.
The disease presentation in Gamma-infected animals was mild and consistent with previous studies that have examined the disease course following inhalation of aerosolized SARS-CoV-2.20,27,34 Animals exposed to higher doses of Gamma developed a transient fever, with an onset between 1- and 3-day postexposure, which resolved within hours of onset. Gamma-infected animals with fever also displayed a transient increase in the IL-6 levels, and a transient lymphopenia that resolved by 4-day postexposure. While IL-6 and lymphopenia have previously been reported to correlate with disease severity in hospitalized COVID-19 patients,35,36 the transient nature of the responses in the present study are suggestive of mild disease. No other changes were observed in parameters reported to correlate with disease severity, including neutrophils, neutrophil-lymphocyte ratio (NLR), blood urea nitrogen (BUN), creatinine, prothrombin time (PT), and activated partial thromboplastin time (aPTT),37–39 again suggestive of mild disease.
While the dose of Gamma required to induce viral shedding in the upper respiratory tract was lower than that of WA-1, the amounts of RNA and infectious virus detected in swabs were similar for both variants. This is similar to another study in NHPs, which compared disease presentation following intranasal infection with different variants of SARS-CoV-2. 40 However, there is evidence that the magnitude of viral shedding in humans may vary for different SARS-CoV-2 variants. One recent study reported differences in the levels of infectious virus detected in nasopharyngeal swabs from individuals infected with different variants of SARS-CoV-2, including Delta and Omicron. 41 Similarly, several studies have reported increased levels of viral RNA in nasal swab samples for more recent variants.13,17,18,42 Another recent study reported that a small cohort of patients infected with the Alpha variant shed greater levels of viral RNA in the exhaled breath relative to other less-transmissible variants. 3
The reason for this difference between swab samples collected from NHPs and humans in these studies is unclear but may be at least partially due to differences in the study populations. Studies with NHPs utilized healthy adult animals that developed mild, predominantly asymptomatic disease, whereas human samples were collected from symptomatic patients.
It should be noted that while data from oro- and nasopharyngeal swabs provide some insights into viral shedding dynamics in infected animals, it is unclear how such data translate to the potential for disease transmission. A recent study reported no correlation between viral RNA levels measured in swabs to those measured in exhaled breath samples in COVID-19 patients, suggesting that viral RNA load in nasal swabs may not be an appropriate surrogate for shedding in the exhaled breath during quiet breathing. 2 This result is not surprising, as previous studies utilizing chemical analysis and breathing maneuvers have demonstrated that particles exhaled during quiet breathing originate in the deep lung, and not the upper respiratory tract.29,43–46 It is known that a fraction of particles emitted from the respiratory tract during speaking or coughing originate from the oral cavity. 29 Thus, the viral load detected in oral swabs or saliva samples may better correlate with the potential for aerosol and/or droplet transmission of disease during speaking or coughing. However, additional studies are needed to better characterize these relationships.
SARS-CoV-2 RNA was detected in five filter samples of the exhaled breath from five different animals—four exposed to the Gamma variant and one exposed to the WA-1 isolate. The emission rates of viral RNA from these animals were between 6 and 7 log10 RNA copies per hour, similar to those reported previously in humans. 4 Unfortunately, a comparison of the levels of viral RNA detected in the exhaled breath to those detected in nasal swabs of NHPs similar to that reported previously for humans 2 was not possible due to the low number of paired positive samples. It should be noted that depressed respiration due to anesthesia has the potential to affect exhaled particle counts, as generation of exhaled particles is known to vary with the degree of respiratory effort.47,48 Therefore, the particle counts measured in the exhaled breath in the present study are likely lower than would be measured in awake, freely moving animals, and this represents a potential difference that complicates the comparison between exhaled breath data from conscious human patients and anesthetized NHPs.
A previous study reported increases in exhaled particle counts postinfection in NHPs exposed by inhalation to the WA-1 isolate of SARS-CoV-2, as well as a correlation between exhaled particle counts and viral loads measured in nasal swabs by PCR. 31 However, similar relationships were not observed in the present study. 31 The reasons for these differences are unclear, as both studies employed comparable methodologies. In the present study, performance data for the exhaled breath collection system provide confidence that the system provided accurate measurements of particle emission from infected animals. Specifically, the system was operated at a slightly positive pressure to prevent infiltration of ambient particles from the animal holding room, resulting in background particle counts near zero despite the high particle concentration in the room.
Additionally, the median pressure in the mask with an animal present was similar to that observed with the sealed mask (+0.1″ water column), providing an indication that the seal on the mask was not leaking with an animal present. Finally, while most animals had relatively low particle emission rates, several animals generated significantly higher levels of particles, in some cases due to sneezes or oral movements, demonstrating that higher particle emission rates are detectable in the system.
While the present study reports novel dose–response data for two isolates of SARS-CoV-2, numerous knowledge gaps remain. The present study and others20,27,31,34 utilize aerosols with aerodynamic diameters in the 1–3 μm range, representative of the size distribution of particles exhaled during quiet breathing.29,44,46 However, larger particle sizes are possible during other respiratory activities, such as speaking or coughing. 29 Previous studies have demonstrated that particle size alters regional deposition within the respiratory tract, and that deposition patterns within the respiratory tract influence dose–infectivity relationships and disease presentation.49–53 Therefore, studies examining dose–response relationships for SARS-CoV-2 in larger particle aerosols are needed to more fully understand the hazard posed by aerosol transmission.
Similarly, studies examining the influence of comorbidities, age, and other risk factors on the dose–infectivity relationships for SARS-CoV-2 are needed as epidemiological data suggest that significant differences in morbidity and mortality exist between some groups.54,55 Finally, while numerous studies have reported the presence of SARS-CoV-2 in the exhaled breath and air samples,3,4,6–8 data on the magnitude of shedding over time are limited. Therefore, additional studies are needed to better understand shedding dynamics in infected individuals.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was funded by the National Institute of Allergy and Infectious Disease (NIAID) under Interagency Agreement (IAA) #20056-001 between NIAID and the U.S. Department of Homeland Security Science and Technology Directorate (DHS S&T). Work was performed at the National Biodefense Analysis and Countermeasures Center (NBACC), a Federally Funded Research and Development Center. Funding from NIAID was provided through DHS S&T under Agreement No. HSHQDC-15-C-00064 awarded to Battelle National Biodefense Institute (BNBI) by DHS S&T for the management and operation of the NBACC. This research was supported in part by the Intramural/Extramural research program of the NCATS, NIH. The views and conclusions contained in this document are those of the authors and should not be interpreted as necessarily representing the official policies, either expressed or implied, of DHS, the National Institutes of Health (NIH), or the U.S. Government. The DHS does not endorse any products or commercial services mentioned in this presentation. In no event shall the DHS, BNBI or NBACC have any responsibility or liability for any use, misuse, inability to use, or reliance upon the information contained herein. In addition, no warranty of fitness for a particular purpose, merchantability, accuracy or adequacy is provided regarding the contents of this document. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Reviewed by: Philip Kuehl and Margaret Pitt
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.