
Abstract
Heteroxylans were solubilized from the alcohol-insoluble residue of switchgrass, rice, Brachypodium, Miscanthus, foxtail millet, and poplar with 1 M KOH. A combination of enzymatic, chemical, nuclear magnetic resonance (NMR), mass spectroscopic, and immunological techniques indicated that grass arabinoxylans have comparable structures and contain no discernible amount of the reducing end sequence present in dicot glucuronoxylan . Our data suggest that rice, Brachypodium, and foxtail millet are suitable model plants for developing technologies to modify arabinoxylan structure and biosynthesis that can be extended to monocot energy crops.
Introduction
The model plant Arabidopsis thaliana has been used extensively to identify and functionally characterize genes involved in plant cell wall biosynthesis. 4 The availability of a large number of mutant lines affected in cell wall-related genes has also provided an opportunity to determine the roles of cell wall polysaccharides in plant growth and development. 4 Nevertheless, Arabidopsis is a dicot and last shared a common ancestor with the grasses (Poaceae) ∼200 million years ago. Moreover, the cell walls of dicots and grasses differ. 5 Thus, Arabidopsis is not a good model plant for studying genetic traits or cell wall structure and synthesis in grasses.
A good model grass should be easy to grow, have a short life cycle, and reproduce by self-fertilization. Its genome should be sequenced and well-annotated. The plant should be easily, rapidly, and stably transformed, and mutant lines available and accessible. Moreover, the composition and structure of the biomass from the model plant and the energy crop should be comparable. Three plants, rice (Oryza sativa), Brachypodium (Brachypodium distachyon), and foxtail millet (Setaria italica), have been promoted as model plants for understanding growth, photosynthetic efficiency, and cell wall biosynthesis in grasses. Rice and Brachypodium are diploid C3 plants with sequenced genomes, and in-bred and mutant lines are available. Brachypodium, with its small size and fast generation time, is a good model for functional genomic studies and for the dissection of more highly conserved traits in bioenergy grasses, including genes involved in cell wall synthesis. 6 Foxtail millet is a diploid C4 plant with a sequenced genome and is closely related to switchgrass. Thus, foxtail millet, is a good model plant for the genetic dissection of traits in bioenergy grasses that carry out C4 photosynthesis, as well as for studies of plant architecture and cell wall synthesis. 3
Arabinoxylan accounts for ∼20% of grass biomass. Some of the arabinose side chains contain ester-linked ferulic and coumaric acids that participate in the cross-linking of arabinoxylans and in the formation of ferulate-polysaccharide-lignin complexes that cross-link the cell wall. 7 Such ferulate-polysaccharide-lignin complexes are hypothesized to function in numerous processes in plants. 7 These complexes together with the interactions between arabinoxylans and cellulose are believed to be the important factors in the recalcitrance of biomass to conversion to sugars. 8 In addition, degradation of xylan and other hemicelluloses during biomass pretreatment may result in the formation of compounds that are toxic to the microorganisms used to deconstruct the biomass and convert the released sugars to liquid fuels. 9 Thus, there is considerable interest in understanding arabinoxylan structure and how it can be altered to improve the bioprocessing characteristics of grass biomass. This paper focuses on the comparison of structures of the carbohydrate moieties of grass arabinoxylans and does not address the structures of the phenolic esters linked to this polysaccharide.
Arabinoxylans have a backbone composed of 1→4-linked β-
Materials and Methods
Plant Material
Stems and leaves of mature Miscanthus x giganteus, B. distachyon, and S. italica were obtained from the Plant Biology greenhouse at the University of Georgia (Athens, GA). Rice straw was obtained from Dr. Wayne Parrot at the University of Georgia. The air-dried stems and leaves were Wiley-milled (−20/+80 mesh) and stored at room temperature. Milled poplar and switchgrass were obtained from the National Renewable Energy Laboratory (Golden, CO). The switchgrass (P. virgatum cultivar Alamo) was grown in Ardmore, OK by The Samuel Roberts Noble Foundation milled using a Hammer mill with a 1-in screen, and then ground in a Wiley mill using a 1-mm screen. The milled material was sieved to −20/+80 mesh. A hybrid poplar (Populus trichocarpa x deltoides) tree was harvested at the Oak Ridge National Laboratory (Oak Ridge, TN) and cut into short logs. The logs were debarked, split with an axe, and chipped using a Yard Machines 10HP chipper (MTD, Cleveland, OH). The chips were then milled in a Model 4 Wiley Mill (Thomas Scientific, Swedesboro, NJ) using a 1-mm screen size. All the material was then sieved (−20/+80 mesh) using a W.S. Tyler Sieve RX-29 Rotap (W.S. Tyler, Mentor, OH).
Preparation of Cell Walls as Alcohol-Insoluble Residues
Cell walls of switchgrass, rice, Miscanthus, Brachypodium, and foxtail millet were prepared as their alcohol-insoluble residue (AIR). 11 The Wiley-milled biomass from all five grasses was ball-milled for 16 h at 4°C and 90–100 rpm in aq. 80% ethyl alcohol (EtOH) (v/v) using ¼-in zirconium grinding media (U.S. Stoneware, East Palestine, OH). The insoluble residues were washed with acetone and dried under vacuum.
Sequential Extraction of Cell Wall Glycans from Air
AIR (0.5 g) was sequentially extracted with 50 mM ammonium oxalate (35 mL), 1 M KOH containing 1% (w/v) NaBH4, and 4 M KOH containing 1% (w/v) NaBH4. The insoluble residue remaining after treatment with 4 M KOH was then delignified using sodium chlorite and acetic acid. 12 The delignified AIR was then extracted with 4 M KOH containing 1% (w/v) NaBH4. 13 The 1 M KOH, 4 M KOH, and post-chlorite 4 M KOH extracts were neutralized with glacial acetic acid if required, dialyzed against deionized water (3,500 molecular weight cut off tubing), and lyophilized.
Glycosyl-Linkage Composition Analysis
Glycosyl-linkage composition was analyzed by generation of partially methylated alditol acetates. Oligosaccharides (∼1 mg) were methylated as described. 11,14 The methylated oligosaccharides (∼200 μg) were then hydrolyzed for 2 h at 120°C with 2 M trifluoroacetic acid (TFA). The released glycoses were converted to their corresponding alditols by treatment with NaBD4 and then treated for 20 min at 50°C with acetic anhydride-TFA (1:1 v/v) to generate the partially methylated alditol acetate derivatives. The partially methylated alditol acetates were separated and quantified using a Hewlett Packard 5890 gas chromatograph coupled to a Hewlett Packard 5870 mass selective detector (Agilent, Santa Clara, CA) as described. 15
Generation of Xylo-Oligosaccharides
Xylo-oligosaccharides were generated by endoxylanase treatment of the 1 M KOH-soluble material. Suspensions of the 1
MALDI-TOF Mass Spectrometry
Positive ion matrix-assisted laser-desorption ionization-time of flight (MALDI-TOF) mass spectra of xylo-oligosaccharides were recorded using a Bruker LT MALDI-TOF mass spectrometer interfaced to a Bruker Biospectrometry workstation (Bruker Daltonics, Billerica, MA). Aqueous samples (1 μL of a 1 mg/mL solution) were mixed with an equal amount of a matrix solution (0.1 M 2,5-dihydroxybenzoic acid in aq. 50% MeCN) and dried on the MALDI target plate. Spectra from 200 laser shots were summed to generate each mass spectrum. 15
1H-NMR Spectroscopy
Xylo-oligosaccharides from the five grasses and poplar (∼3 mg) were dissolved in D2O (∼0.5 mL, 99.9%) and 1H-NMR spectra were recorded with a 600 MHz Varian Inova NMR spectrometer (Agilent, Santa Clara, CA). Two-dimensional spectra were recorded using standard Varian pulse programs. Chemical shifts were measured relative to internal acetone at δ 2.225.
Glycome Profiling
Glycome profiling of cell wall extracts was performed as described, using a collection of plant cell wall glycan-directed monoclonal antibodies (mAbs). 16 –18
Results and Discussion
To compare the arabinoxylans from the five grasses, AIR was prepared from mature leaves and stem tissue and then sequentially extracted with ammonium oxalate, 1 M KOH containing NaBH4, and 4 M KOH containing NaBH4. The insoluble residues were delignified with chlorite-acetic acid and then extracted with 4 M KOH. Glycosyl residue composition and glycosyl-linkage composition analyses established that most of the solubilized xylan was present in the 1 M KOH extract (Table 1). Indeed, 1,4 linked and 1,3,4 linked xylosyl residues were predominant in the 1 M and 4 M KOH extracts from the five grasses, suggesting the presence of branched arabinoxylans in these wall extracts.
Glycosyl-Linkage Compositions of Material Solubilized by 1 M and 4 M KOH Treatments of the Grass Biomass
T-Araf is non-reducing terminal arabinofuranosyl.
Comparison of Structures of the 1 M KOH-Soluble Arabinoxylans
To facilitate comparison of arabinoxylan structures, the 1 M KOH-soluble arabinoxylans from each of the grasses were treated with an endo-β-1,4-xylanase to generate xylo-oligosaccharides. These oligosaccharides were isolated by size-exclusion chromatography (SEC) and then structurally characterized using MALDI-TOF-MS and 1- and 2-dimensional 1H-NMR spectroscopy. The glucuronoxylans from dicots including Arabidopsis and poplar have the glycosyl sequence [4)-β-
MALDI-TOF-MS showed that the most abundant xylo-oligosaccharides generated from the grasses and poplar have molecular masses between 700 Da and 1,400 Da (Fig. 1). The predominant ions ([M+Na]+ and ([M-H+2Na]+) in the grass spectra were consistent with the presence of oligosaccharides composed mainly of pentosyl (Xyl and Ara) residues (Fig. 1A-E). In grasses, the major ion at m/z 701 corresponds to [M+Na]+ of an oligosaccharide composed of five pentosyl residues, consistent with the presence of arabinoxylan. The most abundant poplar oligosaccharides are substituted with a MeGlcA residue to give monoisotopic [M+Na]+ ions at m/z 759, 891, and 1023 and monoisotopic [M-H+2Na]+ ions at m/z 781, 913, and 1045 (Fig. 1F). The monoisotopic ions at m/z 759 and 891 correspond to oligosaccharides that are composed of four xylosyl residues substituted with a MeGlcA and five xylosyl residues substituted with a MeGlcA, respectively. These ions also correspond to monoisotopic [M+Na]+ ion of the sequence [β-
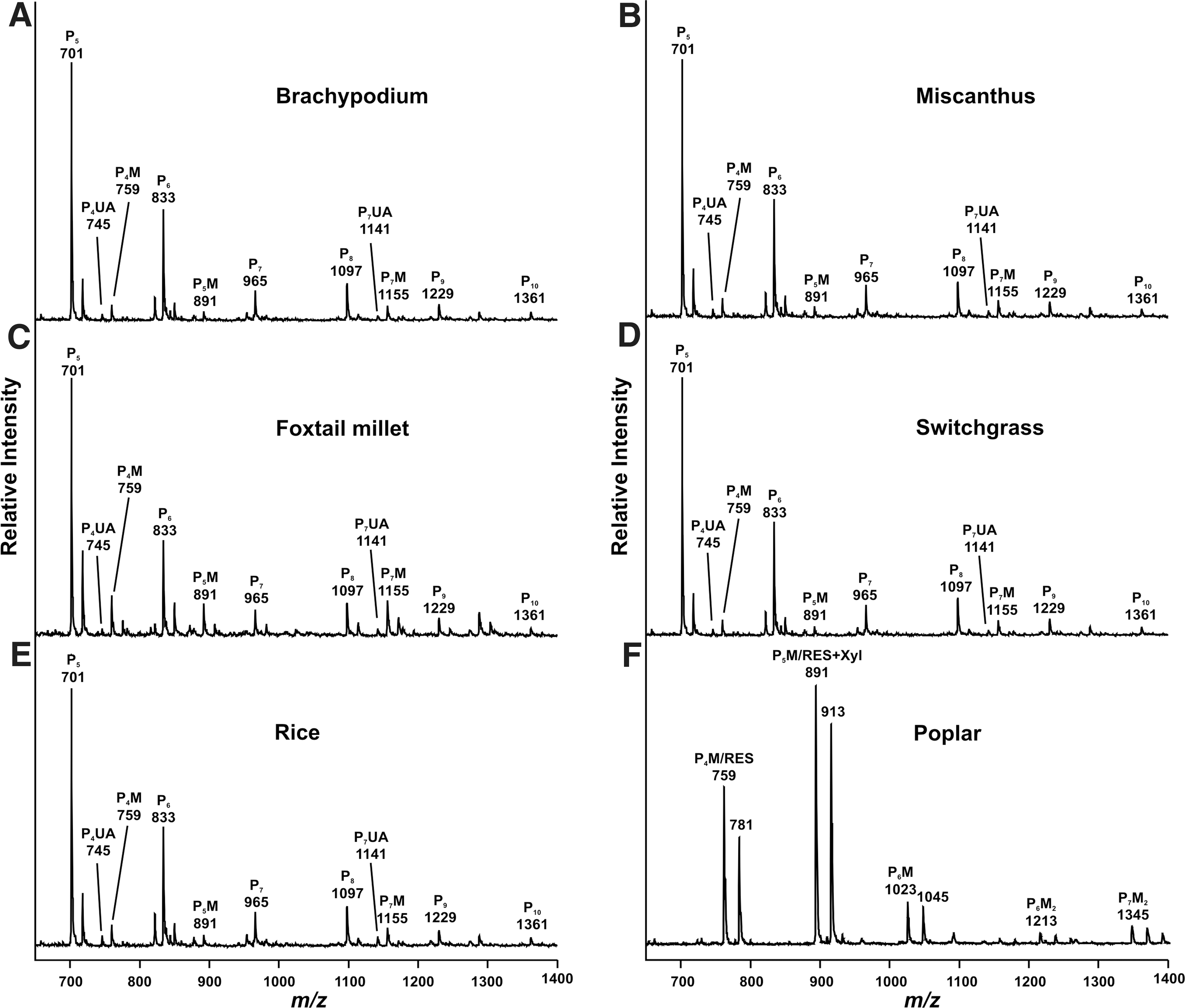
MALDI-TOF spectrum of the purified xylo-oligosaccharides generated by xylanase treatment of the 1 M KOH extracts grasses
To confirm and extend the results obtained by MALDI-TOF-MS, the endo-xylanase-generated xylo-oligosaccharides from the grasses and from poplar were further characterized using 1H-NMR spectroscopy. The chemical shifts and scalar coupling constants for the protons of each glycose were assigned based on 2D gradient correlation spectroscopy (gCOSY) analyses (Fig. 2) and published data.
11,15,21,22
Signals were assigned for the anomeric and ring proton resonances for the terminal non-reducing β-
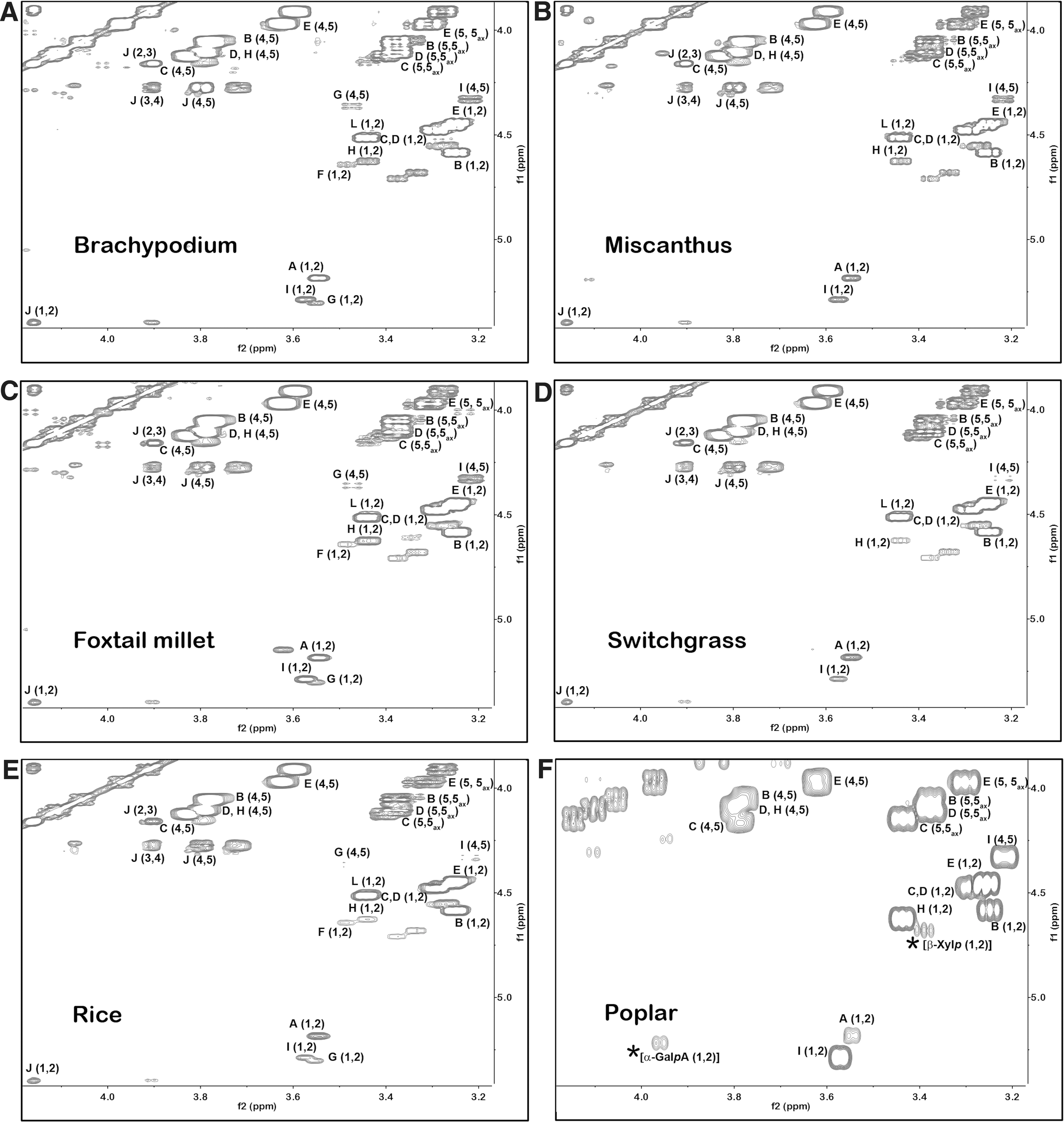
Partial 600-MHz gCOSY NMR spectrum of purified xylo-oligosaccharides generated by β-endoxylanase digestion of the 1 M KOH extracts of AIR from grasses
1H-NMR Assignments of the Xylo-Oligosaccharides Generated by Endoxylanase Treatment of the 1 M KOH Extract of AIR From Five Grasses and Poplar
Chemical shifts are reported in ppm relative to internal acetone, δ 2.225. β-Xyl (α-GlcpA) is a β-linked xylosyl residue that bears a GlcpA sidechain at O-2. β-Xyl (α-L-Araf) is a β-linked xylosyl residue that bears a α-L-Araf sidechain at O-3. H-4 and H-5 of the reducing α-xylose were not assigned. The number of decimal places for each chemical shift reflects the accuracy of the measurement. Residues are indicated by an uppercase letter as a key for cross referencing with Figure 2.
The signals for 4)-β-
Glycome Profiling of Grass Biomass
Glycome profiling utilizing a suite of mAbs that recognize diverse cell wall polysaccharide epitopes was used to determine the major types of polysaccharides present in the grass cell walls and how readily they are solubilized by aqueous buffer and by alkali before and after delignification.
18
The AIR isolated from switchgrass, rice, Brachypodium, Miscanthus, and foxtail millet was sequentially extracted with 50 m
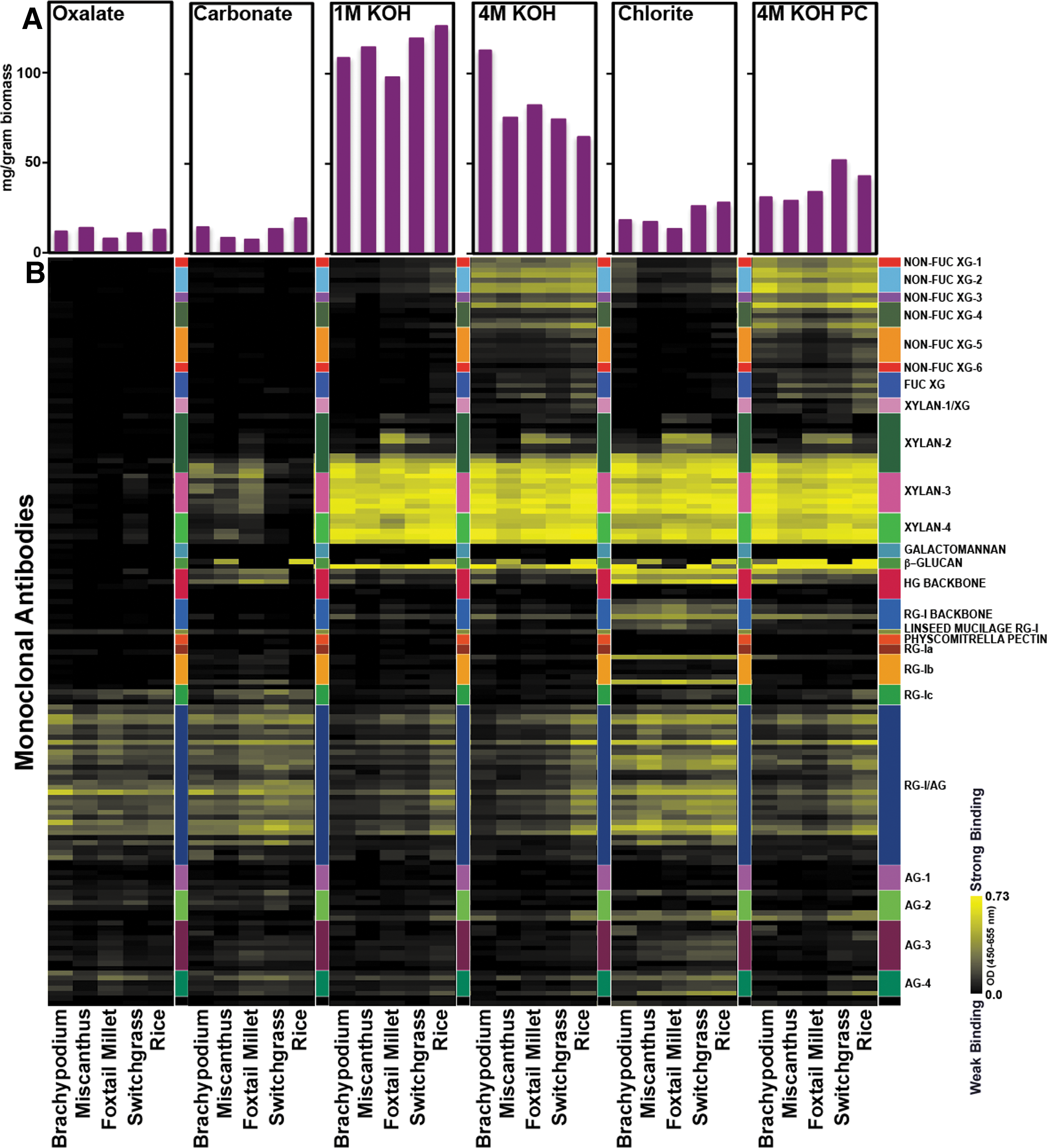
Glycome profiling of five grass cell wall extracts. The extracted AIR by various reagents were loaded onto ELISA plates and were screened against an array of plant glycan-directed monoclonal antibodies. 16 The panel on the right of the figure lists the individual antibodies used, grouped according to the polysaccharides predominantly recognized by these antibodies. Antibody binding is represented as a colored heat map, with black signifying no binding and bright yellow representing the strongest binding. The bar graphs at the top indicate the amount of solubilized material recovered at each extraction step per gram of AIR.
Glycome profiling showed strong binding of all of the xylan-3 and xylan-4 groups of mAbs and some of the antibodies in the xylan-2 mAbs to the 1 M KOH, 4 M KOH, chlorite-acetic acid, and post-chlorite 4 M KOH extracts. These results are consistent with published reports that the cell walls of grasses contain arabinoxylan. 11,23,24 In addition, strong binding of the mAb BG1, which binds to (1→3,1→4)β-glucans, was observed in the alkali and acidified-chlorite extracts. 25 Miscanthus and rice also showed binding of the mAb LAMP, which recognizes (1→3)β-glucans (callose) in these same extracts. 26 The presence of the mixed-linkage β-glucan in the cell walls of mature grass tissue was somewhat unexpected, as this polysaccharide is typically associated with the cell walls of rapidly growing tissues. 27 Likewise, the presence of detectable amounts of callose in the extracts from some grasses was unexpected, as callose deposition is typically associated with wound responses or pathogenesis. 28
In all five grasses, the binding of mAbs that recognize epitopes other than xylan and β-glucan was comparable. For example, xyloglucan epitopes were more abundant than pectic/arabinogalactan epitopes (HG backbone, RG-I backbone, RG-Ia, -Ib, -Ic, RG-I/AG, AG-1, AG-2, AG-3 and AG-4 groups of mAbs) in 4 M KOH and post-chlorite 4 M KOH extracts. The opposite was true in the acidified chlorite extracts. Interestingly, xylan, pectin, and pectic arabinogalactan epitopes were abundant in the acidified chlorite extract suggesting that some of the xylan and pectin are associated with lignin in grass biomass. Together, our results suggest that the cell walls of the five grasses studied contain similar polysaccharides, although there are subtle differences in their glycan epitope compositions.
Conclusions
We have used a combination of chemical, spectroscopic, and immunological techniques to show that similar pectic and hemicellulosic polysaccharides are present in biomass from switchgrass, miscanthus, rice, brachypodium, and foxtail millet. The quantitatively predominant hemicellulose is arabinoxylan. Some subtle structural differences, including the degree of backbone branching of the xylan backbone and the extent of binding of xylan-directed antibodies, were observed. Our study provides evidence that grass arabinoxylans lack discernible amounts of the distinct reducing end sequence of glycoses that is present in glucuronoxylans from hardwoods and softwoods. Nevertheless, the high degree of structural similarity for arabinoxylans from rice, brachypodium, and foxtail millet suggests that these three plants provide useful models for identifying fundamental aspects of xylan biosynthesis in commercial grass bioenergy crops, including Miscanthus and switchgrass.
Footnotes
Acknowledgments
This research was funded by the BioEnergy Science Center (BESC), administered by Oak Ridge National Laboratory, and funded by a grant (DE-AC05-00OR22725) from the Office of Biological and Environmental Research, Office of Science, US Department of Energy. The generation of the CCRC series of plant cell wall glycan-directed monoclonal antibodies used in this work was supported by the NSF Plant Genome Program (DBI-0421683). We would like to acknowledge Maria J. Peña of the CCRC for discussions with respect to NMR spectroscopy.
Author Disclosure Statement
No competing financial interests exist.