
Abstract
As the most versatile and precise gene editing technology, prime editing (PE) can establish a durable cure for most human genetic disorders. Several generations of PE have been developed based on an editor machine or prime editing guide RNA (pegRNA) to achieve any kind of genetic correction. However, due to the early stage of development, PE complex elements need to be optimized for more efficient editing. Smart optimization of editor proteins as well as pegRNA has been contemplated by many researchers, but the universal PE machine’s current shortcomings remain to be solved. The modification of PE elements, fine-tuning of the host genes, manipulation of epigenetics, and blockage of immune responses could be used to reach more efficient PE. Moreover, the host factors involved in the PE process, such as repair and innate immune system genes, have not been determined, and PE cell context dependency is still poorly understood.
Regarding the large size of the PE elements, delivery is a significant challenge and the development of a universal viral or nonviral platform is still far from complete. PE versions with shortened variants of reverse transcriptase are still too large to fit in common viral vectors. Overall, PE faces challenges in optimization for efficiency, high context dependency during the cell cycling, and delivery due to the large size of elements. In addition, immune responses, unpredictability of outcomes, and off-target effects further limit its application, making it essential to address these issues for broader use in nonpersonalized gene editing. Besides, due to the limited number of suitable animal models and computational modeling, the prediction of the PE process remains challenging.
In this review, the fundamentals of PE, including generations, potential, optimization, delivery, in vivo barriers, and the future landscape of the technology are discussed.
INTRODUCTION
Gene editing, a promising avenue for treating hereditary disorders and modifying genomes, initially used DNA insertion as the primary method to deliver genes into the target sites. 1 Subsequent advancements in the 2000s introduced technologies such as zinc-finger nucleases, transcription activator-like effector nucleases, and integrase-homing endonuclease fusion proteins, enabling targeted modifications within the genome. 2 –4
The development of CRISPR-associated (Cas) nucleases, base editors, and prime editors ushered in a new era, allowing precise editing of mammalian cell genomes across a diverse range of target sites. This progress stems from the groundbreaking discovery of guide RNA-programmable CRISPR systems. 5 –8 With the recent FDA approval of the first CRISPR-based product, Casgevy, which is targeted to hematopoietic stem cells for the therapy of β-thalassemia together with sickle cell disease, turned major interest to this technology for curing hard-to-treat genetic disorders. 9
Although these technologies share fundamental principles, their potential, efficiency, and limitations show variations, forming the context of our discussion.
The pivotal breakthrough in gene manipulation occurred in 2012 with the discovery of CRISPR-Cas9. Typically, different Cas nucleases induce a double-strand DNA break (DSB) at a designated genomic location. A guide RNA (gRNA) that comprised 18–24 nucleotides (nt) and complementary to one strand adjacent to the target site directs the Cas nuclease to cause changes. The Cas-gRNA complex is activated upon detecting a protospacer-adjacent motif (PAM) and binds to the corresponding DNA target sequence. PAM sequences vary depending on the bacterial origin of the Cas nuclease; for example, SpCas9, derived from Streptococcus pyogenes, recognizes the PAM 5′-NGG-3′. 5,10 –13 Following recognition, Cas9 cleaves 3 nt upstream of the identified PAM. 14
As nucleases do not naturally repair the broken sequence of DNA, the cell will then carry out mechanisms, such as nonhomologous end-joining (NHEJ), homology-directed repair (HDR), and microhomology-mediated end-joining (MMEJ), to fix the changed sequence (Fig. 1A). 15,16 NHEJ often results in error-prone nucleotide insertions or deletions, leading to gene knockout. 17 MMEJ, however, could cause unintended insertions, deletions, or translocations, 18 whereas HDR, relying on homologous donor DNA, ensures precise repair and gene replacement (17,18). Because the majority of these insertions or deletions (indels) result in frameshifts in the targets’ coding sequences, Cas nucleases are excellent tools for causing gene disruption (Fig. 1A). 15 Despite the potential of Cas nucleases for gene disruption, the resultant indels are challenging to predict or control. 19 –21 Finally, the plasmid donor-templated HDR is inefficient, single-stranded DNA HDR can just knock in small sequence (<50 bp), and MMEJ is also not so effective in mammalian cells. 22
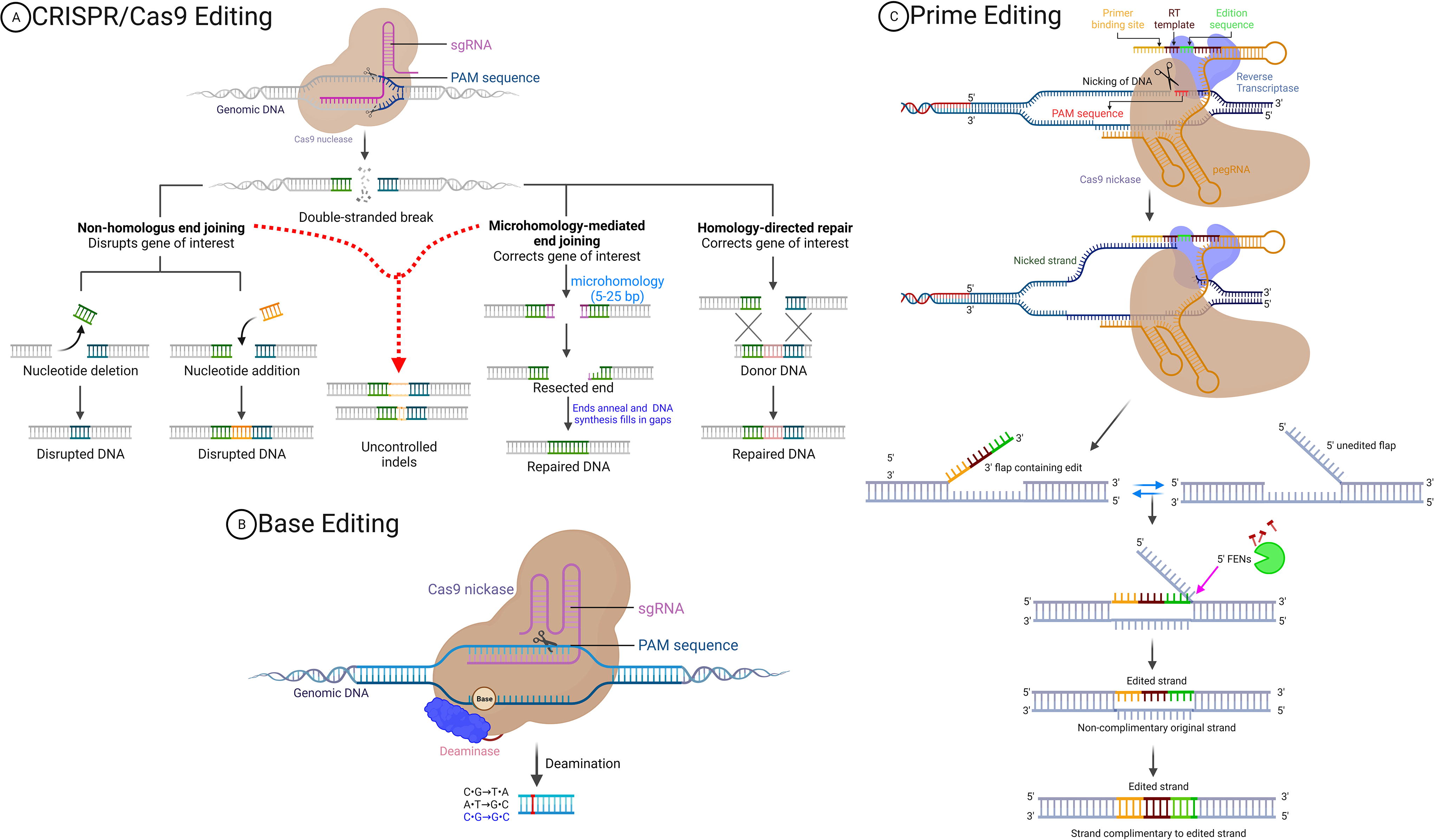
Graphical description of the Cas9-mediated genome editing technologies.
DSB-FREE TECHNOLOGIES FOR PRECISE GENE CORRECTION: LEVERAGING HDR FOR SPECIFIC INSERTIONS AND ALTERNATIVE APPROACHES
Codelivering nucleases with an exogenous donor DNA template, flanked by homologous sequences, allows specific DNA alterations. However, HDR activity in mitotic cells is often surpassed by end-joining, leading to indel by-products, and compromising editing accuracy. 23,24 An alternative approach involves homology-independent targeted integration, where DNA donors lacking homologous sequences are inserted at DSB sites through NHEJ. However, this method lacks control over insertion orientation and number, with undesirable indels frequently outnumbering intended edits. 25 Moreover, DSBs generated by nucleases can trigger p53 activation, massive deletions, chromosomal translocations, and chromothripsis, potentially resulting in transformed cells. 26 –30 In light of these challenges and unintended effects associated with nucleases, the exploration of alternative technologies has gained momentum, aiming to achieve precise gene correction without DSBs.
BASE EDITING TECHNOLOGY
Base editing (BE) has emerged as a consequence of advancements in the CRISPR-Cas9 technology, driven by the demand for minute yet precise gene modifications (Fig. 1B). This technology encompasses three variations: cytosine base editors (CBEs), adenine base editors (ABEs), and C to G base editors (CGBEs), each designed to facilitate single-nucleotide conversions. 31 –33 ABE induces A> G and T > C conversions, while CBE introduces C > T and G > A conversion. On the contrary, CGBE generates C-to-G transversions, although with lower average efficiencies and product purities. 31,34,35 These editors convert all targeted base pairs within a specific window to their counterparts, such as the CBE transforming all C•G base pairs into T•A base pairs.
The advantages of BE over CRISPR-Cas9 lie in its independence from an exogenous donor DNA template and avoidance of a DSB and cell cycle. Utilizing a modified D10A nickase Cas9 minimizes undesired indels and mitigates the adverse effects associated with DSBs. 26,28,30,36 –40 The precision of BE allows specific targeting of a single codon, enhancing accuracy in mutation correction. Comprising a programmable DNA-binding protein (e.g., a transcription activator-like effector repeat array or a catalytically impaired Cas nuclease) linked to a deaminase enzyme, base editors operate under the guidance of the guide RNA (gRNA) to locate a specific genomic sequence. 31,32,36,41 –43 The Cas protein domain displaces single-stranded DNA (ssDNA) at the target site and activates the tethered enzyme, leading to the specific deamination of the accessible target nucleotide (Fig. 1B). 31,32,42 –44
While both base editors and Cas9-based editing rely on gRNA/target sequence recognition, base editors typically achieve higher precision by directly modifying individual nucleotides without inducing DSBs or requiring extensive repair processes. Despite these advantages, BE has limitations. Undesired nonsynonymous mutations may occur if a nondesired base-pair change in the activity window occurs. 45 Bystander editing may also occur when neighboring C or A nucleotides undergo conversion due to the canonical deamination of similar nucleotides within a limited activity window of 4–5 nt. Although base editors mainly induce synonymous mutations within a typical editing activity window, preventing unwanted BE of neighbor nucleotides can be challenging. 46 The targeting scope of the Cas domain, reliant on the presence of a PAM sequence within a specific distance range (usually 15±2 nt) from the target nucleotide, further constrains BE activity. However, there are new versions of BEs which are less dependent of PAM, through protein engineering of Cas9. 47 In addition, certain base editors may inadvertently cause unintended changes in both DNA and RNA. 48 –50
Although engineering efforts have mitigated some of these limitations, contemporary base editors can only generate six of the 12 different types of point mutations, leaving other potential DNA changes, such as insertions, deletions, and most transversions, beyond their reach. 51 In summary, while base editors offer advantages over classical Cas9-based editing, including precision and the creation of many point mutations, they are hindered by a small editing window and the potential for bystander editing near the target site. Table 1 provides a comprehensive comparison of all Cas9-mediated editing strategies.
Comparison of the Different Aspects of the Main CRISPR-Based Edition Strategies 52
PRIME EDITING TECHNOLOGY
Liu’s group first proposed prime editing (PE), combining the potential of retroviral reverse transcriptase (RT) with Cas9 nickase activity in response to the necessity for precise editing. By using a guided Cas9 nickase and RNA template-directed reverse transcription, they introduced different types of genetic mutations at several loci without DSBs. 53 In the first step, they constructed the initial prime editor (PE1) by fusing a SpCas9 nickase to the wild-type RT from Moloney murine leukemia virus. 53 The nickase does not harbor an active HNH domain due to a critical H840A substitution, so only the nontarget strand three nucleotides upstream of the PAM site is cleaved without leaving a DSB.
A lengthy tripartite prime editing guide RNA (pegRNA) includes a classical gRNA, a scaffold, and a variable length 3ʹ extension region, favor settling down the PE machine on the right target sequence. Similar to classical CRISPR gRNA, a 20-nucleotide length gRNA directs the Cas9 to find the target loci. Similarly, a universal scaffold sequence interacts through its 3-dimensional stem-loop structure with Cas9 protein domains to enter the pegRNA into the editor machine. The 3ʹ extension contains at least two parts, including an RT enzyme template (RTT) with a template for editing enclosed by the left and right complementary sequences, and a primer binding site (PBS). The RTT sequence is a provided RNA template for the reverse transcription process; thus, any desired changes should be designed there beforehand. Its left and right arms provide microhomology flaps to facilitate the final edited strand ligation. The PBS is complementary to the target sequence and gRNA is the shortest part of the pegRNA. Its attachment to the PAM strand will initiate the priming step. As its attachment triggers the PE process, it has a crucial role in on-target editing efficiency. After the PAM cleavage, the attached PBS provides a priming point for the extension of the target sequence by RT polymerization. 53 Due to the natural affinity of pegRNA and Cas9/RT fusion protein, they assemble in the nucleus and initiate the “search and replace” editing process. Cas9/RT/pegRNA assembly prevents spacer exonuclease degradation, due to the Cas9 masking activity, however, the 3ʹ extension still is posed to degradation. 53 The 3ʹ extensions anneal to the target point through the PBS, after which the spacer gRNA screens sequences and guides nCas9 to locate on the non-PAM strand to cleave the PAM site on the target strand. The editor machine cleaves the target strand at three nucleotides upstream of the PAM sequence, designated as the 0 position, to allow for the true numbering of the surrounding nucleotides as + for upstream or − for downstream. In the priming step, PBS is still annealed to the target sequence, the resulting nick at the end of PBS exposes a 3′-OH end, which primes the RT enzyme to copy the RTT and extend a new synthesizing target strand (Fig. 1c). Due to prime extension, PAM cleavage, and strand displacement, DNA polymerization at the prime site results in either 3ʹ or 5ʹ overhangs, microhomologous flaps. The 3ʹ flap with the desired edit equilibrates with the original unedited 5ʹ flap. If the 5ʹ flap is excised by host endonuclease, the 3ʹ flap containing the intended edits and PAM strand homology arms anneals with a nonedited strand (Fig. 1c) to produce a heteroduplex DNA with edits. 53 Conversely, ligating the naturally displaced 5ʹ flap into dsDNA can happen by excising the 3ʹ flap, which eliminates the edits. An extra nick positioned far from the target site, upstream or downstream on nontarget strand, induces the repair system to copy the inserted edit in the target site to the nontarget strand, after which the 3ʹ flap has been fixed into the heteroduplex DNA form. The induction of repair system to install the edit on the nontarget strand by an additional gRNA leads to the development of PE3 and PE5 generations. 51
COMPARING PE WITH BE AND CRISPR TECHNOLOGIES
The discovery of an exciting CRISPR-based system and its applications for knockin/knockout genes in human cells led to the development of the most accurate editing methods, PE and BE. These editing methods differ in their potential, specificity, breadth of activity, safety, and limitations. In the CRISPR-Cas9 method, heterogenous DSBs, which are generated by nucleases, can result in large mutations, including deletions, translocations, and chromothripsis. These by-products of DSBs trigger different DNA damage sensors, contribute to cellular stress responses, and culminate in the activation of cell cycle blockers and checkpoint markers such as p53.5,54 In addition, strategies such as inhibitor 53 (i53) have been developed to mitigate the unwanted effects of DSBs. 55 It is worth noting that while DSBs are also involved in BE and PE, they occur at lower levels compared with the CRISPR-Cas technology. 32,53 Moreover, the recent approval of an NHEJ-based gene editing drug product underscores the potential of DSB-based approaches in therapeutic applications. 56 Although the HDR strategy induces less dramatic changes in DNA strands, the consequences of repair are still unpredictable and vulnerable to large deletions and mutations. 51 By harnessing the targeting ability of CRISPR-Cas systems, CRISPR-associated transposons also enable researchers to insert, delete, or modify genes at specific locations in the genome with unprecedented precision. 57 Similarly, homing endonucleases have the ability to induce targeted DNA DSBs, which can be harnessed for precise genome editing. Unlike traditional CRISPR-Cas systems, which rely on RNA guides to specify the target sequence, homing endonucleases directly recognize DNA sequences, offering an alternative approach to genome editing with potential advantages in terms of specificity and targeting range. 58 Moreover, homing endonucleases can be engineered to recognize novel target sequences, expanding the repertoire of possible genomic modifications. 58,59
Due to the unsafe activity of wild-type Cas9 and the potential for significant sequence edits by this nuclease, new BE and PE techniques emerged with a more precise single-nucleotide modification. Table 1 provides a brief comparison of various aspects of all Cas9-based editing strategies. In contrast to CRISPR-Cas9, BEs are a precise method for changing nucleotides with substantially greater efficiency. Theoretically, it might result in a few indel bystander changes and a more controlled edition. Most genetic diseases arise from single-nucleotide changes, making them good candidates for small modifications by BEs instead of the harsh DSB protocol. However, similar bases in the small window of base edition (4–5 nucleotides) are subjected to unintended conversion, leading to bystander mutations. In most cases, this undesired change has a silent effect, but if the bystander edit affects a nonsynonymous amino acid or stop codon, it could significantly impact the cell. In addition, BEs can induce off-target mutations in DNA strands, although much less frequently than the CRISPR-Cas9 method. The nonspecific activity of deaminase has been shown to induce Cas9-independent bystander mutations in DNA and, interestingly, in RNA. 60 PAM dependency in BEs is another limitation that restricts the targeting scope of the machine to a maximum of 15 nucleotides from a PAM site. Finally, this method permits only six out of 12 possible point mutations without the capability for insertion and deletion of any nucleotides.
PE is the most optimal strategy for precise genome editing, owing to a set of advantages over other methods. Most genetic diseases are related to small mutations in the genome; therefore, CRISPR-Cas9 might not be so effective in correcting the sequence without unwanted indels. The range of BEs is also limited to tiny mutations. With PEs, precise genome manipulation in 12 possible point mutations, insertions, and deletions is practical due to the designed RTT in the pegRNA backbone. Moreover, multiple base substitutions at different positions inside the PE window (around 33 bp from the PAM site), medium-size insertions (<40 n), large deletions (up to 100 n by classical variants and up to some kbs by dual-pegRNA variants), and the combination of some genetic alterations are applicable using generations of PEs. The larger the window size, the greater the targeting scope of PEs when compared with BEs. In other words, the manipulation achievable by PEs cannot be reproduced by BEs and more importantly will override the potency of changes by Cas9-mediated HDR. Through its scope, it is suggested that nearly 90% of all human genetic defects could be targeted by PEs. Despite gRNA containing only a 20-nucleotide spacer sequence for the specificity of CRISPR-Cas9 and BEs, pegRNA contains the following three elements: spacer, RTT, and PBS that hybridize to double-stranded DNA, making the cleavage reaction much more specific in PEs. However, bystander mutations could arise due to the genetic context, rate of efficiency, and suboptimal activity of the editor proteins in mammalian cells. Moreover, as PEs do not need DSBs or HDR, they can introduce any kind of mutation to any cells at the G0 or G1 state of the cell cycle, and without cycle arrest. 51 Overall, the on-target frequency of edition by CRISPR-Cas9 and BE systems is high but variable (from 1% to 50%) 61 in human cells and mice, while for Cas9-mediated HDR and PE, it is lower. However, in a similar mouse model to human Leber congenital amaurosis split, AAV-PE2 showed a 28% editing frequency (the highest frequency in a mouse model reported so far), whereas the correction efficiency of Cas9-mediated HDR and BE was 1.2% and >11%, respectively.
PE OFF-TARGETS
The frequency of on-target modifications by PE is generally lower than other CRISPR-based technologies, and several investigations showed the higher frequency of the next generations, PE3Max and PE5Max. However, frequency is vastly different from cell to cell and tissue to tissue. Among the CRISPR-based gene editing technologies, PE has the lowest probability of off-target mutations. In a pioneering study, the rate of off-targets in 293T cells at endogenous loci in HEK3, HEK4, EMX1, and FANCF genes was determined to be <0.1%, <2.2%, <0.1%, and <0.13%, respectively, compared with the traditional Cas9 method with a 4–48% off-target frequency. In addition, rare PE-induced bystander mutations were confirmed in different cell lines, mouse embryos (<0.1%), 60 and organoid cultures (undetectable), indicating the precise activity of the PE machine. Furthermore, in the pioneer work of Anzalone et al., the ratio of correct edition to indel mutations was estimated to be 30-fold higher than in the HDR-mediated edition. 53 However, continuous reverse transcription over the 3′-extension of the pegRNA might copy a part of the scaffold structure and insert it into the target site. 60 Ectopic expression of an exogenous viral RT in the nucleus may also induce PE-independent reverse transcription and unwanted consequences. Genome-wide screening, however, revealed no significant PE-independent off-target variations due to this kind of activity. Although off-target effects have been reported, whole-genome sequencing (WGS) of four prime-edited clonal organoid lines revealed no detectable genome-wide off-target edits. 60 In response to an increasing demand for the standard construction of PE systems, a wide range of easily accessible online resources and applications for customized pegRNA design and off-target site prediction have emerged. 62 In the case of the Cas9 nuclease, a couple of valid and advanced methods are available for identifying off-target locations. Genome-wide identification of DSBs made possible by sequencing, GUIDE-seq, CIRCLE-seq, and Multiplex Digenome-seq, recently upgraded to screen sequences for the majority of Cas9 nucleases. 63
The options for determining the by-products from Cas9-induced DSBs are varied, but they are not entirely suitable for PE. The development of nickase-based Digenome-seq (nDigenome-seq) to assess PE2’s genome-wide specificity has come as a new choice. The PE-tag method, based on amplification tag insertion at the site of prime editor activity, can accomplish genome-wide profiling of off-target sites by PE in vitro and in vivo. 62 Several traditional algorithms, such as Cas-OFFinder, off-Spotter, CasOT, and CROP-IT (for improved off-target binding and cleavage-site predictions) have also been shown to be applicable for PE screening, in addition to WGS and whole-transcriptome sequencing. 64 Interestingly, to enhance its computational prediction strength, CROP-IT has been incorporated with chromatin state data from 125 human cell types to boost prediction potency for PE protocol based on the epigenetic criteria rather than only DNA sequence information. 65
The application of aforementioned methods is of great value, however, long pegRNA interactions with transcriptome in vivo as well as proteome of cell are hardly predictable due to hundreds of 3D structures that RNA can take.
THE EMERGENCE OF RECENT PE GENERATIONS
After the discovery of the most precise strategy of “searching and replacing,” PE, many efforts were made to improve the initial version. Although the primary wild-type PE1 machine theoretically has the potential to create all 24 possible mutations alongside the genome, low efficiency, typically <5% at the targeted allele, and the possibility of off-target effects were significant drawbacks (8). To enhance efficiency, new generations and variants of PE have been developed by modifying (A) the PE editor machine, (B) protocol of pegRNA/sgRNA combination, (C) pegRNA structure, and (D) regulation of host factors such as the repair system, as depicted in Fig. 2.
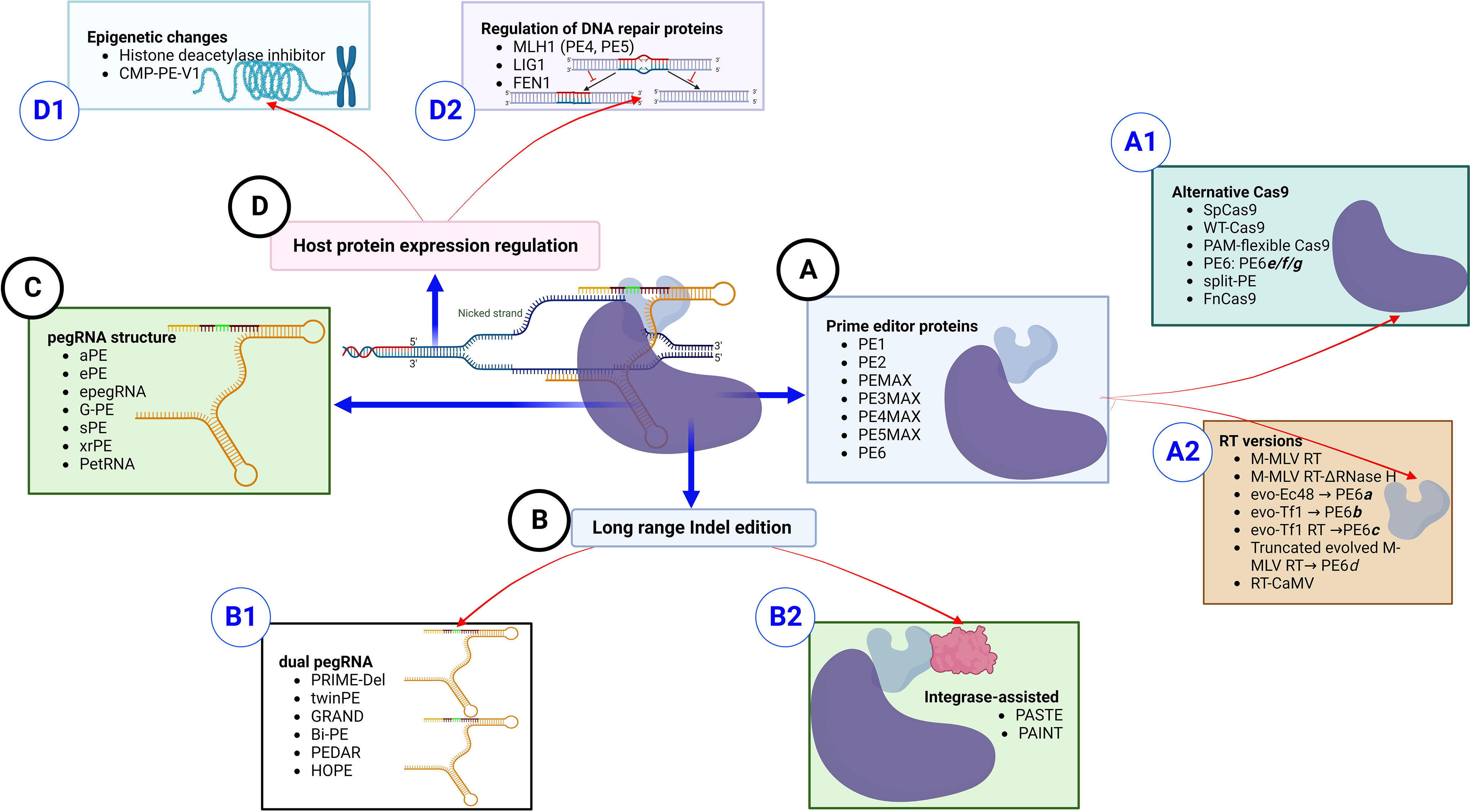
Schematic view of the possible ways to optimize prime editing. 62
As the main PE generations that come from Liu’s group, PE2-PE6 have been developed through fine-tuning modifications to the amino acid sequence of MuLV-RT, nCas9, and using an additional sgRNA. They first introduced five amino acid mutations (D200N/L, T306K, W313F, T330P, and L603W) into the MuLV-RT structure of PE2 to improve its DNA-RNA hybridization strength, thermostability, and enzyme processivity. Even though PE2 has increased efficiency, it still depends on an endogenous repair system to establish an edition at a non-PAM strand of the loci. Therefore, to enforce the repair machine to copy the edited sequence on the nonedited strand, PE3 with an extra sgRNA was included to provide a distal nick to prompt the repair system to copy the edit on the non-PAM strand. 66 The additional sgRNA complementary to the non-PAM strand was demonstrated to be useful when located ±40-100 bp down- or upstream of the edition site; however, the optimum distance and direction need further evaluation. Even though PE3 efficiency was surprisingly high (up to fourfold of PE2), the higher frequency of bystander variation is still an issue. The second nick by sgRNA might increase the rate of indels due to DSBs and overfunctioning of the repair system. 67 Therefore, a version of PE3 and PE3b, with a matched sgRNA to the edited point of the target sequence, was optimized for better specificity. Contrary to the expectation, the superiority of PE3 over PE2 is not always practical for all target cells, and in some cells such as Hematopoietic stem cells (HSCs), PE2 could be wiser to apply. 68 To further optimize the strategy, a new improved PE2 version, PEmax, with a different RT/Cas9 codon usage, increasing the number of NLS signals, and modification in the composition of the peptide linker, was developed (Fig. 2A). They showed that among >20 variants evaluated, the best efficiency was achieved by the PEmax architecture containing a new human codon-optimized MuLV-RT, a new Cas9 (R221K and N394K mutated), a 34-aa peptide linker with a bipartite NLS from SV40, and an additional C-terminal c-Myc NLS. 67 The PEmax version showed superior activity compared with the original architecture of the PE2 and PE3 generations when tested in several loci in HeLa cells. 67 Currently, the PEmax version combined with enhanced pegRNAs (epegRNA) is used as the gold standard for any newly developed version of editing. Nelson et al., designed a new form of pegRNA with an incorporated structural RNA motif at the 3′-terminus. The addition of this element to pegRNA 3′-end improves structural stability and reduces degradation by nuclear exonucleases. They demonstrated that the PEmax-epegRNA version increased efficiency three- to fourfold in different cell lines and primary human fibroblasts without affecting the rate of bystander mutations. 69
Regarding the large size of the editor protein, in several studies, shorter versions of proteins have been developed. In miniPE, the best example, the new nCas9 protein, CjCas9 (H559A) nickase, and an RNase-H truncated MuLV RT were designed in a minimal length of 4.5 kb suited to the capacity size of the AAV vector. 70 Then, the developed PE RT2.10 (F155Y)-Cj was packaged into a single AAV9 for the delivery of mini-PE2 and mini-PE3 to mouse retinas in vivo, but with very low efficiency (<1%). The role of RNaseH was determined to be dispensable for RT during the PE process; however, its deletion makes RT inadequate for hard-to-edit cases and long-range modifications. In a similar effort, Schwanks group, through phage-assisted continuous directed evolution (PACE), developed a more flexible and efficient enzyme, evoCjCas9, to expand the PAM compatibility. In this study, they packaged the new compacted editor machine, CjCas9-PEΔRnH, into a single AAV to base edit different sites in mice, which had encouraging editing efficacy (up to 30%), especially for noncanonical PAM sites. 71
Most recently, Liu’s group has developed the new compacted version of PE2, designated as PE6 generation, by phage-assisted RT domain evolution and nCas9 domain engineering, which could outperform the previous PEmax version. The variants of the RT domain, PE6a-d, and the engineered domain of nCas9, PE6e-g, showed a different efficiency but enough versatility to accomplish editing through different designs regarding size constraints, pegRNA structure, and delivery methods. The evolutionarily constructed versions of RTs, evoEc48 and evoTf1, showed a great increase in the efficiency of the PE method for hard-to-edit points while reducing the size of editors. Finally, an in vivo experiment on the mouse brain showed efficient long insertion potency (40%) in the brain cortex by dual AAV-PE6d, which is substantially higher than with PEmax. 72
OTHER DEVELOPED PE VARIANTS
nCas9 versions with higher PAM flexibility
To increase the coverage area for PE function, new versions of the nCas9 enzyme with less PAM dependency have been investigated. The range of PE activity from the PAM site is <40nt, and in the best condition, it is <10nt, which is a limitation, especially for PE3b. To expand the range, new types of Cas9 variants with non-NGG PAMs or alternative nuclease sites have also been explored (Fig. 2A1). Several studies have described a set of engineered Cas9 enzymes capable of detecting alternative PAMs. 60 Some studies have also developed compact and less PAM-dependent versions of Cas9 that are suitable for efficient packaging into small-capacity vectors. Fortunately, several spCas9 PAM variants have been successfully developed for PEs to broaden the strategy scope. At least three spCas9 PAM variants, including PE-SpG (PAM: NGH), PE-NG (PAM:NG), and PE-SpRY (PAM-independent), have been integrated into PE strategies and have shown encouraging results. 60 For example, the SpRY Cas9 version extends the coverage of PE for 94.4% of human genetic disorders and increases the applicability of PE3b to more loci. Other Cas9 orthologs, such as SaCas9, SauriCas9, and CjCas9, with unique PAM preferences, are also possible replacements for the current version in cases where PAM accessibility is limited. 60 Recently, other Cas9 orthologs, such as evoCjCas9, have been developed to circumvent PAM-dependency limits and large-size issues. However, PAM-less dependency for this nickase is an issue that should be considered cautiously. 71
MANIPULATION OF CELLULAR FACTORS
Based on the crucial role of the repair system in the PE process, Chen et al. screened the effect of over 450 genes involved in all DNA repair pathways on editing fate using CRISPRi-based screening. They demonstrated that key players in the mismatch repair system (MMR) can reduce PE efficiency by causing the reversion of the intended edit and disrupting the 3′ extension flap. To confirm the significant role of the MLH1 protein in reducing PE efficiency, they developed a new MMR-ablating method by introducing dominant negative proteins (such as MLH1dn) into treated cells. As expected, the next generation of PE, PE4 (PE2max+MLH1dn), and PE5 (PE3max+MLH1dn) systems, based on MLH1 ablation, was shown to be twice as effective as PE3 in MMR-proficient cell types. 67 The use of MLH1dn was shown to increase the rate of editing in several cells, but its overexpression has been found to be less effective in mouse and rabbit embryos for creating animal models. 73
In contrast, inducing the expression of host proteins might also aid the PE process. Xu et al. found that the overexpression of certain repair factors, such as flap endonuclease 1 (FEN1) and DNA ligase 1 (LIG1), could enhance the efficiency of PE, similar to MLH1dn but with less potency. FEN1 cleaves the 5′-flap generated by DNA displacement during the Okazaki fragment extension. 74
To promote the stability of the R-loop at the edited site, Song et al. designed a variant of PE2 containing the Rad51 ssDNA binding protein fused to Cas9 by a linker. The developed PE2 variant, HyPE2, showed enhanced editing compared with the original PE2 generation at endogenous sites. They claimed that the interaction of the Rad51 protein with ssDNA stabilizes the single-strand structure and makes it more available for RT transcription at the R-loop. 75
Moreover, Liang et al. developed new variants of PE2 in plants by adding a 5′-to-3′ exonuclease enzyme sequence to the termini of the Cas9-RT fusion proteins. This design aimed to promote the digestion of the 5′ overhang and keep the 5′-flap from competing with the 3′-flap for hybridization to the genome. 76
Cell death is also a potential factor affecting the efficiency of genome editing. Due to the potential of Cas9 to trigger the p53-dependent apoptosis pathway and cell cycle arrest, this pathway has been validated in PE from the very beginning of the method’s discovery. The transient inhibition of p53 by the dominant negative protein p53DD greatly enhances the editing rate among human pluripotent stem cells (hiPSCs). Therefore, transient inhibition of p53 via the codelivery of p53DD, a dominant negative fragment of p53, significantly improves PE outcomes in generating target mutations in hiPSCs. 77
ENHANCEMENT OF pegRNA STRUCTURE
In addition to the aforementioned variants, some minor modifications have also been introduced into the next variation of PE3 to decrease off-target frequency and control the repair system. Strategic silent mutations complementary to the PAM site or near the intended edit are two types of artificial changes in the RTT to avoid retargeting the PAM site by nCas9 and to prevent the MMR system from reverting the edited sequence back to the original one, respectively. 78
The extension of the 3′ DNA flap into the scaffold by MuLV-RT is another issue associated with PE that might induce the insertion of a homologous part of the scaffold into the edited gene. In practice, disrupting the homology between the scaffold and the target gene could inhibit the incorporation of this sequence into the genome. 67
After the development of efficient Cas9-RT fusion, some studies made efforts to improve the noncoding part of the PE machine, pegRNA. As the key player, pegRNA modification is also addressed to improve structural stability, half-life, and RNA-DNA interaction. In contrast to sgRNA, which is protected by the Cas9 protein, the 3′ extension of the pegRNA is exposed to nuclease enzymes. Nelson et al. described pegRNA 3’-end modification through their engineering by the addition of viral or synthetic structured RNA motifs. They showed that two structured RNA motifs, including evopreQ1 (prequeosine1-1 riboswitch aptamer) and knot (frameshifting pseudoknot of MuLV), are suitable candidates to add at the 3′-end and prevent the degradation of the pegRNA. The engineered pegRNA performed threefold superior activity compared with PE3 in HeLa, K562, U2OS, and primary fibroblast. 79 After this progress, other groups also included different structural RNA motifs to preserve the 3′-end moiety of pegRNA by adding flavivirus-derived exoribonuclease-resistant RNA motifs (xrRNA, xrPE), and human telomerase RNA G-quadruplex (GPE). 80,81 In the GPE variant, the G-quadruplex showed a significant increase over PE3 and less than 0.1% off-target editing. It is noteworthy that the incorporation of other structured RNA motifs, such as a 15-bp hairpin, p4-p6 domain of intron I group, and Csy4 recognition site, also enhances the durability and structural stability of the pegRNA. 81
Recently, Ni et al. tested different RNA motifs at the 3′-end of the pegRNA in wheat plants, and selected evopreQ1 (ePE) for the establishment of a multiplex editing platform. With the new platform, they simultaneously edited several genes in protoplasts and regenerated wheat with a frequency of nearly 75%. 82 However, it should be noted that the result of editing in plants cannot always be translated to animal cells. The addition of other stem-loop structures, such as MS2, PP7, Csy4, and BoxB to the 3′-end of the pegRNA can increase the edition even more compared with the epegRNA method. This strategy is also useful to tether split pegRNA(sPE) to the editor enzyme by a relevant protein linker MS2 phage coat protein. 69
In addition to the end modifications of the pegRNA, artificial small-nucleotide changes throughout the pegRNA could enhance the editing mechanism. For example, Li et al. developed an spegRNA strategy by introducing same-sense mutations (SSMs) at proper positions in the RTT sequence. 78 The logic behind this idea was to evade the indel edition in the target loci from MMR by a few base substitutions. MMR is less efficient in correcting a single base mismatch compared to indel; therefore, putting artificial substitutions in the RTT would hide the main change and ultimately increase the PE3 editing frequency. 83 This study also demonstrated that altering the pegRNA secondary structure could improve the edition efficiency. Fine base modifications in the hairpin loop structure of the scaffold gain more stability and better activity in the PE complex. They assumed that hairpins of the scaffold could be broken up by the interaction with another part of pegRNA; therefore, their stability through the incorporation of an artificial C/G pair (apegRNA) instead of a non-C/G pair would be preventive. 79 Their aPE and sPE strategies showed compromising results when applied to PE2, PE4, and especially in PE3 and PE5 methods. Similarly, in other studies, the structure of the scaffold has been modified to increase stability and remove the termination sequence for Pol-III polymerase. The circularization of pegRNA by hybridization of PBS with the spacer sequence might hamper the correct matching of PBS to the target site. To prevent auto-circularization by these elements, Liu et al. incorporated a Csy4 recognition signal at the 3′-end of pegRNA via the formation of the hairpin structure. 85 Even though this strategy increased the rate of edition in comparison with PE3 > 1.5-fold in different genes, a higher incidence of off-target effects detected suggested that more optimization is required. Also, the incorporation of the Csy4 signal could be useful for multiplexing different pegRNAs or sgRNAs in one cassette with the Csy4-Cas9-RT editor machinery.
EPIGENETIC MANIPULATION
The accessibility of genomic loci for the PE complex is another parameter that can be regulated to enhance the editing process. Park et al., with a vision of opening regional chromatin for better gene targeting, used proximal dead sgRNA (dsgRNA) and chromatin-modulating peptides at various target loci to perform gene knock out in mice. They demonstrated a significant effect of both elements on PE3 edition. 86 Subsequently, Lie et al. conducted research to screen molecules targeting chromatin structure, cell cycle regulators, and the DNA repair system to identify key players in PE. The results revealed that histone deacetylase inhibitor (HDACi) molecules significantly boost the efficiency of PE3 for indel-type modifications at endogenous targets. However, the widespread activity of HDACi cannot be solely attributed to opening chromatin protein due to its other functions. 86 The importance of epigenetics in PE was also highlighted by Kim et al., who showed the impeding effect of regional heterochromatin and local nucleosome occlusion on the target loci during PE. They mentioned that a high chromatin structure hinders sufficient genome editing by hiding the loci from the PE machinery. Despite many successes, accurately predicting the outcome of PE is complicated, partly due to context-dependent mechanisms and chromatin accessibility. 87
LONG-RANGE PE FOR LONG GENETIC CHANGE
A shortcoming associated with the initial versions of PE is their limited nucleotide modification potency, a disadvantage compared with classical CRISPR-Cas9. Long-range PEs are a competent alternative to CRISPR-based gene modification but with higher precision. In this setup, a pair of pegRNAs harboring partial/complete 3′-end homology is designed to broaden the edition size. Using two pegRNAs targeting strands at distinct locations, Choi et al. created a long-range edition technique in a groundbreaking study. In this method, after copying the RTT from both 3′ extensions, the resulting homologies from upcoming sequences can hybridize together, and the original sequence would be removed by the ligation of newly created strands. The PRIME-Del variant can induce deletions up to 10 kb, with an efficiency of <30%, as well as insertions of 3-30 bp. 88 Anzalone et al. designed Twin PE using two pegRNAs for programmable long-range modification, including integration, replacement, and in-genome insertion. In this method, two opposite pegRNAs with complementary 3′-flaps and no homology with target sequences are designed to hybridize following RTT extension. 89 TwinPE supported the deletion of 780 bp in Duchenne muscular dystrophy (DMD), insertion of a 108 bp sequence in endogenous sites, long inversions related to Hunter syndrome. Moreover, it can promote the insertion size up to 5.6 kbp DNA into AAVS1, CCR5, and albumin (ALB) (around 6%) when combined with Bxb1 integrase. 90
In the GRAND-editing method, similar to Twin-PE, the RTT sequences of the pegRNA pair contain the desired changes, which are not necessarily homologous to the genomic target but complementary to each other to resolve the 3′-flap. Therefore, after nicking the strands, new ssDNAs synthesized by RT would hybridize together by the RTT homology parts without competition with genomic strands. 91 The insertion efficiency for 150 bp and 250 bp fragments into the endogenous site was determined to be 63% and <30% by this method, which is the highest rate among paired pegRNAs. Based on the accuracy of the GRAND method, a similar strategy, a homologous 3′-extension mediated prime editor system (HOPE) with similar elements, was then developed. 92
In the bidirectional PE, Bi-PE, system, authors designed a two-way primer editing by engineering two different pegRNAs. In this study, a pegRNA with a homologous arm to the opposite side of the deleting sequence and a second pegRNA positioned near the homology arm was designed to bring a long change into the genome. The homology arm permits the synthesized ssDNA to invade dsDNA at the homology site, thereby improving large fragment deletion. This method is efficient in multiplex base conversions, large-sequence replacement, insertion of LoxP sites, 93 the deletion of thousands of base pairs, and the introduction of 10–100 bp small fragments. In another paired pegRNA approach named as precise and specific deletion and repair, instead of nCas9, a natural Cas9 with DSB inducing potency was used with pegF and pegR. The method was demonstrated to be efficient for the deletion of >10kb and the introduction of 60 bp in endogenous sites. 94 The detection of some unintended indels near the target site is a concern associated with this wild Cas9-related method. To overcome the limitations of gene integration, Abudayyeh et al. described a prime-based gene editing to improve the PASTE method, a predescribed strategy for long gene integration. In this study, a programmable integration was oriented via an nCas9-RT fused to serine integrase. By AAV and adenovirus-vector delivery into the humanized mouse liver, they found that PASTE can integrate as high as 2.5% of the desired sequence into hepatocytes at the ACTB locus. 95 Finally, a highly efficient integration method via a combination of primed microhomology and DSB process was recently developed. They exploited a PE strategy based on priming on DSB for long-fragment knockin approach. In PE-assisted integration method (PAINT), a reverse-transcribed ssDNA with a microhomology domain boosted targeted long knockin several times compared with the HDR-mediated edition. The PAINT 3.0 version showed a suitable (up to 80%) nonviral long-fragment genome targeting in different housekeeping genes and in CAR-T cells. 22
CURRENT CHALLENGES IN PE
Despite the current achievements and the fast-growing area of research, several challenges in PE technology remain unresolved. Due to the early stage of development, the PE complexes need to be optimized. Smart optimization of different PE generations and variants has been considered by many, but a universal PE machine containing all criteria for efficient edition is demanding. In addition, the modification of the process should give the editor complex the capability to be context independent and compatible with endogenous factors to overcome the genomic barriers during the cell cycle. For the PE process, the G1 state has shown to be the most proficient step for cell modification, although the genomic context is hard to control and is unpredictable. 96 Considering the context dependency of this strategy, tuning endogenous host factors to make cells permissive for the PE process will be a critical need in the future. The delivery of the PE complex to cells is a major challenge due to the large size of elements. From the initial version of PE (PE1: >6.5 Kbp) to the enlarged PE4/5 (near 8 Kbp) and the recent compacted MiniPE/PE6 versions (near 5 Kbp), size is beyond the capacity of AAV. This becomes even more important when long pegRNAs and proposed regulatory elements, such as promoters, enhancers, and MLH1dn, are included in the final construct. Viral and nonviral delivery methods have recently been developed for PE delivery to cells, but constructing a universal vector for this purpose remains an issue. 97
Following the delivery with viral or nonviral vectors, the next issue that may arise is the induction of an innate immune system. Basic elements of the delivery cargo and, particularly, components of PE possess pathogen-associated molecular patterns (PAMPs) that are detectable by sensors of the innate and pre-existing adaptive immunity. Sensing PAMPs and antigens may activate various innate and adaptive immune pathways, leading to inflammation, interferon (IFN) secretion, and cellular immunity. This immunity might eliminate the transduced cells, change the tissue milieu toward an inflammatory context, and induce autoimmunity. 98 The humoral and cytotoxic T cell responses are triggered by foreign proteins such as Cas9-RT. 99 In three dystrophic dogs undergoing systemic delivery of AAV-CRISPR, it has been shown that vector delivery elicited a detectable immune response that led to the removal of CRISPR-restored cells together with a remarkable muscle inflammation. 100,101
Even though the rate of bystander error of PEs is negligible compared with the other gene editing technologies, there are some concerns about the continuous activity of Cas9 and RT in the nucleus. In fact, persistent expression of AAV, plasmids, or other types of viral vectors could induce unintended sequences alongside the genome. Different modalities, including the inhibition of nuclear export (96), anti-CRISPRs elements, 102 blocker proteins, such as pomalidomide, and targeted delivery with regulatory elements of expression, are also recognized to control the editor machine for a lower off-target rate. 103
Another issue for PE is the unpredictability of the outcome that heavily depends on the host genetic context, the ectopic activity of two enzymes, the presence of a long pegRNA with unknown interaction with host factors, and an extra sgRNA. With such a complex editor, the outcome would be individualized based on many factors. In fact, the PE outcome is an individualized process that might be completely different among a population even with a similar genetic background, and due to differences in epigenetics.
DELIVERY VECTORS FOR PE COMPLEX
Although PE technology grows fast in different areas, a suitable delivery to the target site is of great concern for clinical applications. Viral vectors are usually efficient delivery tools to transduce most tissue types. Adenovirus, AAVs, and retroviruses have been widely used for CRISPR-based editors in vivo. 66 However, the application of these vectors for PE could be challenging due to the limited capacity. Altogether, the total size of the initial version of PE editor proteins (>6kbp), included by MLH1 dominant negative (MLH1dn, in PE4/PE5 versions), pegRNA/sgRNA cassette could be over 7.5kbp that practically is a great challenge for common vector packaging. 66
A. ALL-IN-ONE CONSTRUCTS
The ideal delivery vector for PE should contain all the genetic codes of editor-coding sequences, and noncoding RNAs. The all-in-one vector expressing all the editing elements would be optimal for clinical applications, but putting different elements under different promoters could be problematic.
A1. VIRAL VECTORS
At the moment, viral vectors are the best-suited delivery method for almost any gene therapy applications. Wang et al. developed a high-capacity adenoviral vector containing all the PE elements in one package. They showed that the Advp.PE2.pegRNA vector permits high-efficiency delivery (90% editing frequency) and screening of PE reagents in cells independently of the replication or transformation status. 104 Despite shortcomings such as immune induction, adenoviral vectors are medium-capacity, nonintegrative, and stable vectors that can transduce dividing and quiescent cells, efficiently. Hence, a couple of nonhuman serotypes showed sustained efficiency for transduction of special cells by specific ligands and after bypassing the pre-existing humoral immunity. Despite this, an immune response toward these vectors is a great challenge in the clinic. Adenoviral vectors are categorized into 3 generations as follows: first (ΔE1 and ΔE3), second (ΔE1, ΔE3, and ΔE4), and helper-dependent (gutless vector), based on the number of deleted genes and their capacity for the gene of interest. From the first to third generation, the transduction efficiency, yield of production, and the system stimulation decrease. However, the capacity (from 7–8 to >35 kbp) and duration of expression (from weeks to months) increase. The most attractive adenovirus vector for gene therapy, the first-generation HAd5, has been used in many clinical trials, and it was recently approved by the FDA for the treatment of nonresponsive bladder cancer. 105 Also, encouraging results have been reported regarding the successful delivery of angiogenesis genes by this vector. 106 However, adenovectors have only been used for PE delivery in a few studies.
To design a one-vector construct, reducing the size of the original elements could be rational. The new versions of Cas9 and RT enzymes with a reduced size have been investigated to replace the original large proteins. 107 However, changing the basic elements of PE should be optimized not to affect the efficiency of the complex. Bock et al. systemically administered HAd-5 encoding PE components as well as U6-pegRNA cassette into newborn mice. To fit better into the adenoviral vector, they constructed a smaller PE lacking the RNase H domain. 66 Then, in a model of phenylketonuria mouse, they edited the deficiency with 58% and 11.1% for Dnmt1 and Pah (phenylalanine hydroxylase) loci, respectively. Therefore, they showed that an adenovector with a size-reduced PE2ΔRnH variant (10% size reduction) can efficiently establish a specific edition in vivo. It is noteworthy to emphasize that the optimal capacity of the first-generation adenoviral vectors is less than the PE3 complex length. Recently, an in vivo PE for the cure of sickle cell phenotype was done by using an HDadenovector, HDAd-PE5max, on mobilized mouse HSCs. This all-in-one vector showed 20.2% T > A correction at the mutation site. 108
Lentiviral vectors are among the candidates for the delivery of the PE cargo in one package to human cells. Yet, given the long-lasting expression of genes by lentiviral vectors, adverse events due to their integration should be considered. Integration-deficient lentiviral vectors should be safer and less oncogenic than original vectors but preventing genotoxicity and potential oncogenicity need to be considered. 109
A2. NONVIRAL DELIVERY
Nonviral delivery of PE by plasmid DNA, RNA-based constructs, and ribonucleoproteins (RNPs) is feasible for all PE generations due to the lack of size limits. However, some challenges, such as the formation of the protein corona, passive sequestration by resident phagocytes, kidney filtration, weak extravasation and penetration into tissue, insufficient endosomal escape, and nonspecific uptake by different immune/nonimmune cells, need to be considered. 110
Plasmids containing all PE cassettes are also practical to manipulate, construct, and use for tissue-specific expression. Due to the lack of a packaging limit, the capacity of naked plasmids is larger than the usual viral vectors. As a result, they are suited for designing all-in-one PE constructs for the early steps of pegRNA screening or even in the clinic. Dirkx et al. constructed PE2Max, PE3Max, and PE4Max-based all-in-one plasmids (pAIO) under the control of the EF1a promoter. The elements of the PE machine assembled inside a 15–17 kb plasmid (pAIO-EF1α-PE2/-PE4max) showed better delivery efficiency as well as editing efficiency in Human-induced pluripotent stem cells (hiPSCs) and 293T compared with the multiplasmid method. Interestingly, the study showed for the first time that without positive selection, a new PE2 plasmid can efficiently introduce an edition in hiPSC. Although naked plasmid could be an appealing tool for gene editing owing to stability, easy design and production, innate immune response, risk of integration, off-target editing, and insufficient efficiency are barriers to in vivo application. 111,112 Most of the research focused on PE strategy has been developed through the DNA plasmid constructs. Another strategy that can circumvent the plasmid delivery challenges is a minicircle. This structure is less immune toxic and long-lasting compared with plasmids, due to the removal of bacterial elements from the backbone. It is unclear if the large size of PE hampers the efficiency of the method or scaling up the minicircles, as no studies have yet been done in this area.
The delivery of proteins or RNPs in place of DNA would be the optimal strategy for giving gene editing agents in a safe manner in vivo. Previous findings have shown that delivering BE RNPs instead of BE-encoding DNA or mRNA results in significantly reduced off-target editing, usually without reducing the effectiveness of the on-target editing. The fast-working manner and safe and transient expression of the PE-RNP compared with plasmids or viral vectors make them very promising for clinical use. Delivering BE RNPs has been shown to lead to fewer off-target variations in different tissues, 113 although the specific targeting of the complex to the cell is impossible. The strategy also showed benefits over the CRISPR-Cas9 delivery with acceptable safety. 114 Due to the feasibility and safety potential of lipid nanoparticle (LNP) particle delivery, a PE3-based synthetic mRNA platform has been used to edit PCSK9 in mice. The chemical modification of synthetic pegRNA was shown to improve the output of edition in an animal model. The higher rate of editing in NOD scid mice revealed the negative impact of immune response over the PE process. 115
The virus-like particles (VLPs) are capsid/envelope structures of viruses without an infectious genome inside. As VLPs are derived from the natural viral components, they harbor many properties of wild viruses, including the encapsulation capacity of different cargos, endosomal escape, efficient intracellular trafficking, and the possibility of retargeting without the hazards of the original viruses. 116 VLPs transiently deliver gene editing agents such as mRNA or protein rather than DNA, which significantly reduces the risk of viral genome integration and off-target effects. Therefore, VLPs are popular tools for safe and efficient cargo transfer into host cells. The combination of mRNA/VLPs or RNP/VLPs could be designed for the all-in-one constructs for the delivery of PEs or BEs. Currently, the most popular VLPs used for PE and BE delivery are based on lenti/retrovirus. Retroviruses have some characteristics suited for VLP construction, including medium-sized capsid, stepwise capsid maturation, applicable pseudoenveloping by other glycoproteins, lack of rigid icosahedral symmetry of the capsid, suitable particle-to-Plaque-forming unit (PFU) ratio, and the well-studied genome. 117 More importantly, retroviral-based VLPs are flexible to receive incongruous proteins in the capsid or on the virus envelope. Therefore, gene editor VLPs can be designed through tethering RNP or protein, Cas9/BE, or Cas9/PEs, at different spots of the gag protein. It had been shown that efficient assembly of a retrovirus capsid/RNP complex occurs by fusing editor elements to the gag polyproteins, then protease cleavage would release the Cas9 complex for further activity. 118 In the packaging step, pegRNA/sgRNA and Cas9/PE or Cas9/BE are copackaged into VLPs due to their natural affinity for each other. 113 The engineered VLPs (eVLPs) based on MuLV can mediate strong, therapeutic levels of gene editing in multiple mouse organs by packaging Cas9 nuclease or base editor RNPs. Based on bottlenecks on VLP efficiency, they optimized the cleavage linker stoichiometry of normal to modified gag, and enhanced the nuclear localization of gag-cargo in producer cells. These eVLPs showed to be efficient for BE and PE purposes in vitro and in vivo. After a local injection into the mouse brain of eVLPs, 5% single-nucleotide edition of bulk cells was detected. Furthermore, a single subretinal injection of eVLPs into a genetically blind mouse model and an intravenous injection for the correction of liver PCSK9 disorder showed the efficiency of eVLPs.
Several VLP-BEs and VLP-PEs have been developed for gene editing based on the retroviral structure. In the past, Lyu et al. created a VLP by designing delicate interactions between an aptamer and an ABP protein to transport the whole editor. 119 Banskota et al. have used a new VLP derived from the friend murine leukemia virus (FMLV) to deliver BE. 113 To enhance particle formation, they increased the ratio of cargo-fused to normal Gag, added a nucleus export signal to enhance cytoplasmic availability of the Gag-cargo for self-assembly, and selected a suitable cleavage site for cargo release. The second type of VLPs for gene edition is based on mRNA delivery, and in this architecture, Cas9/RT editor mRNA cargo needs a packaging signal to incorporate inside the VLP capsid. To direct new mRNA toward VLPs instead of the retroviral packaging signal (Ψ), Pages and coworkers derived a phage assembly system based on the interaction between the MS2 coat protein (MS2cp) and its aptamer (MS2apt). 120 In this method, pegRNA and sgRNA are typically encoded from an integration-deficient lentivirus genome, so RNAs can be packed during VLP assembly.
Although RNP-packaging VLPs showed encouraging results in vitro, the methods showed poor efficacy (<10% editing) in vivo. From several studies on the construction of VLPs for Cas9 and BE protein delivery, the editing efficiency in 293T (up to 90%) or other investigated cell lines was promising but not in primary cells (up to 30%) and particularly in vivo. 97 More recently, Haldrup et al. developed an efficient delivery of SpCas9-derived BE/PE by lentivirus-derived VLPs. In their protocol, RNP packaging in VLP is made easier through special Gag/GagPol protein fusion techniques, and improved lentivirus stoichiometry to allow increased particle yield. The subretinal injection of VLP showed suitable capability to disrupt VEGFA gene in vivo, as the first proof-of-concept. However, the PE edition rate needed more optimization in vitro. 121 The use of VLP derived from domestic retrotransposons’ gag protein, PEG10, has also been considered by Segel et al. It has been shown that this protein complex can bind and secrete its mRNA. Therefore, they engrafted the mRNA packaging signal to adopt PEG10 potency for encapsulation and delivery of mRNA of spCas9 and sgRNA. This human cell endogenous system named selective endogenous encapsidation for cellular delivery is a novel delivery method completely without the needs for nonhuman components. 122 In the recent publication v1.3 PE-eVLPs, a VSV-G pseudotype lentivirus construct developed by modification of protease sensitive regions inside the polyprotein, improvement of PE machinery, pegRAN (epegRNA), and COM aptamer bridling for assembly. This VLP showed more efficient PE efficiency in HEK3 (3.2%) and Dnmt1 (1.3%), respectively, compared with v1.1 PE-eVLPs. The mouse study for delivery into the brain and eye also showed enhanced efficiency but still edition results were to 10%. 123 In mouse models of genetic blindness (with Mfrp and Rpe65 impairment), a single injection of new VLP provided a partial restoring of the visual function.
To have an all-in-one construct, a multiplexing strategy should be adopted to bring all elements together. Although most studies use more than one construct for delivery, Yuan and Gao et al. developed a construct with an array containing tandem transfer RNA (tRNA)-sgRNAs for targeting several targets. This type of array can be used to combine pegRNAs and sgRNAs or other elements in one construct to enhance efficiency, using dual pegRNA for larger editions and also to reduce the size of the PE cassette. 124
B. DUAL/SPLIT STRATEGY, ADENOASSOCIATED VIRUS (AAV) FOR PE DELIVERY
Regarding the family criteria, in the Parvoviridae family, AAVs are the smallest human viruses with a diameter size of >20 nm, T1 nonenveloped capsid architecture with a 5 kb ssDNA genome. Several attractive features make AAV the most popular vector for gene delivery, including structural stability, capsid variety (several serotypes and clades), low immunogenicity, no serious diseases, broad tropism to different cells, FDA approval on some products, easy production process, long expression, and rare genotoxicity. 97 The recombinant AAV vector is produced by removing the rep/cap expressing frame, leaving near 4.7kbp space for putting the cassette of interest inside. 125 AAVs are versatile vectors with tropism to different tissues, and the capsid architecture is tolerant to modification for specified targeting of cells by rational design or directed evolution in vitro. 125 Despite many advantages, AAVs induce interferon response and memory cytotoxic T cell expansion, which might pose a risk to patients.
AAV has been used as the most applicable delivery tool for PEs in vivo in the format of split/dual forms. 126 –129 Despite many advantages, the small capacity of the AAV vector is a challenge for large genes such as spCas9 (>4kbp) and particularly fusion of Cas9/RT. In this method, the assembly of PE components is designed to be fulfilled by trans-splicing of the produced pre-mRNA, intein-based fusion of split protein hybrid formation, or homologous recombination between similar elements at the termini of the cassettes. 130 In the dual strategy, PE components are divided into two separate AAV genomes as amino-terminal (N-PE) and carboxy-terminal (C-PE) parts. 131 After transduction, separate parts of the PE proteins can assemble in cotransduced cells. The pioneer study for PE in adult mice used a dual AAV intein-split PE3 variant with success in different sites. 132 To enhance the edition rate in vivo by AAV dual strategy, size-optimized and codon-optimized versions of the editors have recently been investigated. Zheng et al. and Bök et al. have designed RNaseH-deleted PE with 6% and 14% edition at the liver, respectively, and with low off-target frequency. 126,133 It should be noted that although dual vector delivery is suitable for PE, it is still less efficient than the single delivery, as variable results in animals have been demonstrated. 134
To correct the most frequent phenylketonuria variant in a mouse model, a dual AAV8 method was used to restore the Phe concentration in blood with up to 52%. The data of the study showed up to 52% whole-liver PAH editing. 135 It is suggested that untethered MuLV-RT and spCAS9 proteins from separate expressing vectors function in trans when they are cotransduced in vitro. Grünewald et al. developed the split-PE platform consisting of an engineered PE with an untethered, truncated M-MLV RT lacking the RNase H domain. 136 They showed that separated PE2(RT and nCas9 and pegRNA) components expressed by plasmids and AAV function efficiently in cells. The new Split-PE2 (ΔRnaseH RT) demonstrated a comparable edition efficiency with an alternative split-intein PE2. They also suggested that the separate RT activity might be facilitated by some soluble factors. Nevertheless, designing cassettes with some tethering modalities for RT-nCas9 bond can provide a potent semifused RT structure that can function in cis to the nCas9. Before this study, Liu et al. demonstrated that prime edition was possible even when RT and nCas9 are expressed by two separate RNAs. 137 The delivery of the new split PE expressing plasmid to the mouse liver by a hydrodynamic injection resulted in an edition at the β-catenin gene with a similar efficiency as PE3. Interestingly, they suggest the pegRNA-split method and use of circular RT-PBS (petRNA) template matched to a viral capsid to facilitate the connection between Cas9 and RT, and increase the flexibility and stability of the PE complex through the chaperon-like activity of the virus capsid. 138 The other advantage of the split PE architecture is that it can facilitate the screening of new PE methods. For example, in the Grünewald study, they screened a series of viral and non-viral RTs to determine the most desirable ones. 136 Even though viral AAV vectors are the most popular system for in vivo delivery of PEs so far, the high risk of off-target editing due to long-term expression is concerning. 113 The persistent expression of either nCas9 or RT enzymes and the natural integration potency of the AAV vector might trigger tumorigenic pathways or other unintended outcomes. 113 Although several groups have developed a single AAV vector for packaging of BE cassette with encouraging results, the idea remains to be practical in the case of PE delivery. 128,139
Use of separate naked RNAs by microinjections is a possible way to deliver mRNA/sgRNA/pegRNA into cells and zygotes. This method was utilized to make gene correction by PE3 in a mouse model of thalassemia IVS-II-654 mutation (C > T). The base conversion rate of 14% was achieved with controlled beta-globin expression. 140
PE is in the early phase of development, and the components differ from other editing strategies and testing in vivo has been still limited. Therefore, it is likely that different modes of RNA, DNA, and protein delivery offer great potential for this genome editing strategy in the future. However, further optimization of vehicles for safe and efficient clinical applications is still needed. 66
CLINICAL SCOPE AND LIMITATIONS OF PRECLINICAL MODELS
The data analysis and pegRNA design showed that from pathogenic human genetic variants of ClinVar (n = 69,481), more than 90% are targetable by PE3 generation methods. On the contrary, for the correction of known pathogenic alleles, at least a single pegRNA spacer is available, and an average of 3.7 pegRNAs per site. Moreover, the scope of the PE design for gene correction could be increased by including the PAM-less-dependent variants of nCas9. Regarding this, the potential of PE to target most of the hereditary diseases, including cardiovascular, liver, retinal, and brain genetic diseases with a promising outcome, is significant. 141,142 Even more, the technology could be used for nontherapeutic approaches such as high-throughput screening of variations responsible for tumor progression. 143
Owing to the potency, PE is predicted to reach clinical trials earlier than what has usually happened for other gene therapy protocols. However, the lack of suitable animal models might slow down the process. The use of animal models to assess the efficiency and safety of PE has some limitations such as between-species variation and context dependency of PE. Today, most of the animal studies done for the evaluation of PE have been performed in mouse models. 68,71,108,126,132,135
Differences in epigenetics, tissue regeneration pattern, cell cycling, and physiology would impede easy translation of mouse experiments into large animal settings. The species-related activity of different PE generations has been shown between mouse and rabbit embryos, indicating a high level of variation among species. In addition, MLH1dn-dependency was also shown to be different between mouse and rabbit embryos. 73 Theoretically, patient’s cells engrafted into immune compromised mice would be a possible preclinical model of PE.
As recently reported, ex vivo PE-modified hematopoietic stem cells (HSC) derived from sickle cell anemia patients maintain the sufficient level of globin expression after engrafting erythroblasts and reticulocytes in mice. 144 However, the in vivo edition of mouse models for curing disease might not mimic the real circumstances of human genome. 108,135
Zebrafish is another vertebrate candidate that has been used to create human disease models. PE-RNP complex editing indeed efficiently showed desirable changes in the model. However, the fact that this model is very different from humans might hinder the development of PE models from zebrafish. 84
The safety pattern of PE needs investigation in primate models. The stability of editing and the long-term consequences in large-animal genomes need to be clarified. As an example, even though an LNP-encapsulated BE was approved for a clinical trial in humans for the treatment of familial hypercholesterolemia, it was later stopped temporarily for safety concerns. 68
FUTURE PROSPECTIVE
By the recent development of gene editing, particularly PE, hopes have come up for the elimination of most human genetic disorders. However, several challenges in this technology remain unsolved.
The safety issue for PEs has not been followed up in large animals, with only a few studies in mice. The safety pattern of technology needs more investigation in vivo, particularly on primate models. Because of the novelty, the stability of editing and long-term consequences in the large animal genomes need to be clarified. As an example, although an LNP-encapsulated BE has been approved for a clinical trial in humans for the treatment of familial hypercholesterolemia, it was temporarily halted due to safety issues. 145
Unraveling the safety pattern and increasing the accuracy of the strategies are critical for future developments. 146 In theory, the rate of indel/bystander increases with a higher generation of PE, as the edition rate is boosted. Even though the highly scored and topologically well-constructed pegRNA/sgRNA should reduce off-targets, the unwanted events should always be traced from cell models to in vivo, through thorough DNA, exon, and RNA next-generation sequencing (NGS) sequencing. Currently, several platforms, such as GUIDE-seq and VIVO are used as key methods of analysis in vitro and in vivo. A combination of different analyses using more sequencing results will be beneficial for deep unraveling the safety pattern. 147 Furthermore, the application of high-fidelity Cas9, such as Hifi-Cas9, HypaCas9, and miCas9, has been shown to significantly reduce the bystander effects. 148
The application of AAVs and lentiviral vectors should be approached with more caution for PEs due to integration risk and prolonged expression that expose the genome to hazardous off-target effects and unpredicted results. 97 The LNP carrier was recently used in a clinical trial of genome editing; however, nonliver targeting is problematic, as with other vectors. 145 The short half-life of LNP and VLP-based vectors is an advantage due to reducing off-targets. Also, the restriction of Cas9 to specific tissues could be possible by siRNA blocking effect. 145
The efficient and regulated delivery of PE cargo also crucially needs to be considered. Designing a nonviral or viral-based all-in-one construct expressing/carrying the whole PE package could be an optimal construct. It seems easier to have the nCas9/RT/pegRNA/sgRNA in RNPs enclosed by viral vectors, LNPs, or VLPs. 121 However, AAV dual-vector delivery is the only in vivo method practically used and even going to be applied in large animals soon. 72 More importantly, recent versions of PEs are not manageable to drive the expression under specific circumstances. They could be designed for the optimized expression of several elements together in a defined ratio and under the drive of tissue-specific promoters and other cis-regulatory sequences. The development of timing elements to regulate the duration of PE machinery activity and expression is also critical for enhancing safety and efficiency, and preventing unintended consequences. 146 In dual systems, the assembly of proteins and RNAs together needs a more sophisticated design. Moreover, vector targeting to cell receptors would also help in better transduction of cells for a specific purpose.
There are some other applications for gene editing tools in health sciences besides creating animal models and curing the pathogenic genetic background. Genome editing is a way of manipulating critical proteins to appear on the protein domains. Lebek et al. showed that the BE strategy can remove oxidation-sensitive methionine residues on regulatory CaMKII kinase. 149 For example, due to the similar functioning of CaMKII in ventricular arrhythmogenesis and atrial fibrillation, PE can ablate related proteins contributing to other malfunctional factors 150 or even to reshape different pathways in cells. The immunogenicity of Cas9-RT fusion and pegRNA is another safety challenge for clinical use, due to pre-existing immunity or following immune induction. Humoral immunity to Cas9 might lead to the fast recognition of the transduced cells and their elimination. 151 However, targeted delivery to keep delivered cargo out of immune cells during the postnatal stage might prevent triggering of immunity. 150 Masking dominant epitopes in the editor proteins through protein engineering, usage of ortholog proteins, or codon-optimization of nCas9-RT is also effective. 152 In addition, transient expression of editor machines by VLPs or LNPs could be impeding for the burst of both arms of immune responses.
The species-related activity of different generations of PEs has been mentioned in a study on mice and rabbit embryos. The result of the study demonstrated that for both mouse and rabbit, ePE3Max is performing the edition in different loci with high success, but not ePE5Max that was only efficient in the mouse. 73 Based on this, the studies on small animals might not be translatable to larger animals, and the selection of methods should be considered with caution.
Predicting the efficiency and safety of PE methods is a critical demand for the future. Besides many unknown factors related to transcriptomics and genomics, local chromatin structure, epigenetic status, and the architecture of adjacent sequences of the target site could affect the accessibility of the loci and editing efficiency. Phases of the cell cycle and type of cell might impact the rate of on-target efficacy 153 The unraveling of the correlation between host factors, cell milieu, and the editor machine in cells permits better management of PE under any cellular circumstance. Considering that successful PE depends on multiple unknown host factors mediating DNA repair, cell cycle checkpoints, replication, and innate immunity, more understanding of the host response to the process should make it more efficient. 154
Even though the rate of bystander error of PEs is very low, continuous expression of Cas9 and RT in the cell is a big concern, particularly for pluripotent cells. For stopping the Cas9 expression, the use of inhibitors of nuclear export, such as SINEs, might offer one solution to increase the PE safety. 146
PE technology is on its optimization route to reach a clinically efficient level. The recent large fragment PE-based integration strategies showed insufficient efficiency. In the PASSIGE version, even after the efficient establishment of the recombinase landing site by dual-flap PE editing, fragment integration by Bxb1 recombinase was detected in a range of 2.6–6.8%. Therefore, newer versions of this recombinase, evoBxb1 or eeBxb1, which were developed through phage assisted evolution, showed several times higher integration rates compared to the original Bxb1 enzyme (155). In addition, due to the importance of the genome context on the fate of the PE method, machine learning predictive models such as ePRIDICT have been developed to quantify the local chromatin context’s impact on PE editing rates (156). These kinds of improvements bring more hope on the road to editing by the PE strategy.
Finally, regarding the unpredictable outcome of PE in different contexts, personalized PE might be another aspect that becomes necessary to adjust the process individually in a context-dependent manner.
Footnotes
ACKNOWLEDGMENT
The study was supported by the
AUTHORS’ CONTRIBUTIONS
S.Ylä-Herttuala, P.Mäkinen, and S.Younes Hosseini devised the topic and the principal conceptual ideas. S.Younes Hosseini and R.Mallick mainly wrote the manuscript. S.Younes Hosseini and R.Mallick contributed to the design of the figure. All authors discussed and commented on the manuscript.
AUTHOR DISCLOSURE
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
FUNDING INFORMATION
The study was supported by the ERC Advanced grant (884382), Flagship program of the Finnish Medical Research Council (337120) and Horizon EU grant, Geremy (101080204).