
Abstract
Limb-girdle muscular dystrophy type 2D/R3 (LGMD2D/R3) is a progressive muscular dystrophy that manifests with muscle weakness, respiratory abnormalities, and in rare cases cardiomyopathy. LGMD2D/R3 is caused by mutations in the SGCA gene resulting in loss of protein and concomitant loss of some or all components of the dystrophin-associated glycoprotein complex. The sgca-null (sgca−/−
) mouse recapitulates the clinical phenotype of patients with LGMD2D/R3, including dystrophic features such as muscle necrosis and fibrosis, elevated serum creatine kinase (CK), and reduction in the generation of absolute muscle force and locomotor activity. Thus, sgca−/−
mice provide a relevant model to test the safety and efficacy of gene transfer. We designed a self-complementary AAVrh74 vector containing a codon-optimized full-length human SGCA (hSGCA) transgene driven by a muscle-specific promoter, shortened muscle creatine kinase (tMCK). In this report, we test the efficacy and safety of
Introduction
Limb-girdle muscular dystrophy (LGMD) is a heterogeneous group of genetic forms of muscular dystrophy with many subtypes that are each caused by a unique mutation and compilation of symptoms. 1 The sarcoglycanopathies are included as a subset of LGMDs and comprise four major subtypes, namely LGMD2C/R5, -D/R3, -E/R4, and -F/R6, of which each subtype is caused by mutations in the gene that codes for a sarcoglycan protein subunit within the sarcoglycan complex (i.e., γ-sarcoglycan, α-sarcoglycan, β-sarcoglycan, and δ-sarcoglycan, respectively). 1,2 Sarcoglycans play a vital role in preventing muscle damage during muscle contraction and all four sarcoglycans must be present to form a functional sarcoglycan complex. 1,2 The analogous features of the LGMDs are the weakening and atrophy of proximal muscles, specifically in the limb-girdle region. 1,2 However, even within each of the LGMD subtypes, there is considerable variability in the age of onset, severity, prognosis, and disease progression. 1
LGMD subtype 2D/R3 (LGMD2D/R3), often referred to as α-sarcoglycanopathy, is caused by mutations in the SGCA gene, which leads to complete or reduced loss of functional protein with impairment of other structural components of the dystrophin-associated protein complex. 3,4 Notably, loss of the α-sarcoglycan (α-SG) protein leads to a progressive muscular dystrophy with deteriorating muscle function, with onset from 1 to 15 years of age. 5 Symptoms include the following: delayed ambulation, weakness in proximal muscles caused by fat replacement and fibrosis, elevated creatine kinase (CK), scoliosis, and joint contractures. 1,6 The debilitating disease often leads to wheelchair dependency and death due to respiratory failure. 4,6 To date, there are no disease-modifying therapies for LGMD2D/R3. 5,6
The sgca−/− murine knockout model of LGMD2D/R3 lacks α-SG protein and displays a dystrophic phenotype similar to the human disease due to its ongoing and progressive muscle necrosis that occurs with age. 7 Specifically, the sgca−/− mice display other molecular and chemical characteristics, which include elevated CK, centralized nuclei (CN), increased necrosis, fibrosis, and inflammation. 7 In addition, the mice recapitulate the human disease with their presentation of functional deficits in muscle force generation and ambulation, thus making the sgca−/− mice an ideal model to study and assess the effects of disease-modifying therapies. 7
Gene transfer therapy is a promising modality given the monogenetic nature of the disease. 5,6 We previously conducted a preclinical comparison study delivering the adeno-associated virus (AAV) vector carrying the full-length hSGCA cDNA driven by a muscle-specific promoter (shortened muscle creatine kinase, tMCK). 8 This was packaged into an AAV1 and delivered intramuscularly into the tibialis anterior (TA) compartment of sgca−/− mice. The results from this previous comparison study confirmed that the tMCK promoter was more efficient in transducing muscle fibers than MCK, cytomegalovirus, or desmin promoters. 8
To further increase transduction efficiency in muscle fibers, we have packaged the same full-length SGCA, driven by tMCK, into an AAVrh74 vector. The AAVrh74 vector has been used previously in various studies to deliver transgenes successfully, including GALGT2,
9
dysferlin,
10
β-sarcoglycan,
11,12
and microdystrophin.
13
More recently, local intravascular delivery of this construct (
Materials and Methods
Animal models
All procedures were approved by The Research Institute at the Nationwide Children's Hospital Institutional Animal Care and Use Committee. C57BL/6-TgN homozygous mice were provided as gift from Kevin Campbell. 7 Knockout (sgca−/− ) mice were bred and maintained as homozygous animals under standardized conditions in the Animal Resources Core at the Research Institute at Nationwide Children's Hospital (RINCH). Wild-type (WT) C57/BL6 mice were from the breeding colony at RINCH and purchased from The Jackson Laboratory. Mice were maintained on Teklad Global Rodent Diet (3.8% fiber, 18.8% protein, 5% fat chow) with a 12:12-h dark:light cycle. All animals were housed in standard mouse cages with food and water ad libitum. For all experiments, mice from both sexes were used.
Genotyping
DNA genotyping was used to identify sgca−/− mice. DNA from tail clippings was isolated and analyzed by polymerase chain reaction (PCR) using OneTaq DNA Polymerase (New England Biolabs, Ipswich, MA). A series of primers was used in the PCR analysis to determine the SGCA knockout status. The following primers and conditions were used: Intron1 (CAGGGCTGGGAGCTGGGTTCTG); mutant primer-intron 3 (CCCAGGGCCTTGATGCCT); and NEOTR (GCTATCAGGACATAGCGTTGGCTA). Reactions were carried out on genomic DNA for 30 cycles under the following conditions: 94°C, 5 min; 94°C, 1 min; 64°C, 1 min; 72°C, 2.5 min; and 72°C, 7 min.
hSGCA gene construction
The
Vector production
AAVrh74.tMCK.hSGCA was packaged into the AAV serotype rh74 capsid using the standard triple transfection protocol as previously described. 16 –18 A quantitative PCR (qPCR)-based titration method was used to determine an encapsulated vector genome titer utilizing a Prism 7500 Fast TaqMan detector system (PE Applied Biosystems). 19
Gene delivery
Systemic delivery in mice
Systemic delivery was administered via injection of vector into the tail vein of sgca−/−
mice. Mice were randomly assigned and injected with 1.0 × 1012 vg (n = 6, 3 male [M]/3 female [F]), 3.0 × 1012 vg (n = 6, 1M/5F), or 6.0 × 1012 vg (n = 6, 3M/3F) total dose (5.0 × 1013, 1.0 × 1014, and 2.0 × 1014 vg/kg, respectively) of
Laser monitoring of open-field cage activity
An open-field activity chamber was used to determine the overall activity of the experimental mice. Mice at 16–17 weeks of age from the WT C57BL/6 (n = 6, 6M) and sgca−/−
LR (n = 6, 3M/3F) control groups, along with
Mouse activity was monitored using the Photobeam activity system (San Diego Instruments, San Diego, CA). This system uses a grid of invisible infrared light beams that traverse the animal chamber front to back and left to right to monitor the position and movement of the mouse within an x-y-z plane. Activity was recorded for 1-h cycles at 5-min intervals. Mice were acclimatized to the activity test room for an initial 1-h session 3 and 4 days before data acquisition began. Mice were tested in individual chambers. The testing equipment was cleaned between each use to reduce mouse reactionary behavioral variables that could alter results. The data were converted to a Microsoft Excel worksheet, and all calculations were done within the Excel program. Individual beam breaks for movement in the x and y planes were added up for each mouse to represent total ambulation, and beam breaks in the z plane were added up to obtain vertical activity within the 1-h time interval.
TA tetanic contraction for functional assessment
The TA assessment procedure followed the protocol listed in Hakim et al. 23 Mice were anesthetized with ketamine/xylazine mixture (137.5 and 10 mg/kg, respectively) administered intraperitoneally. The hind limb skin was removed to expose the TA muscle and patella. The length of muscle is measured after dissection, before placement of mouse, and the length is entered into the software. Care was taken to limit drying of the exposed muscle by constantly dampening the exposed muscles with a saline-dampened Kimwipe drape. The TA distal tendon was then dissected out (left and right side per animal; average of both legs used for analysis [n = 12 per cohort]), and a double square knot was tied around the tendon with 4-0 suture as close to the muscle as possible and then the tendon was cut.
Mice were then transferred to a thermal controlled platform and maintained at 37°C. To stabilize the leg, a metal pin was placed behind the patellar tendon and the knee was secured to the platform with the distal TA tendon sutured to the level arm of the force transducer (Aurora Scientific, Aurora, Canada). An electrode was placed near the sciatic nerve to stimulate it. A warm-up protocol designed by Aurora Scientific was initiated where the resting tension was set at 3–4 g force and maintained for 5 min, muscle stimulation at 1 Hz (three times–30 s apart), and an additional muscle stimulation at 150 Hz (three times–60 s apart).
Once the muscle was stabilized, the resting tension was set to a length (optimal length) where twitch contractions were maximal. After a 3-min rest period, the TA was stimulated at 50, 100, 150, and 200 Hz, allowing a 1-min rest between each stimulus. Following a 5-min rest, the muscles were then subjected to a series of 10 isometric contractions, occurring at 1-min intervals with a 10% stretch relengthening procedure. The duration of tetanic contraction lasts 200 ms. After the eccentric contractions, the mice were euthanized and both TA muscles were dissected and frozen for histology and molecular studies. Maximum absolute force is determined from the plateau of the frequency/force relationship; force generated at 150 Hz frequency was used for the purposes of data analysis.
Formulas: Absolute Force = Force at 150 Hz*9.8 (where 9.8 mN = 1 g) TA-Specific Force = Absolute Force/Cross-Sectional Area Cross-Sectional Area = muscle weight (g)/[TA fiber length (cm) × 1.06 (g/cm3)]
Diaphragm tetanic contraction for functional assessment
Mice were euthanized and the diaphragm (DIA) was dissected with rib attachments and central tendon intact and placed in Kreb's–Henseleit (K-H) buffer (118 mM NaCl, 4.7 mM KCl, 1.2 mM MgSO4, 1.25 mM CaCl2, 1.2 mM KH2PO4, 25 mM NaHCO3, 11 mM glucose) as previously described. 20,24,25 A 2–4-mm-wide section of DIA was isolated per animal per cohort (n = 6). DIA strips were tied firmly with braided surgical silk (6/0; Surgical Specialties, Reading, PA) at the central tendon, and sutured through a portion of rib bone affixed to the distal end of the strip. Each muscle was transferred to a water bath filled with oxygenated K-H solution that was maintained at 37°C. The muscles were aligned horizontally and tied directly between a fixed pin and a dual-mode force transducer-servomotor (305C; Aurora Scientific). Two platinum plate electrodes were positioned in the organ bath so as to flank the length of the muscle. The muscle was stretched to optimal length for measurement of twitch contractions, and then allowed to rest for 10 min before initiation of the tetanic protocol. Once the muscle was stabilized, it was set to an optimal tension of 1 g and subjected to a warm-up, which consisted of three 1-Hz twitches every 30 s followed by three 150-Hz twitches every minute. After a 3-min rest period, the DIA was stimulated at 20, 50, 80, 120, 150, and 180 Hz, allowing a 2-min rest period between each stimulus, each with a duration of 250 ms to determine maximum tetanic force. Muscle length and weight were measured. The force was normalized for muscle weight and length. Maximum absolute force is determined from the plateau of the frequency/force relationship; force generated at 150 Hz frequency was used for the purposes of data analysis.
Formulas:
Absolute Force = Force at 150 Hz*9.8 (where 9.8 mN = 1 g)
DIA-Specific Force = Absolute Force/Cross-Sectional Area
Cross-Sectional Area = muscle weight (g)/[DIA fiber length (cm) × 1.06 (g/cm3)]
Tissue processing
Skeletal muscles were extracted from each mouse, placed on a saline-dampened gauze, then placed on 7% gum tragacanth mounted wooden chucks and fresh frozen in cooled methyl butane. Organs were bisected and half were placed in 10% neutral buffered formalin followed by paraffin embedding for sectioning and hematoxylin and eosin (H&E) staining. The other half of the organ was fresh frozen for subsequent molecular studies.
Immunofluorescence
Cryosections (12-μm thick) from the TA, gastrocnemius (GAS), quadriceps (QD), psoas major (PSO), gluteus (GLUT), triceps (TRI), DIA, and heart (HRT) muscles were subjected to immunofluorescence staining for the hSGCA transgene via our previously used protocol. 11 Sections were incubated with a rabbit monoclonal α-SG primary antibody (Catalog No. ab189254; Abcam, Cambridge, United Kingdom) at a dilution of 1:100. Four random 20 × images covering the four different quadrants of the muscle section were taken using a Zeiss (Germany) AxioCam MRC5 camera. The percentage of fibers positive for α-SG staining compared with controls was determined for each image and averaged for each muscle. Positive α-SG fiber expression was defined as having at least 50% of the fiber staining brighter than the vehicle-treated sgca−/− LR controls, as previously described. 11 The test article was blinded at the time of injection. The operator who conducted the injections did not perform any analysis outside of the injection. There is no expression or residual protein in the untreated group, and so, it is clear which animal received treatment and which did not when observing under the scope, leaving blinding irrelevant for immunofluorescence quantification. To mitigate variability in intensity, images were taken at the same exposure.
Fibers counted were defined by the structural appearance of the fiber's cross section. To facilitate scoring, National Institutes of Health (NIH) ImageJ with the Cell Counter plugin was used to count total fibers. By convention, a “Type #” and color were selected to score/mark positive fibers. The Cell Counter tracked the counts as fibers were selected. Positive fibers were scored based on the original image exposure; there was no adjustment to the brightness or contrast of any image during the positive image scoring process. Once positive fiber selections were completed, the brightness and contrast of the image were adjusted to facilitate visualization of the negative fibers. No additional positive fibers were scored once the brightness and contrast were adjusted. The remaining fibers were scored as negative using a different “Type #” and color for the score/mark. Once scoring was complete, “Export Image” was selected from the Cell Count workspace. This image was saved separately as an annotated .jpeg image. Total fiber counts were determined after all fibers were counted. Quantification data of immunofluorescent-positive fibers expressing α-SG protein are reported as mean ± standard of error of the mean, with six mice per treatment group.
Western blot analysis
Western blots were performed according to our previously used protocol, with several modifications specific for each antibody used. 11 Samples from WT C57BL/6 mice, sgca−/− LR mice, and vector-dosed sgca−/− mice were used for each Western blot. A 1:10,000 dilution of a rabbit monoclonal α-SG antibody (Catalog No. ab189254; Abcam) was used to detect α-SG in all blots. For the protein loading control, a 1:5,000 dilution of a mouse monoclonal α-actinin antibody (Catalog No. A7811; Sigma-Aldrich) and a 1:1,000 dilution of a rabbit monoclonal mouse vinculin antibody (Catalog No. 70062; Invitrogen) were used to detect α-actinin and vinculin in muscle-tissue and nonmuscle-tissue blots, respectively. Anti-mouse (Catalog No. AP308P; Millipore) and anti-rabbit (Catalog No. 656120; Life Technologies) secondary horseradish peroxidase antibodies were used for enhanced chemiluminescence immunodetection. For muscle-tissue blots, densitometry quantification was determined using ImageQuant TL analysis software, which quantified the raw densitometric units for each protein band at the appropriate size.
Morphometric analysis
H&E staining was performed on 12-μm-thick cryosections of muscle from 16- to 17-week-old WT C57BL/6 mice (n = 6), sgca−/−
LR mice (n = 6), and
Picrosirius red stain and collagen quantification
Picrosirius red staining was performed to determine the levels of collagen deposition in muscle tissue using a previously published protocol from our laboratory.
12
Staining was performed on 12-μm cryosections from 16- to 17-week-old WT C57BL/6 (n = 6), sgca−/−
LR (n = 6), and
Biodistribution qPCR analysis
TaqMan qPCR was performed to quantify the number of vector genome copies as previously described.
11,26,27
A vector-specific primer probe set was used to amplify a sequence of the intronic region directly downstream from the tMCK promoter that is unique and located within the
Hematology
Whole blood was retrieved from cardiac puncture for blood chemistries from treated WT and knockout mice. Blood was collected in a serum separating tube and centrifuged for 10 min at 15,000 rpm. Serum was collected, frozen, and sent to Charles River Laboratories for processing and assessment of serum chemistry. Due to limited sample volume, liver enzymes and glucose chemistries were prioritized.
Serum CK measurement
Levels of CK were measured in the sera of WT C57BL/6 mice (n = 6), sgca−/−
LR mice (n = 6), and
Histopathology
At necropsy, muscles were fresh frozen in liquid nitrogen-cooled methyl butane, and tissues were stained with H&E. All other organs were harvested and fixed in formalin and embedded in paraffin. Slides and all tissues were sent to GEMpath, Inc., for formal review by a veterinary pathologist.
Statistical analysis
Data were analyzed using a one-way ANOVA with multiple comparisons between groups assessed by Tukey's or Dunnett's post hoc analysis test using GraphPad Prism 5 (GraphPad Software, La Jolla, CA), unless otherwise specified.
Results
Dose-escalation study of scAAVrh74.tMCK.hSGCA systemic gene transfer to sgca−/−
mice
To determine the safest and most efficacious dose, we investigated the delivery of three separate doses of vector in a dose-escalation study, where the lateral tail vein of 4-week-old sgca−/− mice were treated with 1.0 × 1012 vg total dose (5.0 × 1013 vg/kg), 3.0 × 1012 vg total dose (1.0 × 1014 vg/kg), or 6.0 × 1012 vg total dose (2.0 × 1014 vg/kg) of scAAVrh74.tMCK.hSGCA, corresponding to low, intermediate, and high doses. Mice were euthanized 12 weeks postgene transfer to assess hSGCA transgene expression in the TA, GAS, QD, GLUT, PSO, TRI, DIA, and HRT muscles using immunofluorescence. Robust expression of hSGCA was observed at the sarcolemma across skeletal tissues (Fig. 1A).
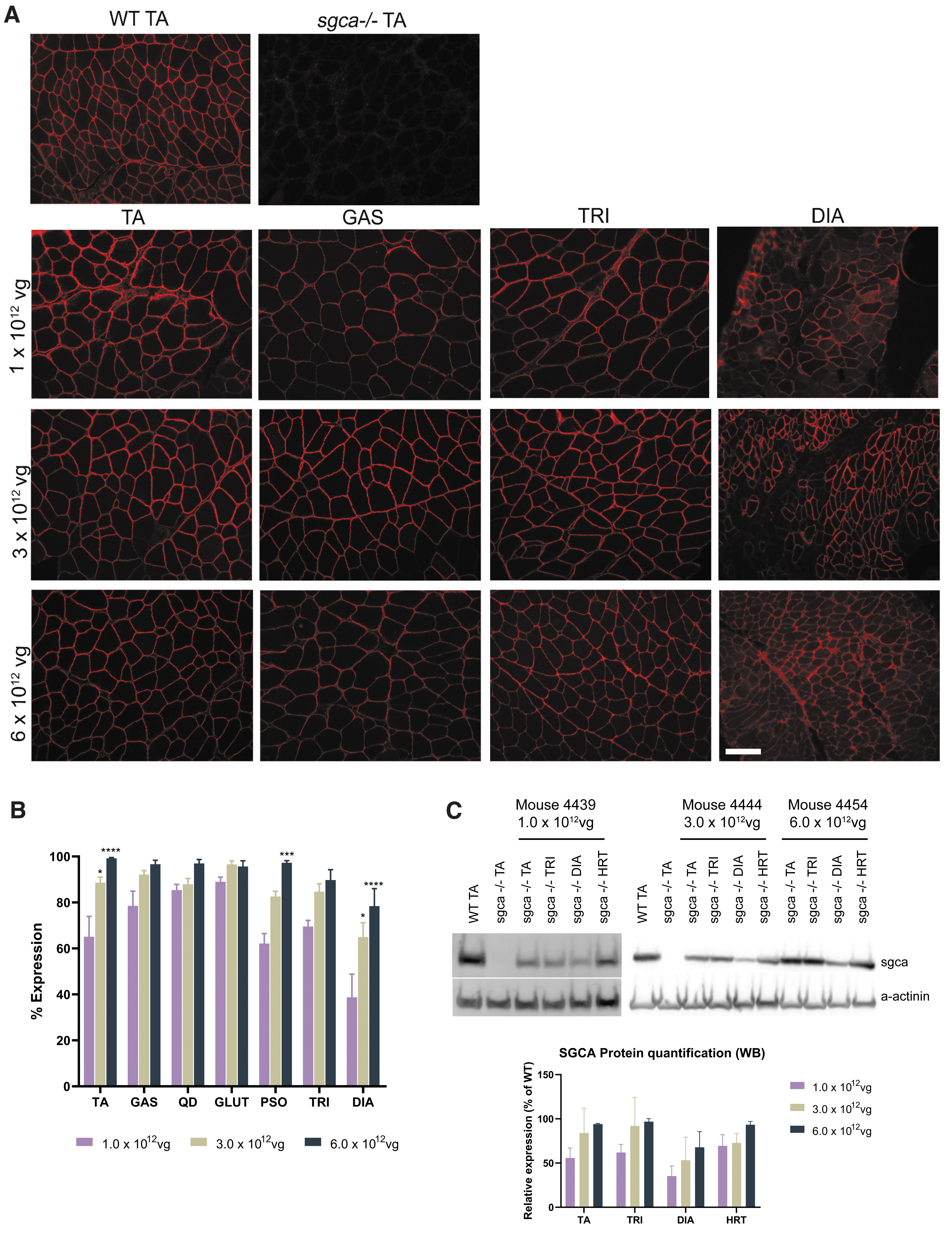
Systemic treatment with
Quantitative analysis of positive fiber expression demonstrated a dose-dependent increase in expression in each muscle examined (Fig. 1B). Mean hSGCA expression assessed by immunohistochemistry in mice treated with the lowest tested dose of 1.0 × 1012 vg total dose was 70.07% ± 3.71% overall expression of positive fibers in the skeletal muscles. Mean hSGCA overall expression in mice treated with the intermediate dose of 3.0 × 1012 vg total dose was 85.35% ± 2.36% in all skeletal muscles. Mean hSGCA expression in mice treated with the highest dose of 6.0 × 1012 vg total dose was 93.86% ± 2.02% in all skeletal muscles. Moreover, Western blots shown in Fig. 1C confirm protein expression across muscle tissues in treated mice at the lowest dose tested; the hSGCA expression in the HRT muscle remained at least 69% of WT for the low, middle, and high doses.
Histopathological characteristics of both humans and mice devoid of α-SG protein include CN, irregularities in fiber size distribution, necrosis, and fibrosis.
1,7
H&E staining was used to visualize muscle morphology, including fiber size and CN (Fig. 2). As shown in Fig. 2A and B, a normalization of fiber size distribution, similar to that observed in WT controls, was observed in the TA, QD, TRI, GLUT, and PSO of
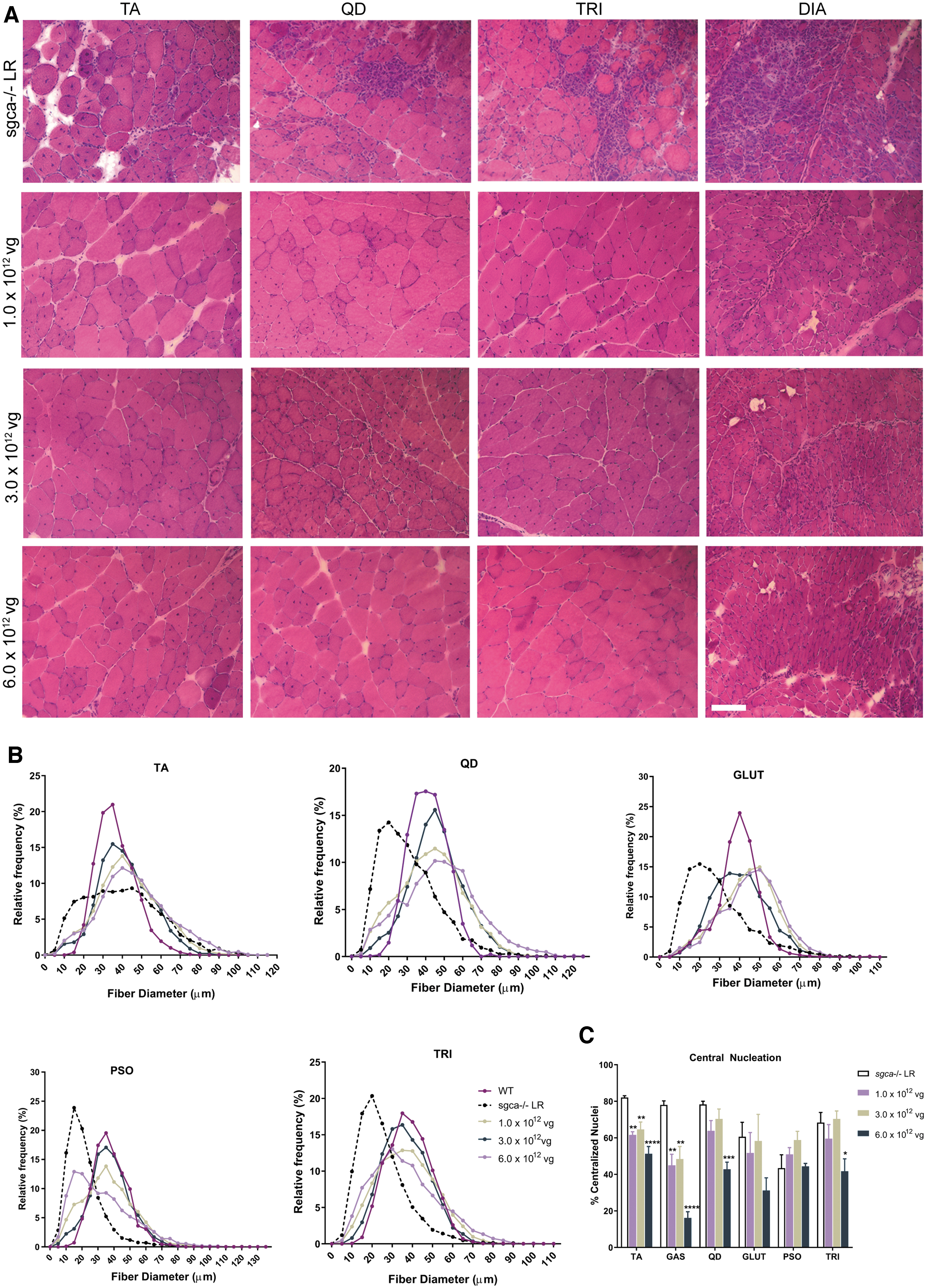
Improvement in muscle morphology by
In sgca−/−
mice treated with scAAVrh74.tMCK.hSGCA, reduction in CN was also observed. The effects of
Fibrosis, where the tissue is overcome by collagen, often occurs in the muscles of patients with LGMD, leading to the formation of scar tissue. As the GLUT and PSO muscles are critical for ambulation, reduction in fibrotic content is particularly important to evaluate in these muscles. Fibrosis was assessed using a picrosirius red stain to detect collagen I and III content, as a marker of fibrosis. As shown in Fig. 3A, a robust reduction in red staining was observed in sgca−/−
mice after treatment with
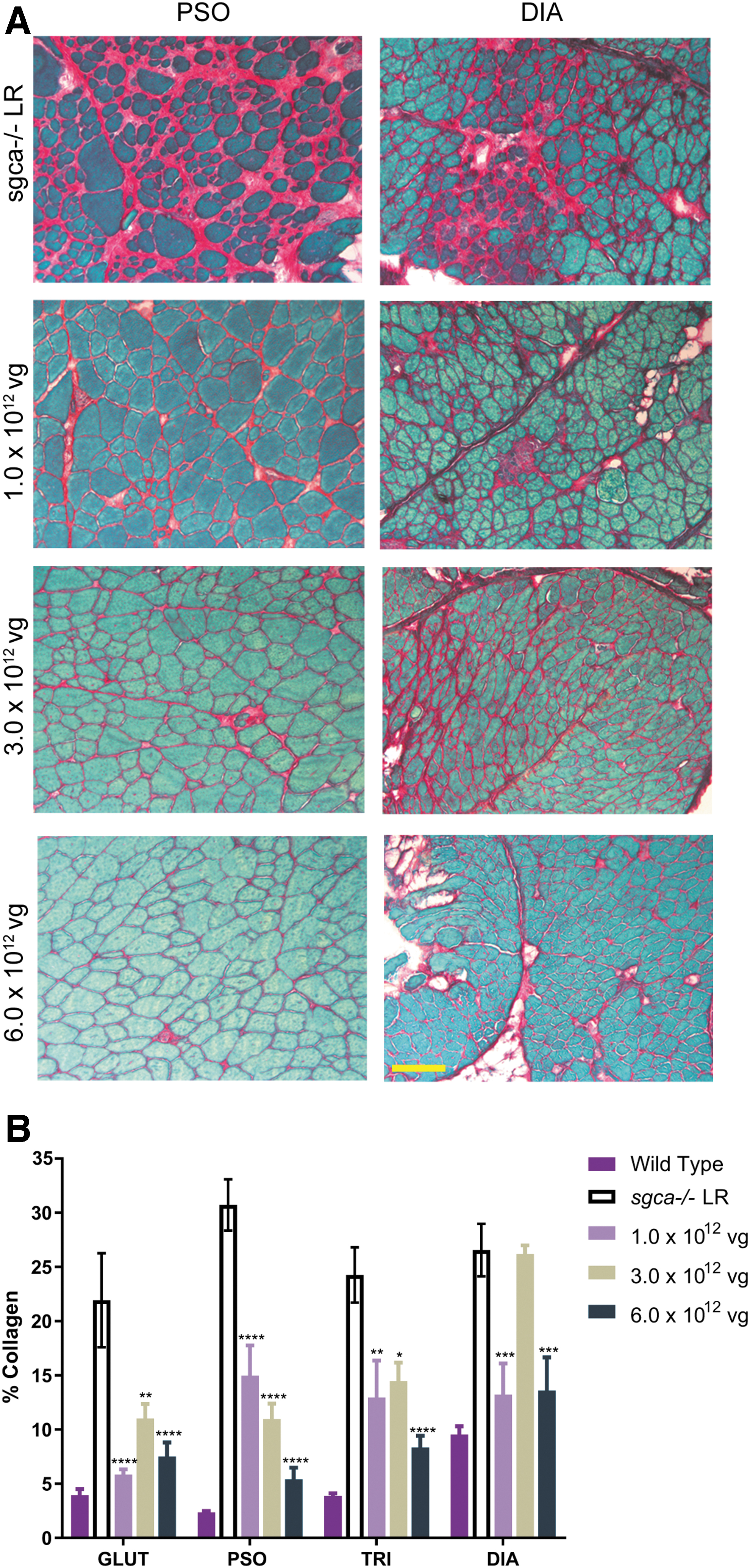
Reduction in fibrosis in sgca−/−
mice treated with
scAAVrh74.tMCK.hSGCA improves diaphragm and TA muscle function and increases locomotor ability
As weakness and loss of function of proximal muscles are major symptoms of LGMD2D/R3, and respiratory failure is the leading cause of death in LGMD2D/R3, 5,28 improving the functionality and strength of the TA and DIA are imperative to increasing the length and quality of life in patients with LGMD2D/R3. As an experimental tool, strips of the DIA and whole TA muscles were used to confirm the correlation between hSGCA expression and muscle strength. As shown in Fig. 4A and B, a deficit in specific force and resistance to contraction-induced injury was identified in the TA and specific force in the DIA muscles of sgca−/− LR mice compared with WT mice.
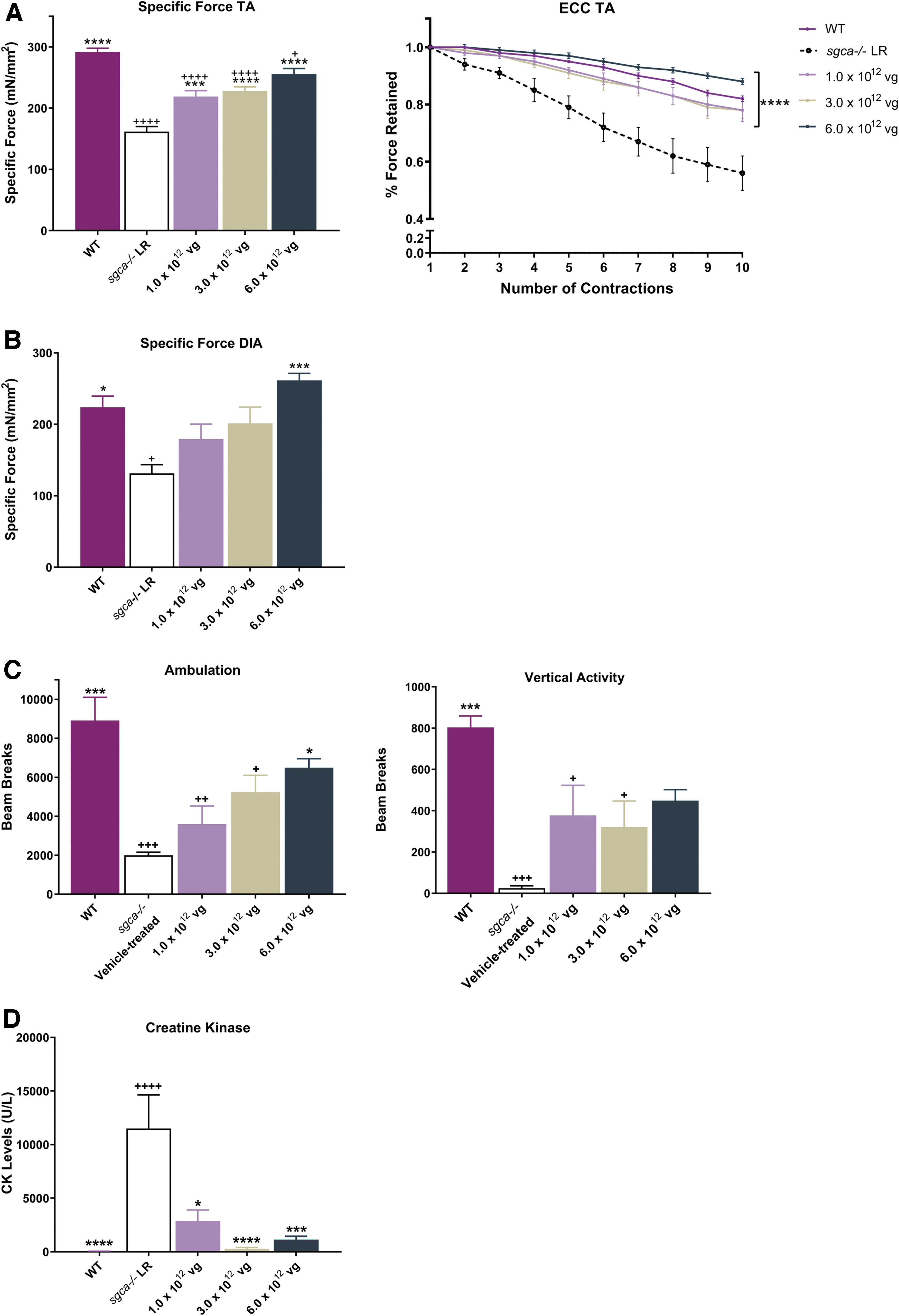
Functional benefits to skeletal muscle after treatment with
TA muscles of sgca−/−
mice exhibited a significant functional deficit of 44% in the reduction of specific force output compared with WT mice (161.6 ± 8.20 mN/mm2 vs. 291.7 ± 6.17 mN/mm2, respectively; p < 0.0001), as well as a greater loss of force from that produced following a rigorous eccentric contraction protocol (44.0% ± 6.0% loss in sgca−/−
LR mice; 18.0% ± 1.0% loss in WT mice; p < 0.0001) (Fig. 4A). Twelve weeks following tail vein delivery, we noted a significant improvement after treatment with low, intermediate, and high doses of
In the DIA of sgca−/−
LR mice, the specific force generated showed a 41% reduction in strength compared with WT mice (131.5 ± 12.07 mN/mm2 vs. 223.8 ± 15.85 mN/mm2). A significant improvement in force was observed following treatment with
Additional symptoms of LGMD2D/R3 include exercise intolerance and reduced activity and ambulation, possibly due to muscle damage, resulting in muscle loss and fatigue. 4 –6 To assess the level of physical activity, sgca−/− LR and WT mice were subjected to an open-field activity protocol similar to that used in previous reports. 20,21 The ambulation-related activities of mice were monitored to determine if the lack of SGCA in the sgca−/− mouse leads to a decrease in ambulation compared with WT mice. The graphs in Fig. 4C depict a reduction in ambulation and vertical rearing in the sgca−/− mouse model compared with WT controls. The mean horizontal ambulatory beam breaks recorded in the sgca−/− LR mice were 2,000 ± 159 beam breaks per hour, a 77.5% decrease in ambulation compared with 8,911 ± 1,193 beam breaks per hour in WT controls. The mean vertical rearing beam breaks recorded in the sgca−/− mice were 24.75 ± 11.47 beam breaks per hour, a 97% decrease in vertical rearing compared with 803.3 ± 55.03 beam breaks per hour in WT mice.
After treatment with scAAVrh74.tMCK.hSGCA, the ambulation and vertical rearing activities of mice increased 12 weeks postgene transfer. The mean horizontal ambulation increased to 3,595 ± 55.03 beam breaks per hour in mice treated with 1.0 × 1012 vg total dose, 5,238 ± 861.9 beam breaks per hour in mice treated with 3.0 × 1012 vg total dose, and 6,487 ± 467.9 beam breaks per hour in mice treated with 6.0 × 1012 vg total dose. The mean vertical rearing activity increased to 377 ± 146.1 beam breaks per hour in mice treated with 1.0 × 1012 vg total dose, 321 ± 126.1 beam breaks per hour in mice treated with 3.0 × 1012 vg total dose, and 448.8 ± 53.43 beam breaks per hour in mice treated with 6.0 × 1012 vg total dose (Fig. 4C). The physical activities of the vector-treated mice showed improvement from 44% to 69% in ambulation and 92% to 94% in vertical rearing compared with sgca−/− LR mice. In addition, serum CK levels were significantly reduced in all treated groups compared with untreated mice (Fig. 4D). Together, these data show that the delivery of hSGCA restores the physical activity and protects against the breakdown of muscle in sgca−/− mice.
Safety and biodistribution analysis of scAAVrh74.tMCK.hSGCA
As a safety provision, blood chemistries and hematology studies were performed on vector-dosed sgca−/−
and WT mice. All values were within the normal reference ranges for mice (Fig. 5). Furthermore, tissue sections of all organs stained with H&E from
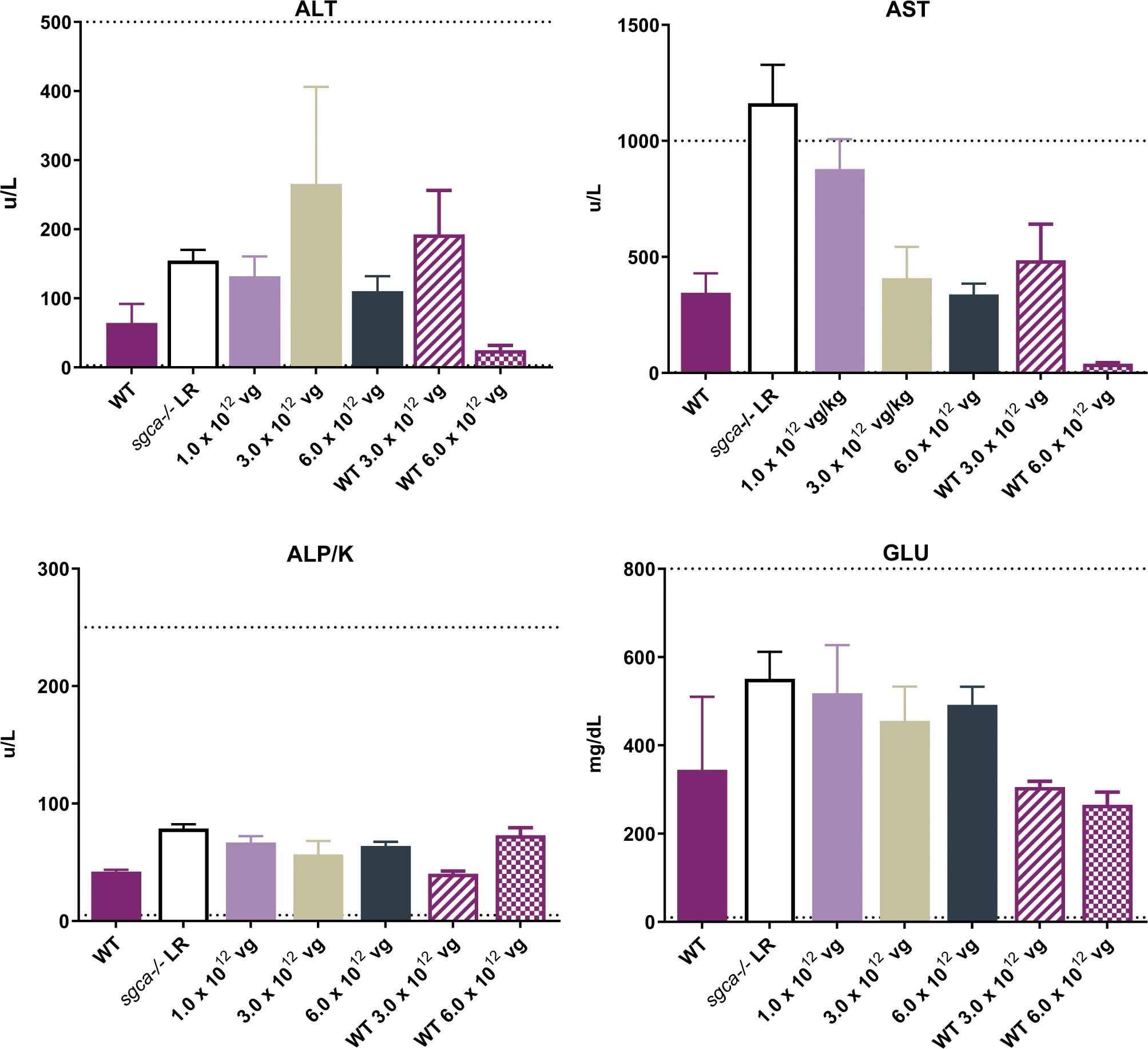
No evidence of toxicity through blood chemistries after treatment with
Vector biodistribution qPCR was performed to quantify vector genome presence following delivery of
Discussion
LGMD2D/R3 is caused by mutations in the SGCA gene, which leads to complete or reduced loss of functional α-SG protein and concomitant loss of other structural components of the dystrophin-associated protein complex. 3,4,6 The loss of the α-SG protein leads to a progressive muscular dystrophy with deteriorating muscle function and respiratory abnormalities. 4,6 To date, there are no disease-modifying therapies for LGMD2D/R3. 5 Given the monogenetic nature of the disease, gene transfer therapy has emerged as a promising treatment modality for patients with LGMD2D/R3.
In our recent Phase I/II LGMD2D/R3 intravascular gene transfer (via isolated limb infusion [ILI]) trial, we demonstrated improvement in force in target muscles; however, there was no improvement in walk time due to the inability to target hip girdle muscles.
14
Thus, the successful systemic (intravenous) gene transfer of
Although cardiac defects are sporadically observed in patients with LGMD2D/R3, having additional cardiac protection after treatment can be beneficial. Prior studies indicate that the sgca−/−
mice do not exhibit gross morphological changes in heart at up to 8 months of age.
29
However, we hypothesize that the increase in ambulatory motor activity as a result of
In previous clinical studies, we have shown that hSGCA packaged into the AAV1 capsid with expression driven by muscle-specific promoters resulted in transgene expression in skeletal muscle tissue after local delivery. 8,18,30 In this study, we packaged hSGCA into rAAVrh74 to maximize delivery to target tissue (biodistribution). rAAVrh74 is a serotype isolated from lymph nodes of rhesus monkeys with high homology (93%) to the AAV8 capsid that displays high affinity for skeletal and cardiac muscle tissue. 9,15 The data in this study further support the ability of rAAVrh74 to achieve targeted delivery to various skeletal muscle tissue. In addition, in the organs we tested, we demonstrated limited off-target effects that could be attributed to use of the muscle-specific promoter, tMCK, in driving hSGCA transgene expression in the appropriate tissues.
To improve efficiency of α-SG protein expression, we packaged hSGCA into a self-complementary vector. Self-complementary AAV vectors contain an inverted repeat genome that promotes the formation of dsDNA, thus allowing replication and transcription to occur without the need for multiple vector genomes to promote these processes. 31,32 As such, use of self-complementary vectors eliminates the rate-limiting step to allow more rapid expression of the transgene. Indeed, the results from this study demonstrate expression 12 weeks after systemic administration at all doses tested.
Of interest, we recently have shown that intravascular gene transfer of
Supportive functional data in this study further demonstrate that the intravenous gene transfer of
In addition, treatment with
Prominent histopathology, which includes centrally located nuclei, wide variability in fiber size, inflammation, necrosis, and fibrosis, is typically observed through muscle biopsies of patients with LGMD2D/R3. 5 After hSGCA delivery, mice had a reduction in CN, a more even distribution of myofiber size, and a reduction in collagen content, with muscles having an overall healthier appearance compared with vehicle-treated mice. It must be noted that the ability to reduce/reverse CN was muscle- and dose-dependent. We observed that the CN reduction and fibrosis levels did not follow a dose-dependent pattern in the intermediate-dose cohort; however, this could be possibly attributed to baseline differences among the cohorts. Onset of progressive muscular dystrophy in sgca−/− mice occurs as early as 1 week of age. 7 Nonetheless, the total reduction in histopathology and scar tissue was concomitantly associated with improvement in the overall normal function and physiology of the muscles in vector-treated mice.
Finally, safety studies conducted through qPCR, serum chemistry analysis, and histopathology lacked any signs of toxicity. Consistent with an earlier study by Wang et al.,
34
in which expression of luciferase driven by tMCK leakage in the liver was observed but was significantly less than the constitutive cytomegalovirus promoter and other promoters being tested, transgene expression regulated by the tMCK promoter was detected only in targeted tissues (all muscles) and was absent in other (nonmuscle) organs, except for the liver. Histopathology review of all tissues (including liver) by a certified veterinarian pathologist concluded that the systemic delivery of
In sum, this dose-escalation study provides preclinical data to support that the lowest dose systemically tested here—that is, 1.0 × 1012 vg total (5.0 × 1013 vg/kg)—is sufficient to reduce the signs and symptoms associated with loss of α-SG protein. At the lowest dose tested, functional improvement in all muscles, as demonstrated by increase in strength and locomotor behavior (ambulation and rearing), was observed in vector-treated mice. Safety studies show no signs of toxicity, even at the highest delivered dose of 6.0 × 1012 vg total (2.0 × 1014 vg/kg), suggesting the potential ability to increase dose based on progression and the severity of the disease in a clinical setting of patients with LGMD2D/R3.
Footnotes
Authors' Contributions
D.A.G. and L.R.K. designed the experiments. L.R.K. cloned and prepped the
Acknowledgments
We thank Jerry R Mendell, MD, for providing advice on interpretation of findings and the Nationwide Children's Viral Vector Core for Vector Production. We would also like to thank Terri Shaffer, MLAS, RLATG, for performing intravenous tail vein injections. Finally, we would like to thank Dr. Kevin Campbell for his gift of the sgca−/− mice. Medical writing and editorial support were provided by Khampaseuth Thapa, PhD and Lucia Quintana-Gallardo, PhD, of Sarepta Therapeutics, Inc., and Purvi Kobawala Smith, MS, MPH, of Health & Wellness Partners, LLC, Upper Saddle River, NJ, funded by Sarepta Therapeutics, Inc.
Author Disclosure
D.A.G., E.R.P., R.A.P., E.L.P., and L.R.K. are employees of Sarepta Therapeutics, Inc. and may have stock options. K.N.H. has nothing to disclose.
Funding Information
This study was funded by Sarepta Therapeutics, Inc.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.