
Abstract
Urocortin-2 (UCn2) peptide infusion increases cardiac function in patients with heart failure, but chronic peptide infusion is cumbersome, is costly, and provides only short-term benefits. Gene transfer would circumvent these shortcomings. We previously showed that a single intravenous (IV) injection of AAV8.UCn2 increases plasma UCn2 and left ventricular (LV) systolic and diastolic function for at least 7 months in normal mice. Here we test the hypothesis that IV delivery of AAV8.UCn2 increases function of the failing heart. Myocardial infarction (MI, by coronary ligation) was used to induce heart failure, which was assessed by echocardiography 3 weeks after MI. Mice with LV ejection fraction (EF) <25% received IV delivery of AAV8.UCn2 (5×1011 gc) or saline, and 5 weeks later echocardiography showed increased LV EF in mice that received UCn2 gene transfer (p=0.01). In vivo physiological studies showed a 2-fold increase in peak rate of LV pressure development (LV +dP/dt; p<0.0001) and a 1.6-fold increase in peak rate of LV pressure decay (LV −dP/dt; p=0.0007), indicating increased LV systolic and diastolic function in treated mice. UCn2 gene transfer was associated with increased peak systolic Ca2+ transient amplitude and rate of Ca2+ decline and increased SERCA2a expression. In addition, UCn2 gene transfer reduced Thr286 phosphorylation of Cam kinase II, and increased expression of cardiac myosin light chain kinase, findings that would be anticipated to increase function of the failing heart. We conclude that a single IV injection of AAV8.UCn2 increases function of the failing heart. The simplicity of IV injection of a vector encoding a gene with beneficial paracrine effects to increase cardiac function is an attractive potential clinical strategy.
Introduction
C
Current methods of gene transfer for heart diseases include intramuscular (IM) injection into heart muscle and intracoronary delivery, which are cumbersome to apply. Gene transfer trials for heart disease have thus far been disappointing perhaps because of inadequate levels of gene expression in the heart. To circumvent this impediment, we have proposed a different approach: intravenous (IV) delivery of a vector encoding a transgene with paracrine activity affecting the heart. In this strategy, a peptide with beneficial cardiovascular effects is released to the circulation from distant sites after systemic delivery of a long-term expression vector encoding the peptide. This approach enables gene transfer via a simple IV injection during an office visit, circumvents the need for more invasive delivery methods, and potentially may give gene transfer for heart failure a much needed boost.
Clinical trials of systemic delivery of genes are underway for hemophilia B 2,3 and α1-antitrypsin deficiency 4 and others are in development. However, systemic delivery of vectors encoding peptides with beneficial paracrine activity has not been performed in clinical CHF trials, and is uncommon in preclinical studies. 5,6 Intravenous infusion of potentially beneficial peptides such as urodilatin, 7 relaxin-2, 8 urocortin-2 (UCn2), 9,10 and stresscopin 11 shows promise in clinical CHF trials, but gene transfer of these peptides has not been attempted.
Urocortins 1, 2, and 3 (38–41 amino acids) belong to the corticotropin-releasing factor (CRF) family. These peptides stimulate CRF receptors 1 and 2 (CRFR1 and CRFR2). UCn1 binds to CRFR1 and CRFR2, but UCn2 and UCn3 exclusively bind to CRFR2, which is expressed in cardiac myocytes, vasculature, gut, brain, and skeletal muscle. 12 –14 Through selective CRFR2 activation, UCn2 mediates protean beneficial effects, including reduced renin-aldosterone activation, and is a potent inotrope with minimal effects on cardiac cAMP. 15 In preclinical and clinical HF, peptide infusions of UCn2 or stresscopin, a related peptide, have shown favorable effects on left ventricular (LV) function. 9 –11,16
Since plasma half-life of UCn2 is 15 min, 10 chronic infusion is required for sustained effects. The inconvenience, need for hospitalization and associated hazards, and the expense of therapeutic peptide infusions for CHF are considerable, and impede the broad use of these otherwise attractive peptides. The present study was conducted to address and resolve these shortcomings using UCn2 gene transfer in a murine model of CHF.
Adeno-associated virus (AAV) vectors enable prolonged transgene expression. For example, persistent transgene expression has been shown in nonhuman primates for sustained periods after IM 17 or IV delivery of AAV vectors. 18 –20 We have confirmed this in rats. 6 Although recent clinical trials have found that some AAV serotypes incite immune responses after IM injection, 21,22 newer generation AAV vectors (AAV5, 6, 8, 9, rh-10) do not appear to have similar problems in nonhuman primates. 19,20,23,24 IV AAV delivery is superior to IM vis-à-vis serum transgene levels, and AAV9 and AAV8 are superior to AAV5. 25 Moreover, preexisting anti-AAV8 antibodies are not as prevalent in humans (19%) as are other AAV serotypes, including AAV1 and AAV2 (50–59%). 26 These data suggest that IV AAV8 may be an attractive delivery route and vector to attain sustained increased levels of plasma UCn2. IV delivery of an AAV vector encoding a paracrine gene, as compared with IV peptide infusion, has the potential to circumvent infection and reduce repeated and prolonged hospital stays, thereby reducing costs. Systemic vector delivery may be an advantage in this approach—it provides the highest level of expression for any given AAV dose—by exploiting widespread distribution of the vector.
In a previous study we showed that a single IV injection of AAV8.UCn2 increases plasma UCn2 and increases LV systolic and diastolic function for at least 7 months in normal mice. 27 The present study was conducted to test the hypothesis that IV delivery of AAV8.UCn2 will increase function of the failing heart.
Methods
AAV8.UCn2 vector production
A helper virus-free AAV8 vector encoding murine UCn2 driven by a chicken β-actin (CBA) promoter (AAV8.CBA.UCn2; Fig. 1) was produced by transient transfection of HEK293T cells with the vector plasmid pRep2/Cap8 and pAd-Helper plasmid. 28 Plasmid pRep2/Cap8 was obtained from the University of Pennsylvania Vector Core. Cell lysates prepared after 72 hr of transfection were treated with benzonase and viruses were consolidated through 25% sucrose-cushion ultracentrifugation. The pellets were resuspended for further purification of the virus through anion-exchange column chromatography (Q-Sepharose; GE Health Science) and concentrated by 25% sucrose-cushion ultracentrifugation. 29,30 Subsequently, the pellets were resuspended in 10 mM Tris-HCl (pH 7.9, 1 mM MgCl2, 3% sucrose). Virus titers were determined by real-time qPCR with virus genome DNA prepared from purified virus.
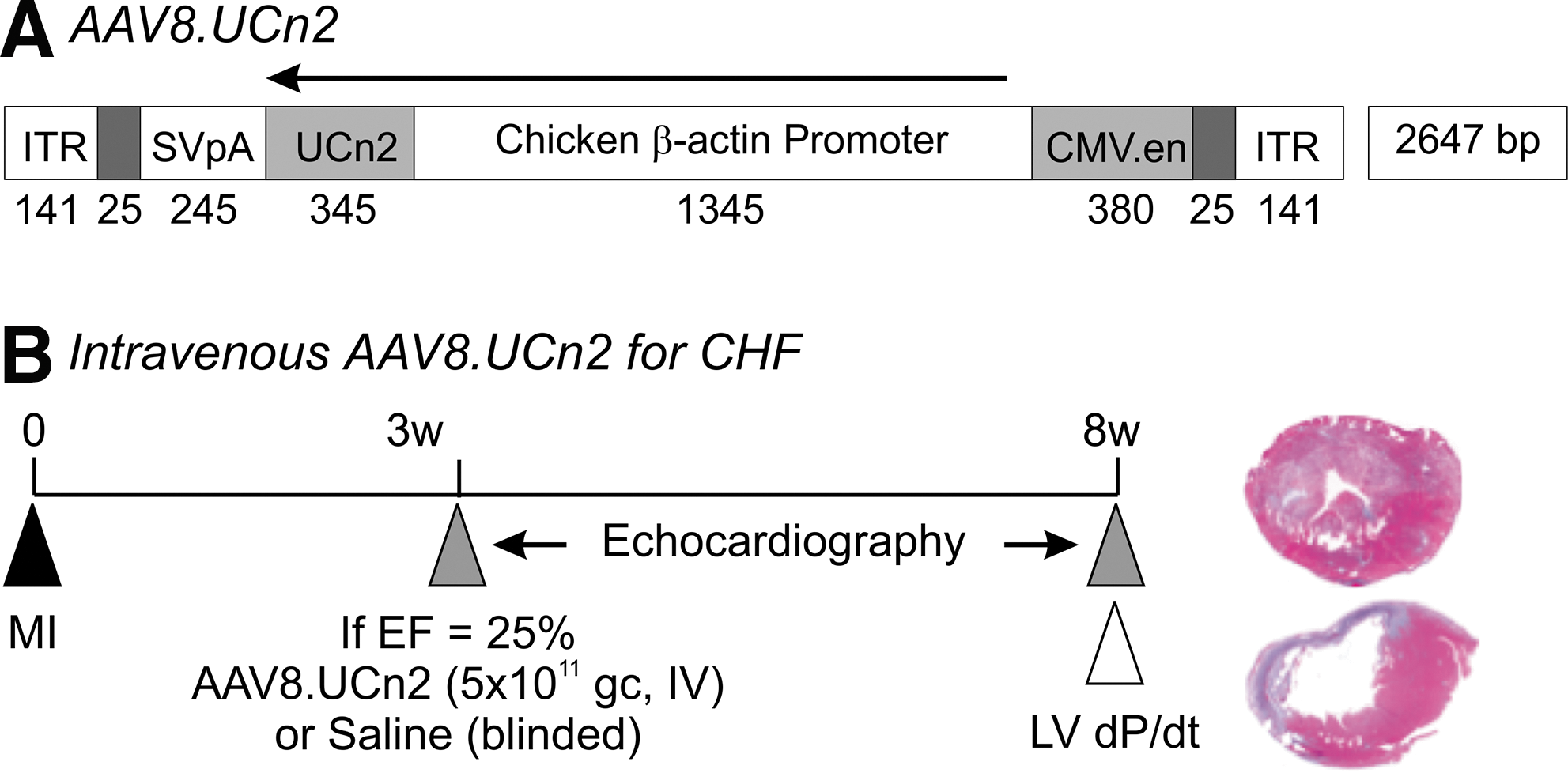
AAV8.CBA.UCn2 map and experimental protocol.
Heart failure model
The Animal Use and Care Committee of the VA San Diego Healthcare System approved the studies. Two hundred thirty-one male C57BL/6J mice (Jackson Laboratories) aged 10–12 weeks, weighing 26.1±0.2 g, were used. We used coronary occlusion to induce large anterior wall myocardial infarction (MI) and CHF as described in detail previously. 31,32 MI size deliberately was large—50% of LV, comprising most of the LV free wall (Fig. 1). Consequently, this model is associated with a high initial mortality. Of 231 mice that underwent coronary occlusion, 125 (54%) died before randomization (AAV8.UCn2 or saline) primarily in the first few days after MI. An additional 45 mice (19%) did not show sufficient LV dysfunction 3 weeks after MI to be randomized. Sixty-one mice (26%) had sufficiently low LV ejection fractions (EF <25%) and were randomized, and 11 of these mice died before the final study 5 weeks after randomization: 4 UCn2 (mortality 13%) and 7 saline (mortality 23%). The primary end point was LV function 5 weeks after IV delivery of AAV8.UCn2 versus saline in mice with severe heart failure (Fig. 1). Data were acquired and analyzed without knowledge of group identity.
AAV8.UCn2 delivery
Under anesthesia (1.5% isoflurane via nose cone), a small incision was made to expose the jugular vein for IV delivery of AAV8.UCn2 (5×1011 genome copies [gc] in 100 μl) or a similar volume of saline (control).
Effects of UCn2 gene transfer on heart rate and blood pressure
These studies were conducted to assess the effects of UCn2 gene transfer on heart rate and blood in unsedated mice with heart failure. Impaired LV EF was confirmed 3 weeks after MI, and mice received IV AAV8.UCn2 (5×1011 gc) or saline. Systolic and diastolic blood pressure and heart rate were measured by tail cuff (Visitech Systems) in unsedated mice.
Echocardiography
Echocardiography was performed as previously described. 33 Echocardiography was performed 3 weeks after MI to document reduced LV function (EF <25%) and to record LV chamber dimensions. Echocardiographic assessment was then repeated 5 weeks after randomization of mice to receive IV delivery of AAV8.UCn2 or saline.
LV systolic and diastolic function
Mice were anesthetized with sodium pentobarbital (80 mg/kg, ip) and a 1.4F conductance-micromanometer catheter (SPR 839; Millar Instruments) was advanced via the right carotid artery across the aortic valve and into the LV cavity. LV pressure was recorded and stored digitally for processing (IOX1.8; Emka Technologies) as previously reported. 6 Subsequently, blood and tissue samples were obtained. After acquisition, the first derivatives of LV pressure development (LV +dP/dt) and decline (LV –dP/dt) were used to assess LV systolic and diastolic function. Data were acquired and analyzed without knowledge of group identity.
Cardiac myocyte isolation
Cardiac myocytes were isolated as previously described. 33
Ca2+ transients
Cytosolic Ca2+ transients were measured using Indo-1 as described previously 27,34 with modifications. Cardiac myocytes were plated onto laminin-coated glass cover slips and loaded with indo-1/AM (3 μM; Calbiochem) and dispersing agent, pluronic F-127 (0.02 mg/ml; Calbiochem), for 30 min. Following dye loading, cover slips were mounted in a superfusion chamber, rinsed to remove excess indo-1-AM, and mounted on a Nikon Diaphot epifluorescence microscope equipped with a 40×objective interfaced to a Photon Technologies photometry system with the excitation wavelength set to 365 nm via a monochromator. Fluorescence emission was split and directed to two photomultiplier tubes through 20 nm band-pass filters centered at 405 and 485 nm, respectively. The ratio F405/F485 represents a measure for [Ca2+]i. During these measurements, cardiac myocytes were superfused with 25 mM HEPES (pH 7.3) containing 2 mM CaCl2. Myocytes were field-stimulated at 0.3 Hz. Ca2+ transients were recorded from 144 cardiac myocytes obtained from 6 hearts (3 per group). Diastolic and systolic intracellular Ca2+ levels were inferred from the basal and maximal indo-1 ratio per cycle, respectively. Diastolic decay time (tau) was calculated from the normalized Ca2+ transient.
Quantitative RT-PCR and immunoblotting
LV and liver samples were collected and stored at −80°C for quantitative RT-PCR and Western blotting.
Quantitative RT-PCR
LV and liver RNA was isolated using RNeasy mini kit (Qiagen) and qRT-PCR conducted as previously described 27 under the following conditions: 5 min at 98°C, 40 cycles of 30 s at 95°C, 30 s at 55°C, and 30 s at 72°C. RNA equivalents were normalized to simultaneously determined glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA levels in each sample. Primers are listed in Table 1.
ANF, atrial natriuretic peptide; BNP, brain natriuretic peptide; Coll, collagen; MEF2, myocyte enhancer factor-2; MMP, matrix metallopeptidase; TIMP, tissue inhibitor of metalloproteinases; UCn2, urocortin 2; α-Cd-actin, alpha-cardiac actin; α-MHC, alpha-myosin heavy chain; α-SK-actin, alpha-skeletal actin; β-MHC, beta-myosin heavy chain.
Immunoblotting
Immunoblotting was performed as described previously. 35 The following antibodies were used: cMLCK (Abgen/Thermo Scientific); p286 CamKII (Santa Cruz); phospho-PKA catalytic subunit, PKA catalytic subunit, troponin I (TnI), and 22/23-phospho-TnI (Cell Signaling Technology); phospholamban (PLB) (Thermo Fisher Scientific); Ser 16 and Thr 17-phospho-PLB (Badrilla, Ltd.); and SERCA2a (Enzo Life Sciences).
Cyclic AMP and protein kinase A activity
Transmural LV samples underwent cAMP measurement before and after stimulation with isoproterenol (10 mM, 10 min) and NKH477 (10 mM, 10 min), and cAMP was measured using the Biotrak Enzymeimmunoassay System (GE Healthcare) as previously described. 36 PKA activity was determined as previously described. 27 Cardiac myocytes underwent cAMP measurement before and after stimulation with isoproterenol (10 μM, 10 min) and NKH477 (10 μM, 10 min) and subsequently homogenized in buffer A, 20 mM Tris-HCl (pH 7.4), 0.5 mM EGTA, 0.5 mM EDTA, and protease inhibitor cocktail (Invitrogen), and centrifuged (14,000×g, 5 min, 4°C). The supernatant was incubated with PKA biotinylated peptide substrate (SignaTECT cAMP-Dependent Protein Kinase Assay System; Promega) in the presence of [γ-32P]ATP. The 32P-labeled biotinylated substrate was recovered with a streptavidin matrix and the specific activity of PKA determined.
Histology
Samples of liver and transmural sections of the uninfarcted LV septum were formalin fixed and paraffin embedded. Five-micrometer sections were mounted and counterstained with hematoxylin and eosin and with Masson's trichrome. For quantitative assessment of LV fibrosis, images of a short-axis midwall LV ring were obtained with a Nikon Eclipse Ti-U microscope. Blinded analysis of the degree of fibrosis in the viable LV region (excluding the infarcted region) was conducted using NIS-Elements AR 3.10 software (Nikon Inc.). A similar analytical process was performed on fixed and counterstained liver samples.
Statistical analysis
Data represent mean±SE; group differences were tested for statistical significance with ANOVA followed by Bonferroni t-test. Between-group comparisons were made using Student's t-test (unpaired, two-tailed). The null hypothesis was rejected when p<0.05.
Results
Heart rate and blood pressure in unsedated mice
No group differences were seen in heart rate or systolic, diastolic, or mean arterial blood pressure 5 weeks after UCn2 gene transfer (Table 2), although heart rates tended to be quite high in the untreated group and closer to normal in mice that had received UCn2 gene transfer.
The effects of UCn2 gene transfer on blood pressure and heart rate (HR) were assessed in unsedated mice with heart failure (HF) 5 weeks after UCn2 gene transfer (HF+UCn2, 5×1011 gc, IV) or IV saline (HF). Systolic and diastolic blood pressure was measured by tail cuff and mean blood pressure calculated. No group differences were seen in heart rate or blood pressure. Values denote mean±SE; p-values are from Student's t-test (unpaired, 2-tailed).
Urocortin 2 expression
Five weeks after IV delivery of AAV8.UCn2 (5×1011 gc; n=6), UCn2 mRNA was increased 15,263-fold in liver (p<0.0001) and 70-fold in LV (p=0.03) versus endogenous UCn2 mRNA.
Echocardiography
Intravenous delivery of AAV8.UCn2 to mice with HF was associated with increased EF (p=0.01), and velocity of circumferential fiber shortening was increased but did not reach statistical significance (p=0.09). Mice that received AAV8.UCn2 also exhibited reductions in LV end-diastolic diameter (EDD; p<0.001) and LV end-systolic diameter (ESD; p=0.002). The saline-treated mice showed an 11% increase in LV EDD, while the UCn2-treated group showed a 2% decrease in LV EDD. Similarly, the saline group showed a 16% increase in LV ESD, while the UCn2 group experienced a 6% reduction. Although these changes in LV dimension are small, volume is a cubic function of dimension, so the changes are considerable—a calculated 64% increase in ESD (saline vs. UCn2) and a 46% increase in EDD (saline vs. UCn2). Posterior and septal wall thickness showed no group differences (Table 3).
bpm, beats per minute; EDD, LV end-diastolic diameter; ESD, LV end-systolic diameter; HF, heart failure; HR, heart rate; IVS Th, interventricular wall thickness at end-diastole; LV EF, left ventricular ejection fraction; Post–Pre, the value 5 weeks after saline or UCn2 gene transfer minus the value before; PW Th, posterior wall thickness at end-diastole; UCn2, urocortin-2; VCFc, velocity of circumferential fiber shortening (corrected for heart rate).
p-Values from Student's t-test (paired data, 2-tailed) for group difference in change, Post-Pre.
LV systolic and diastolic function
In vivo assessment of LV pressure development showed substantial increases in rates of LV pressure development (LV +dP/dt; p<0.0001) and in LV relaxation (LV −dP/dt; p<0.0007) (Fig. 2 and Table 4). There were no group differences in mean arterial pressure (Table 4). Heart rate during these studies, conducted under anesthesia, was somewhat higher in mice that had received UCn2 gene transfer, but the difference did not reach statistical significance.
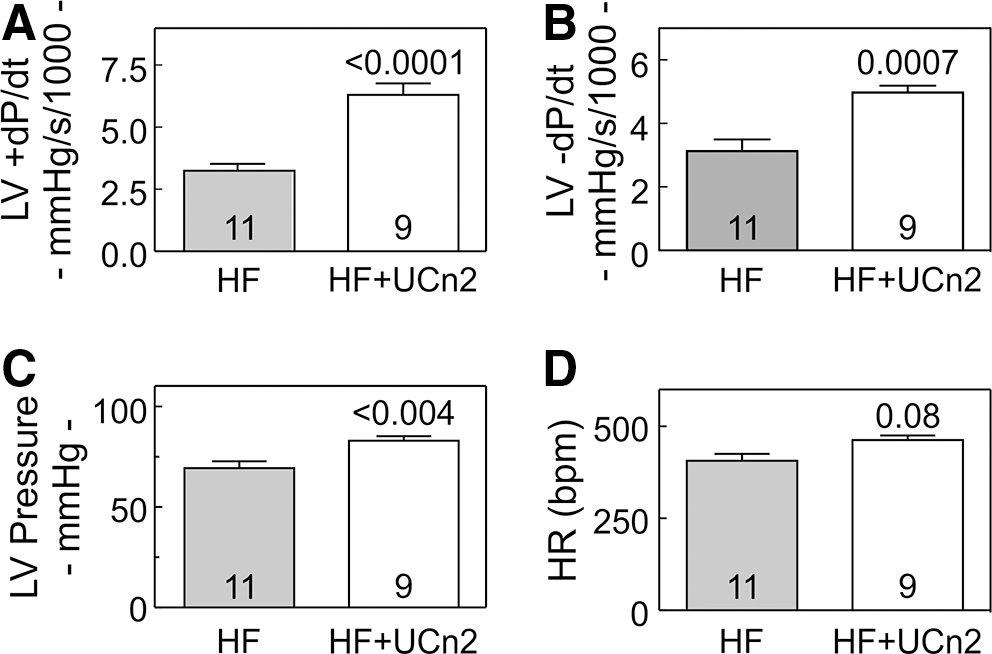
LV function in vivo.
HR, heart rate; LV, left ventricle; LVP, left ventricular developed pressure; MAP, mean arterial pressure; UCn2, urocortin-2 gene transfer.
Three weeks after myocardial infarction, mice received intravenous saline or AAV8.UCn2 (5×1011 gc). Mice underwent physiological studies 5 weeks later. Values represent mean±SE. p-Values are from Student's t-test (unpaired, 2-tailed).
Cytosolic Ca2+ transients and related genes
Basal Ca2+ released (systolic–diastolic Ca2+) was increased in cardiac myocytes from heart failure mice that had received UCn2 gene transfer (p=0.0001; Fig. 3A and B). UCn2 gene transfer was also associated with a reduced Ca2+ decline time (t ½, Tau) in cardiac myocytes from mice with heart failure 5 weeks after UCn2 gene transfer p=0.001; Fig. 3C and D). Increased UCn2 was associated with increased expression of SERCA2a mRNA and protein in normal and failing LV (Fig. 3E). However, no group differences were seen in LV protein expression and phosphorylation of PLB or TnI (data not shown).
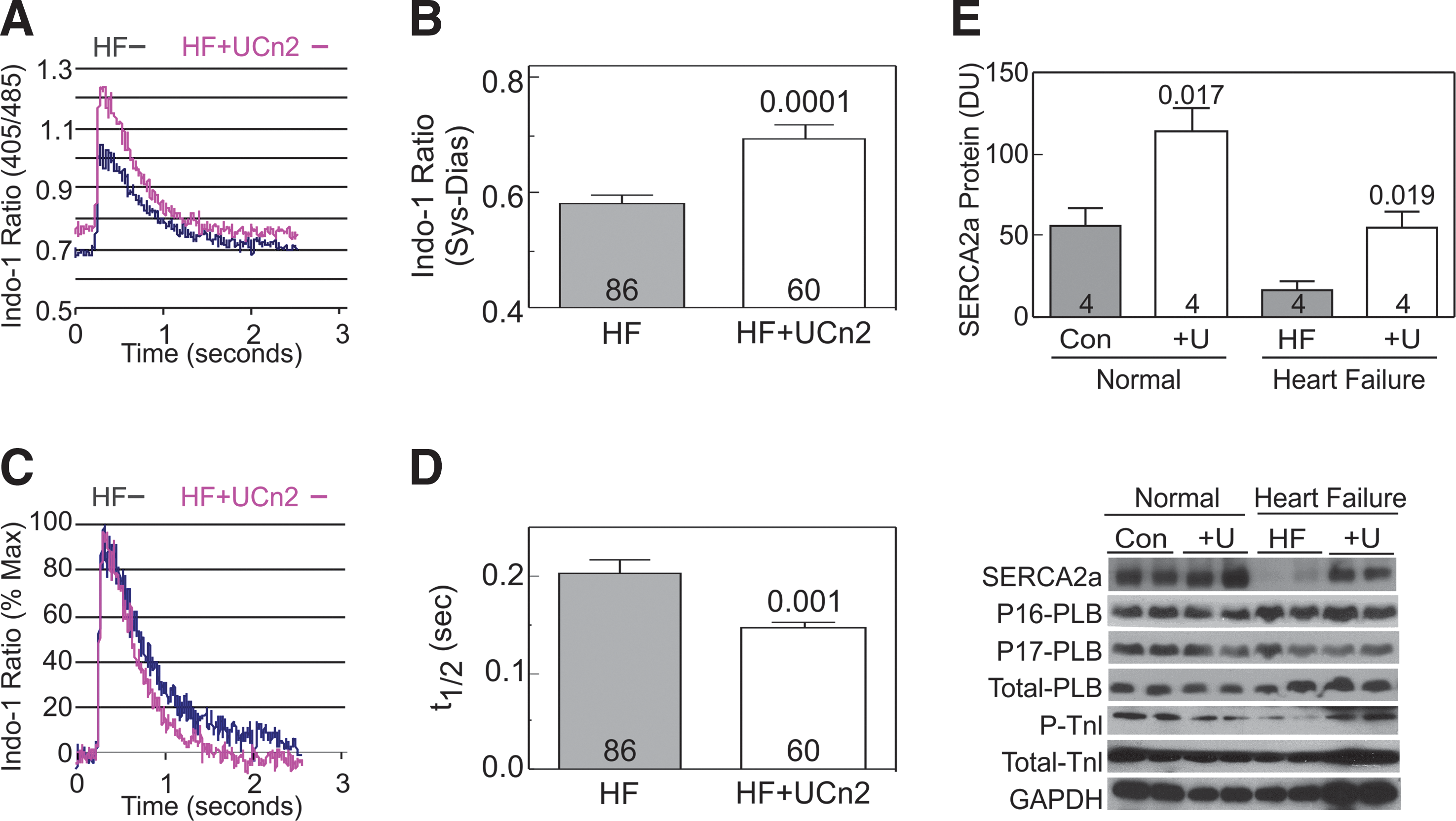
Cytosolic Ca2+ transients in cardiac myocytes from mice with heart failure (HF) 5 weeks after IV AAV8.UCn2 (HF+UCn2) or IV saline.
Cyclic AMP and PKA activity
LV samples and cardiac myocytes isolated from hearts of both groups showed no differences in cAMP or PKA activity (Fig. 4). Cyclic AMP production and PKA activity were assessed before and after stimulation with isoproterenol or NKH477, a water-soluble forskolin analog that stimulates adenylate cyclase independently of β-adrenergic receptors. No group differences were seen in basal, Iso, or NKH477-stimulated cAMP production (Fig. 4A) or in PKA activity (Fig. 4B). Expression of PKA family proteins (catalytic α unit and regulatory α and β subunits and their phosphorylation) was not altered (data not shown).
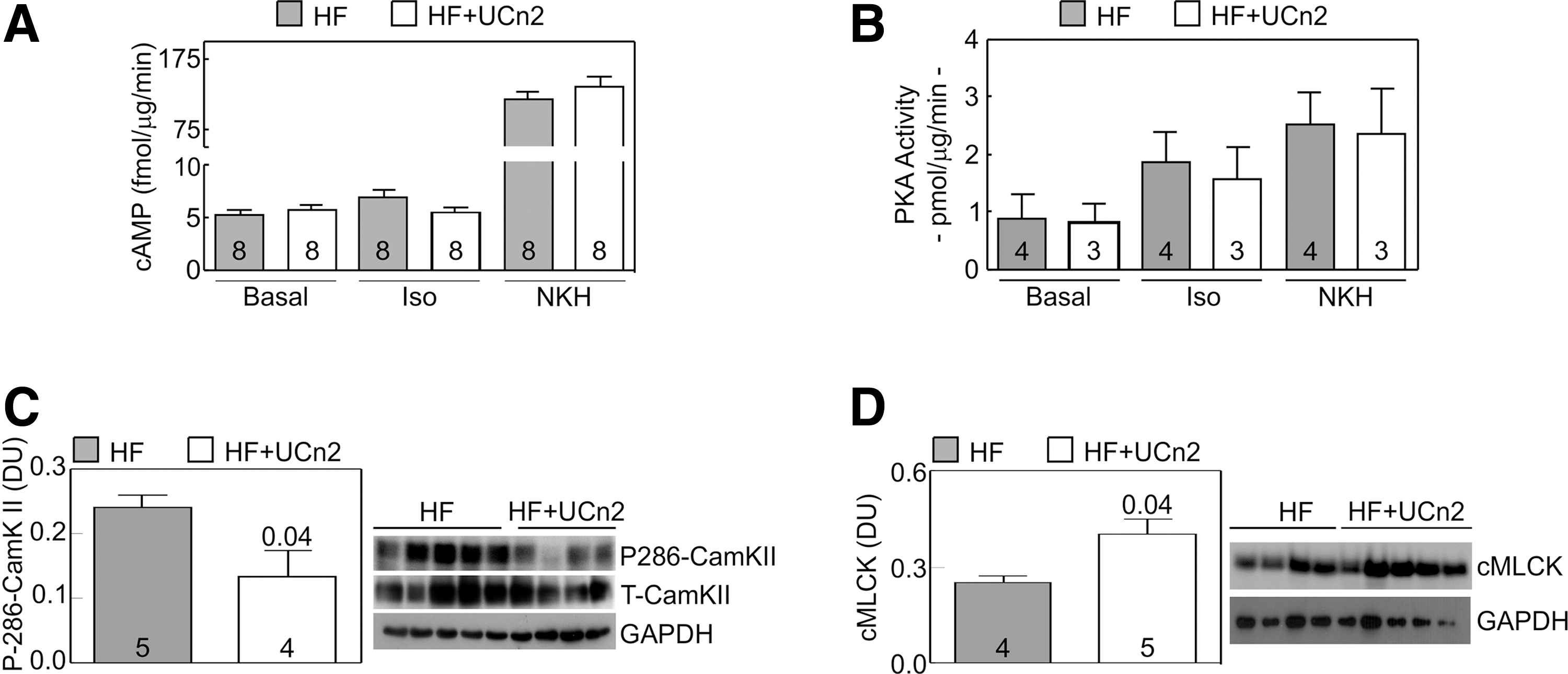
Cardiac myocyte cAMP-PKA signaling. LV samples
CamKII and cMLCK
To seek mechanisms to explain increased function of the failing heart evoked by UCn2 gene transfer, we measured LV expression and phosphorylation of calcium/calmodulin-dependent protein kinase II (CamKII) and expression of cardiac myosin light chain kinase 3 (cMLCK). CamKII phosphorylation at Ser286 was reduced in LV samples from HF mice after UCn2 gene transfer (47% reduction, p=0.04; Fig. 4C), although total CamKII protein expression showed no group difference. Seeking alterations in myofilament sensitivity to Ca2+, we assessed LV cardiac myosin light chain kinase 3 (cMLCK) expression after UCn2 gene transfer, finding a 1.6-fold increase (p<0.04) (Fig. 4D).
Necropsy
Liver, lung, and body weights showed no group differences. UCn2 gene transfer tended to reduce LV weight, and LV-to-body weight ratio was reduced (12% reduction; p=0.01) (Table 5).
BW, body weight; LV, left ventricle; UCn2, urocortin-2 gene transfer.
Three weeks after myocardial infarction, mice received intravenous saline or AAV8.UCn2 (5×1011 gc). Mice were killed 6 weeks later and necropsy conducted. Values represent mean±SE. p-Values are from Student's t-test (unpaired, 2-tailed).
Markers of stress, inflammation, and tissue injury
The expression of several markers of LV stress, inflammation, and tissue injury was examined using RT-PCR. HF altered the expression of most of these genes (Table 6). Increased UCn2 expression did not influence alterations associated with HF. However, in normal mice, increased UCn2 expression was associated with reduced expression of ANF (p=0.007), BNP (p=0.01), β-MyHC, and α-SK-actin (p=0.03).
ANF, atrial natriuretic peptide; BNP, brain natriuretic peptide; Coll, collagen; MEF2, myocyte enhancer factor-2; MMP, matrix metallopeptidase; TIMP, tissue inhibitor of metalloproteinases; α-Cd-actin, α-cardiac actin; α-MHC, α-myosin heavy chain; α-SK-actin, α-skeletal actin; β-MHC, β-myosin heavy chain.
RT-PCR detection of gene expression in LV tissues of normal and HF mice. RNA equivalents were normalized to simultaneously determined glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA levels in each sample and data are normalized to control in normal LV.
LV and liver histology
Hematoxylin and eosin staining of samples of liver and LV showed no evidence of group differences (data not shown). Masson's trichrome staining revealed no group differences in fibrosis in liver (p=0.79).
Discussion
The most important finding in this study is that a single IV injection of AAV8.UCn2 increased function of the failing heart, demonstrating the feasibility and effectiveness of IV delivery of a long-term expression vector encoding a peptide with beneficial paracrine effects to treat heart failure.
Two measures of cardiac function confirmed increased LV function 5 weeks after IV AAV8.UCn2 delivery to animals with severely dysfunctional left ventricles. Echocardiography showed increases in LV EF, and reductions in LV volumes (Table 3). Second, UCn2 gene transfer increased peak LV +dP/dt, indicating enhanced LV contractile function, and reduced LV–dP/dt, indicating enhanced LV diastolic function (Table 4 and Fig. 2).
Although the absolute degree of LV EF change was only 8% units (HF: 12%±1%; HF+UCn2: 20%±4%), the relative increase was 67%. The small absolute change reflects the large size of the infarction—the mean prerandomization LV EFs were ≤20% in both groups. Despite such large infarctions, UCn2 gene transfer attenuated LV chamber dilation and increased EF, while saline-treated mice showed progressive LV chamber dilation and further deterioration of LV EF. One would not expect UCn2 gene transfer to remedy the problems associated with such a large area of scar, representing virtually the entirety of the LV free wall. The cardiac benefits of UCn2 gene transfer would be anticipated to be limited to the viable portion of the LV, which, in the current model, represents the interventricular septum. EF in this setting may underestimate the benefits on LV function, especially since we observed dyskinesia of the infarcted wall during ejection. Assessment of LV contractile function using peak LV +dP/dt reveals a larger absolute increase in LV function—an increase of 3129 mmHg/sec in peak LV +dP/dt, and a 1847 mmHg/sec increase in peak −dP/dt conferred by UCn2 gene transfer. These represent a 2-fold increase in peak LV +dP/dt, and a 1.6-fold increase in peak −dP/dt. A doubling of peak LV +dP/dt in clinical heart failure would normalize LV contractile function. 37,38
Heart rate and blood pressure in the unsedated state are not affected by IV delivery of AAV8.UCn2 despite sustained high levels of transgene UCn2 in normal mice 27 or in mice with CHF, as shown in the current study. Similarly, in clinical trials of peptide infusions of UCn2 and stresscopin (similar to UCn3), the rate-pressure product is unchanged. 9 –11 One would, therefore, not anticipate an increase in cardiac metabolic demands associated with UCn2 gene transfer, but more direct metabolic studies must be performed to know this with certainty.
The present study focused on the feasibility and physiological consequences of IV delivery of AAV8.UCn2 in the setting of a severely compromised and failing heart, and we found a pronounced positive effect. The mechanisms by which UCn2 gene transfer evoked beneficial physiological changes, although not the primary focus of the present study, were also examined.
For example, we found that UCn2 gene transfer was associated with (1) increased peak systolic Ca2+ transient amplitude and increased rate of Ca2+ decline in cardiac myocytes isolated from HF mice (Fig. 3A–D), and (2) increased SERCA2a expression (Fig. 3E) as we previously reported in mice with normal hearts. 27 Increased LV SERCA2a expression provides a mechanism by which LV contractile function and relaxation would be increased, as was observed (Fig. 2). SERCA2a returns cytosolic Ca2+ to the sarcoplasmic reticulum. An increased amount of SERCA2a would be anticipated to yield a more rapid cytosolic Ca2+ decline, which is what we found (Fig. 3C and D), and consequently to increase the rate of LV pressure decline (LV −dP/dt), as we also found (Fig. 2B).
In addition, we found alterations in LV expression of two additional proteins that are likely to have been of mechanistic importance in the observed beneficial effects of UCn2 gene transfer on function of the failing LV: reduced Thr286 phosphorylation of Ca2+/calmodulin-dependent kinase II (CaMKII), and increased LV expression of cardiac myosin light chain kinase (cMLCK) (Fig. 4).
CaMKII Thr286 phosphorylation
Our data show that UCn2 gene transfer was associated with reduced Thr286 phosphorylation of CaMKII (Fig. 4C). CaMKII expression and activation are important determinants of cardiac function. 39 For example, cardiac-directed expression of CaMKII results in heart failure in mice. 40 Others have shown increased CaMKII activity and expression in MI-induced heart failure in mice. 41 The clinical relevance of these findings was demonstrated recently by the demonstration that inhibition of LV CaMKII increases function of the failing human heart. 42 Although we speculate that reduced Thr286 phosphorylation of CaMKII may have been important mechanistically in the observed increase in LV function, we were unable to determine the pathway by which increased UCn2 reduces Thr286 CaMKII phosphorylation, which will require focused studies in cultured cardiac myocytes that are underway.
Cardiac myosin light chain kinase
We found increased cardiac myosin light chain kinase (cMLCK) expression associated with UCn2 gene transfer (Fig. 4D). Phosphorylation of cardiac myosin light chain 2v by cMLCK increases the rate of cross-bridge recruitment in cardiac myocytes and influences contractile function. 43,44 Increased levels cMLCK are associated with increased LV function in the setting of MI-induced heart failure. 45 In contrast, the deletion of cMLCK reduces cardiac performance. 46 Sadly, there is no antibody available to assess myosin light chain 2v phosphorylation, and so the biological importance of the increase in cMLCK associated with UCn2 gene transfer in the present study must remain speculative.
UCn2 gene transfer was associated with a doubling in the peak rate of LV pressure development (LV +dP/dt; Table 4 and Fig. 2). This finding was supported by evaluation of LV dimension and function by echocardiography (Table 3), enhanced Ca2+ handling (Fig. 3), and signaling changes in LV predicted to increase contractile function, including increased SERCA2a protein expression (Figs. 3 and 4). Because of the consistency of these findings, which reverberated from isolated cardiac myocytes to in vivo physiology, we were less concerned by the absence of group differences in BNP and ANF mRNA in LV (Table 6). Perhaps plasma levels or BNP/ANF expression in LA would have revealed group differences that LV mRNA levels missed. It is also possible that despite increased LV contractile function there was sufficient persistent chamber dilation—owing to infarction of the entire LV free wall—to provide ongoing stimulation of ANF and BNP expression.
We saw no group difference in lung or liver weight (Table 5). Liver weights were not increased in mice with heart failure compared with normal mice, 27 and so, despite severe LV failure, there is no liver congestion. Whether this is unique to MI-induced CHF in mice is unknown. Lung weights increased by 23% versus normal age-matched mice, 27 but did not show a group difference. We speculate that despite a doubling of LV contractile function (peak +dP/dt) conferred by UCn2 gene transfer (Table 4 and Fig. 2), there may have been persistent left-sided congestion 5 weeks after treatment.
Clinical application
Intravenous delivery of AAV8 enables transfection of many organs and is especially effective in liver, skeletal muscle, and heart. 47 These organs, because they comprise an enormous mass of tissue and therefore can release abundant transgene UCn2, will enable us to reduce the vector dose. Indeed, a vector dose 10-fold lower (5×1010 gc per mouse or 2×1012 gc/kg) is still effective in increasing LV +dP/dt. 27 A dose of 2×1012 gc/kg of AAV8 encoding human factor IX was delivered intravenously safely and effectively in a clinical trial in subjects with hemophilia B. 2
An additional feature to consider in translating our findings to clinical applications is the use of a regulated expression system, 5,6 which would enable turning UCn2 expression on or off at will. We have designed such AAV8 vectors using tetracycline and rapamycin regulation systems and are conducting preclinical studies with these regulated expression vectors.
LV Ca2+ handling is different in humans than in mice, 48 but peptide infusions of UCn2 or stresscopin (similar to UCn3) in patients with HF increases LV function. 9 –11 Whether this is through Ca2+ handling is unknown because Ca2+ transients and Ca2+ handling proteins have not been assessed in cardiac myocytes or myocardium before and after UCn2 peptide infusions in humans.
Finally, now that we have demonstrated that UCn2 gene transfer increases function of the severely failing heart, it will be important to determine how long the effect persists and whether it reduces mortality. Such studies using a less severe model of CHF with better long-term survival are planned.
Conclusions
A single IV injection of AAV8.UCn2 increases both systolic and diastolic function of the severely failing heart. Systemic delivery of the vector ensures that the transgene is expressed in the heart, but also is continuously released into the circulation, thereby providing sustained benefits that would otherwise not be possible. Other theoretical advantages of gene transfer as compared with IV infusion of paracrine-acting peptides include reduction in catheter-based infections, no need for hospitalization, and reduced costs. Whether this strategy will be effective in increasing function of the failing heart in clinical settings will require additional studies.
Footnotes
Acknowledgments
This work was supported by National Institutes of Health grants (P01 HL66941, HL088426, HL081741, and HL107200) and Merit grants from the Department of Veteran's Affairs (1101BX001515 and 1101bBX000783).
Author Disclosure Statement
H.K.H. is founder, unpaid consultant, and equity holder of Renova Therapeutics. Renova did not fund the work and was not involved in its planning, interpretation, or writing. None of the other authors have disclosures.