
Abstract
The Campylobacter genus is the leading cause of human gastroenteritis, with the consumption of contaminated poultry meat as the main route of infection. Probiotic bacteria, such as Lactobacillus, Bacillus, Escherichia coli Nissle, and Bifidobacterium species, have a great immunomodulatory capacity and exhibit antipathogenic effects through various molecular mechanisms. Reducing Campylobacter levels in livestock animals, such as poultry, will have a substantial benefit to humans as it will reduce disease transmissibility through the food chain. Moreover, probiotic-based strategies might attenuate intestinal inflammatory processes, which consequently reduce the severity of Campylobacter disease progression. At a molecular level, probiotics can also negatively impact on the functionality of various Campylobacter virulence and survival factors (e.g., adhesion, invasion), and on the associated colonization proteins involved in epithelial translocation. The current review describes recent in vitro, in vivo, and preclinical findings on probiotic therapies, aiming to reduce Campylobacter counts in poultry and reduce the pathogen's virulence in the avian and human host. Moreover, we focused in particular on probiotics with known anti-Campylobacter activity seeking to understand the biological mechanisms involved in their mode of action.
Introduction
Members of the genus Campylobacter are the utmost documented foodborne pathogens of the present century (Śmiałek et al, 2021). Being first isolated in 1963, Campylobacter is renowned for its exquisite capacity of adherence and biofilm formation on different surfaces, high invasion of various hosts, and for its increased viability outside of its natural biological niche (Erega et al, 2021a). Adhesion is one of the most important virulence factors required for Campylobacter survival during host–pathogen interaction being correlated with the animal and human infection rates (Šikić Pogačar et al, 2020). Campylobacter jejuni and Campylobacter coli are the most representative species of the Campylobacter genus known for their ability to form mono- or multispecies biofilms (Elgamoudi and Korolik, 2021).
These two species are responsible for causing 90% of the estimated human campylobacteriosis cases and are accountable for ≈84% (C. jejuni) and ≈9% (C. coli) of the total diagnosed cases in Europe (Soro et al, 2020). To successfully invade the human intestine and cause disease, the ingestion of only a few hundred bacterial cells is sufficient to initiate the Campylobacter infection, with an incubation period that varies from 24 to 72 h before the onset of illness (Soro et al, 2020). Campylobacter-induced disease is usually followed by postacute infection sequelae such as bacteremia, urinary tract infections, sepsis, and complicated immune-mediated neuropathies (Hayat et al, 2022) (e.g., Guillain–Barré and Miller-Fisher syndromes). These neuropathies are characterized by neuromuscular paralysis (Itamura et al, 2022), posing significant human health risks (Balta et al, 2021b; Elgamoudi and Korolik, 2021).
Poultry flocks are carriers of high levels of campylobacters (≈109), which makes consumption of contaminated poultry meat the main source of infection in humans (Rawson et al, 2022). Henceforth, controlling Campylobacter levels at primary production represents a major step in preventing human infections; however, no single control method is yet capable of complete pathogen elimination (Hakeem and Lu, 2021). Probiotics are considered a viable tool for pathogen reduction (Cean et al, 2015) as they can limit the use of antibiotics and improve animal performance with the added benefit of antibiotic-free food products availability for human consumption (Sibanda et al, 2018). One of the probiotic positive impacts on gut microbiota is the endogenous production of short-chain fatty acids inducing immunostimulatory effects in farm animals (Melara et al, 2022).
Nevertheless, a more focused approach and a more profound understanding on how to standardize probiotic use in livestock and on their antipathogen efficacy are required. Based on the main effects of probiotics presented in Figure 1, in this study, we aim to discuss the latest advances in probiotic use as a strategy to reduce the pathogen virulence and transmissibility, in with a special focus on poultry but tangential to human infections.
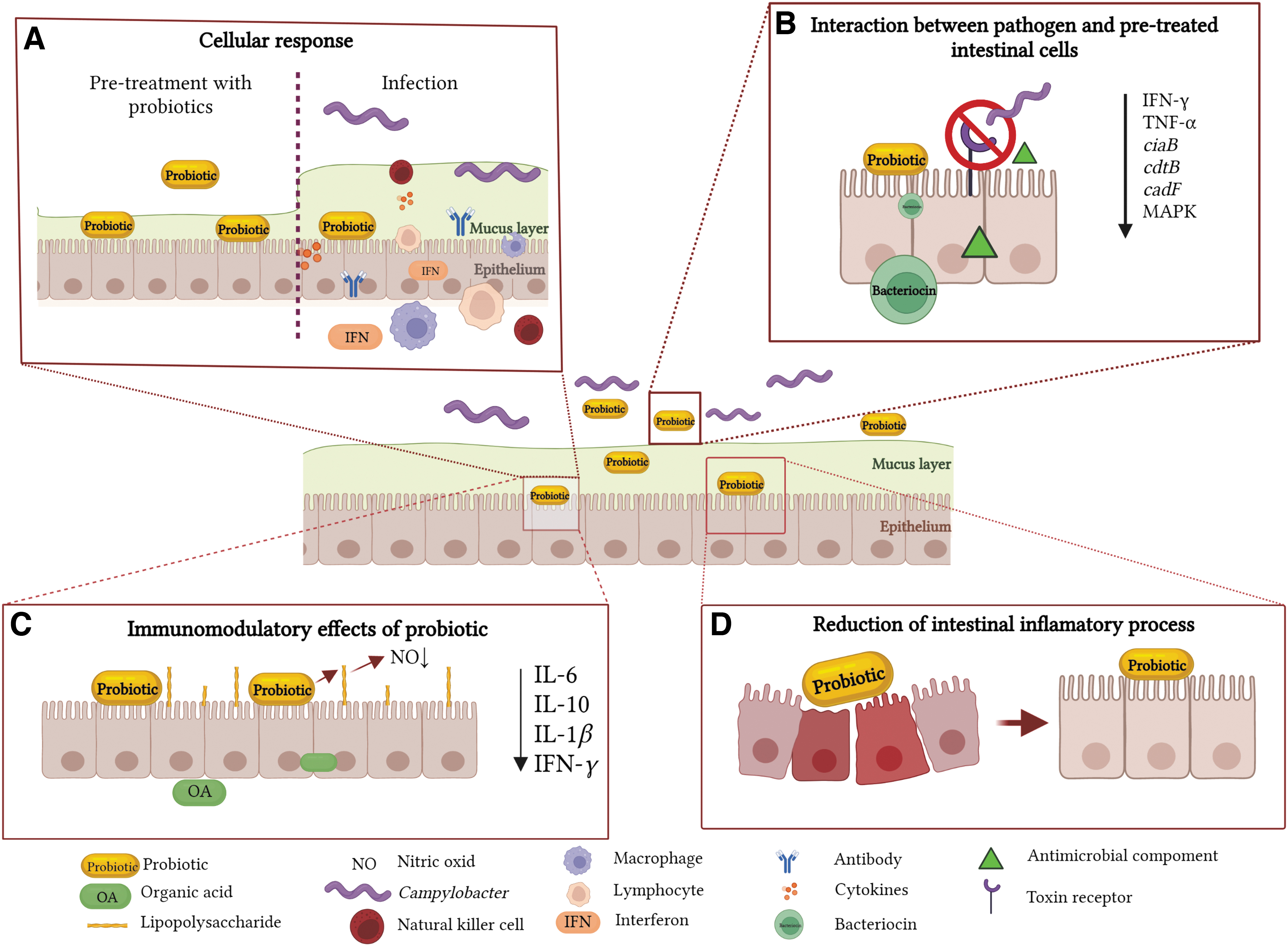
Probiotic mechanisms of action in alleviating Campylobacter infection.
Probiotics and Campylobacter Colonization in Poultry
Campylobacter colonization of the avian gastrointestinal tract (GIT) initiates in the small intestines, and continues to the cecum and cloaca without any clinical manifestations of the disease (Dec et al, 2018; Śmiałek et al, 2021). Nevertheless, several reports have indicated that challenged chickens could experience focal hepatic necrosis, signs of disseminated hemorrhagic gastroenteritis, and jejunal distention (Workman et al, 2005). The natural Campylobacter infection occurs through the fecal–oral route and is being established in the cecum at levels of ∼109 CFU/g of cecal content, with persistence during the entire bird lifetime (Awad et al, 2018; Dec et al, 2018). Once colonization is established, the entire flock becomes infected within a couple of days, with the infected birds showing excessive mucus production and enriched viscosity at an intestinal level (Awad et al, 2018).
It was also reported that higher levels of Campylobacter could be spotted in the crop region and with a lesser proportion in the broiler's gizzard (Smith and Berrang, 2006). In some cases, the Campylobacter spp. could also be detected in the internal organs (e.g., spleen, liver, bursa of Fabricius, and thymus), muscles, and blood samples during avian infection (Deng et al, 2020; Śmiałek et al, 2021). In response to infection, the avian host immunity triggers the production of proinflammatory cytokines, which modulate the GIT barrier function (Awad et al, 2018). The intestinal damage has a consequential impact on gut integrity by facilitating the transcellular/paracellular internalization of Campylobacter and progresses toward the underlying connective tissue, meanwhile eliciting the translocation of luminal pathogenic bacteria such as Salmonella, E. coli, and Clostridium to the internal organ compartments (Awad et al, 2018).
Nonetheless, can probiotics help in alleviating some of these effects? The biological mechanisms by which probiotics exhibit positive effects in poultry are not yet deciphered, but substantial advancements have been recently achieved in the case of Campylobacter spp. (Erega et al, 2021a; Kobierecka et al, 2017; Saint-Cyr et al, 2017). Details on the probiotic effects against Campylobacter spp. are in Table 1.
Anti-Campylobacter Probiotics in Poultry
BW, body weight; GIT, gastrointestinal tract; n.i., not identified.
The beneficial effects of probiotics are attributed to their ability to improve feed digestibility, nutrient bioavailability, and to enhance the immune system (Emami et al, 2020) and health leading to improved animal performance and carcass quality (Saint-Cyr et al, 2017; Yan and Polk, 2020). Most of the pathogen-associated inhibitory mechanisms of probiotics refer to nutrient depletion in a specific environmental niche, to the obstruction of the surface epithelial target receptors, usually used by pathogens during infection, and to their ability to synthesize natural antimicrobial molecules (Yan and Polk, 2020). The ability to induce potent immunomodulatory outcomes fortifies the avian-specific GIT immune mechanisms by producing interferon (IFN), antibodies, activated lymphocytes, macrophages, and natural killer cells to combat the diversity of infections and inflammatory processes upon Campylobacter infection (Śmiałek et al, 2021; Taha-Abdelaziz et al, 2019).
Clearly, probiotics have an immunomodulatory effect in poultry regardless of the pre- or posthatch administration as it has been shown that lactobacilli can modulate the immune response in newly hatched chickens (Alizadeh et al, 2021).
Organic acids were associated with antibacterial and immunomodulatory effects in poultry (Khan et al, 2022), also having an anti-Campylobacter effect (Sima et al, 2018). Butyrate-producing probiotic strains (e.g., Butyricicoccus pullicaecorum), improved feed efficiency and lowered Campylobacter levels in the ceca, by ∼1.5 logs, when fed to Ross 308 chicken broilers (Eeckhaut et al, 2016). Probiotic-derived butyrate also reduces the expression of proinflammatory cytokines (interleukin [IL]-6, IL-10, IL-1β, and IFN-γ) and hinders the production of nitric oxide in lipopolysaccharide (LPS)-stimulated cells (Trukhachev et al, 2021). Likewise, birds subjected to an oral gavage with a probiotic suspension of Lactobacillus salivarius SMXD51, 107 CFU at 14 and 35 d, showed significant reductions (0.82–2.81 logs) in Campylobacter levels (Saint-Cyr et al, 2017). Furthermore, L. salivarius SMXD51, an effective producer of salivaricin, was previously associated with induced immunomodulatory effects by boosting the IL-8 production and increased secretion of β-defensin 2 (Saint-Cyr et al, 2017).
Probiotic beneficial effects are not only limited to lowering Campylobacter levels in the GIT, they also extend to reducing environmental contamination (e.g., farms), and ultimately contributing to enhanced carcass hygienic indices (Smialek et al, 2018). The positive impact in the GIT was illustrated when C. jejuni-challenged Ross 308 chicks were fed with 1 × 109 CFU/kg Saccharomyces cerevisiae boulardii (CNCM I-1079). The significant improvement in villi height and crypt depth was accompanied by increased body weight gain and lower Campylobacter levels in the fecal and cecal samples (Massacci et al, 2019). Other similar broiler experiments concluded that Lactobacillus gallinarum PL53 (∼108 CFU) has the ability to lower C. jejuni presence in the cloaca and ceca, emphasizing the potential application as inhibitors of Campylobacter colonization at primary production (Khan, 2019).
Low numbers of campylobacters are able to also invade chicken intestinal cells (Fig. 2B) (Byrne et al, 2007), and thus, gaining knowledge on how probiotics can reduce colonization is beneficial and challenging. In summary, recent literature suggests that dietary supplementation of probiotics in poultry promotes an enforced intestinal barrier (Šikić Pogačar et al, 2020), enriches gut microbiota (Ty et al, 2022), lowers the systemic triglyceride and cholesterol levels (Vourakis et al, 2021), balances major blood biomarkers (e.g., albumin and glucose), and supports an enhanced absorption and digestion of the nutrients (Aponte et al, 2020). These benefits will contribute to disease prevention as well as to a reduced colonization of C. jejuni throughout infection (Dai et al, 2020).
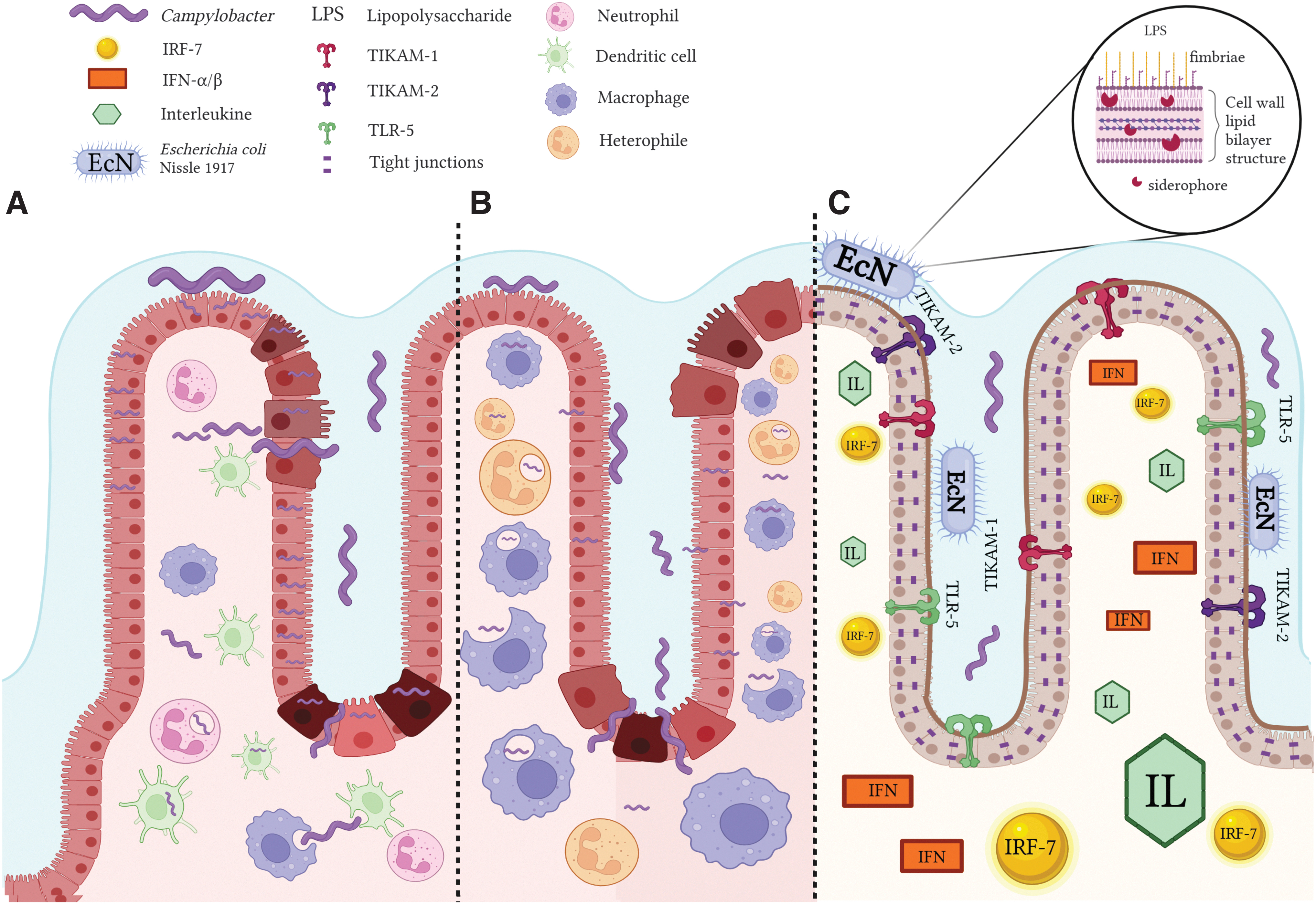
Campylobacter spp. infection of human and avian epithelium, and the proposed EcN mode of action against Campylobacter infection.
Probiotics and Preclinical Outcomes in Human Campylobacter Infections
In humans, oral and intestinal colonization (Lee et al, 2022) represents the first stages of C. jejuni pathogenesis (Fig. 2A) and is usually followed by distending toxin-mediated enteric infection and campylobacteriosis (Upadhyay et al, 2019). Several other findings have also established that the Campylobacter spp. isolated from the human oral cavity are indeed correlated with intestinal disease, and, in some instances, the detection levels are higher than in the fecal or intestinal biopsies (Xu et al, 2021). In addition, oral health studies have reported that Campylobacter abundance was slightly increased in the oral cavity of individuals encountering periodontal diseases and dental caries comparing with healthy patients (Lif Holgerson et al, 2015; Xu et al, 2021).
With the consumption of contaminated poultry meat being the main source of infection in humans, reducing Campylobacter levels in poultry through dietary probiotics could equally reduce human exposure to Campylobacter (Šikić Pogačar et al, 2020; Taha-Abdelaziz et al, 2019). Although previously thought that probiotic-based therapies are still marginalized in the absence of extensive clinical research, however, several recent findings have reported that probiotics are implicated in attenuating Campylobacter infection severity and are listed in Table 2. Probiotics can create a molecular weaponry necessary to outcompete and eliminate pathogens, colonize the environmental niche, and secrete various antimicrobial components. They can deploy enzymatic mechanisms responsible for modifying pathogen toxin receptors, hydrolysis of bacterial toxins, and inhibition of pathogen-induced illness (Trukhachev et al, 2021).
Anti-Campylobacter Probiotics, Evidence at Preclinical Level
APF, antiproliferative factor; n.i., not identified; NO, nitric oxide; PBS, phosphate-buffered saline; IL, interleukin; MCP-1, monocyte chemoattractant protein-1; TNF, tumor necrosis factor.
The competitive exclusion skill of probiotics could be enhanced with synbiotics, as it was recently exemplified that Bifidobacterium longum subsp. infantis ATCC 15697 and goat milk oligosaccharides prevented C. jejuni 81–176 invasion and adhesion of HT-29 cells (Quinn et al,
Recent preclinical findings reported that during C. jejuni-induced murine enterocolitis, the per-oral administration of a commercial probiotic (Aviguard) attenuated the apoptotic cell responses in C. jejuni 81–176-infected large intestine and improved the clinical outcomes (Heimesaat et al, 2021). The investigation of colonic biopsies, at 6 d postinfection, indicated statistically lower levels of IFN-γ and tumor necrosis factor alpha (TFN-α) after Aviguard treatment compared with the placebo groups. Meanwhile, the probiotic treatment averted C. jejuni-induced IFN-γ secretion from extraintestinal organs such as the liver, lungs, and kidneys. Lastly, an interesting fact from this pre-clinical trial indicated that the probiotic suspension Aviguard was successfully recovered from the intestines of treated subjects (Heimesaat et al, 2021) indicating its survival within the gut microbiome.
Recent data suggest that Caenorhabditis elegans activates the antibacterial peptide genes, including the upregulation of Daf-16 transcription factor and of MAPK signaling pathways, as an immunogenic effect against C. jejuni (Jin et al, 2021). Similarly, genetically engineered lactic acid-producing strains, Lactobacillus casei (LC+mcra), have the capacity to generate higher concentrations of conjugated linoleic acid, which showed a statistically high efficiency in C. jejuni growth decline (Tabashsum et al, 2018). Under coculturing conditions, the modified L. casei and its cell-free culture supernatants reduced the invasion and adherence of C. jejuni to HeLa and HD-11 cell lines. Some of C. jejuni virulence, including ciaB, cdtB, and cadF genes, was significantly downregulated in contrast to the flaB gene, which was significantly upregulated (Tabashsum et al, 2018).
In mice, using unconventional preclinical interventions, based on human fecal microbiota transplantation (FMT), per-oral treatment with probiotic strains provided evidence of the effective decrease in C. jejuni 81–176 levels and led to the mitigation of systemic inflammation and reduced pathogen-induced intestinal sequelae (Heimesaat et al, 2019a; Heimesaat et al, 2019b). Conclusively, it was suggested that novel probiotic formulations, as alternative strategies to FMT during severe GIT inflammations, might be efficient in lowering pathogen levels in vertebrates and farm animals and could even treat campylobacteriosis (Heimesaat et al, 2019b). Taken together, these studies indicate that clinical trials are the next obvious step in elucidating the in vivo effects of probiotics in preventing campylobacteriosis or in alleviating its secondary effects.
Types of Probiotics Used Against Campylobacter spp.
In the absence of virulence factors and with a boost in fitness patterns and survival elements, E. coli Nissle 1917 (EcN) (Fig. 1) represents the best example of a potent probiotic with a unique LPS that attributes immunogenicity without triggering immunotoxicity (Mawad et al, 2018). The biological safety of EcN was previously investigated, in both animal and human trials, with promising results in mitigating human ulcerative colitis, diarrhea, and other inflammatory-related diseases (Balta et al, 2021a; Garrido-Mesa et al, 2011; Mawad et al, 2018; Scaldaferri et al, 2016). Moreover, EcN was involved in the upregulation of intestinal antioxidant and anti-inflammatory reactionsm, which improved the antipathogen effect in poultry and mammalian cells and reduced diarrheal infection (Garrido-Mesa et al, 2011; Mawad et al, 2018). Alongside other recent in vitro findings, the EcN inhibitory activity against Campylobacter is further detailed in Figure 2 and Table 3.
In Vitro Evidence of Probiotics Efficient Against Campylobacter
n.i., not identified; NF-κB, nuclear factor kappa B; TLR, toll-like receptor.
As described in Figure 2C, the immunomodulatory effects of EcN, with impact on C. jejuni infection, include enabling of the nuclear factor kappa B (NF-κB) signaling pathways, the interferon regulatory factors (IRF-7), and the toll-like receptor (TLR) adaptor molecules (TICAM-1 and TICAM-2). In the HT-29 model of infection, the EcN modulatory effects extend to TLR-4 signaling, IL-12A/B, IL-1B, IL-17A, IFN-α/β, and the extracellular signal-regulated kinase pathway (ERK-1 and ERK-2), p38MAPK, antiapoptotic Akt signaling, and the c-Jun-NH2-kinase (JNK) (Helmy et al, 2021). Another interesting observation was that the expression of proinflammatory cytokines (IL-6/8/18, IL12-B, and TNF), NF-κB, mitogen-activated protein kinases (MAPK-1/3/8/14 and MAP2K3), TLR and TLR adaptor molecules (TLR-4/TLR-5, TICAM-1, and TICAM-2), NOD-1, apoptosis regulating factors (CASP-8 and RIPK-2), JUN, and MYD88-related (IRAK-3 and TRAF-6) genes was downregulated when cells were pretreated with EcN before C. jejuni infection (Helmy et al, 2021).
These mechanistic insights are linked to prestimulation of the epithelial immune protection systems to facilitate counteraction of the proinflammatory response caused by C. jejuni infection. Overall, these results improved our knowledge on how probiotic agents, such as EcN, interrupt C. jejuni infection and gastroenteritis, prevent disruption of the epithelial barrier, and inhibit the host's proinflammatory responses (Helmy et al, 2017; Helmy et al, 2021; Mawad et al, 2018).
Other means by which probiotics exclude bacterial pathogens include the production of bacteriocins, efficient antimicrobial peptides (Ahsan et al, 2022). One example is curvaticin, a bacteriocin synthesized by Lactobacillus curvatus DN317, which expressed a remarkable bacteriostatic activity against the chicken isolate C. jejuni ATCC 33560 (Zommiti et al, 2016). Growth inhibition was achieved when 50, 100, and 150 AU/mL of L. curvatus supernatant was introduced to the growing culture of C. jejuni ATCC 33560 and decreased the viable bacterial counts by ≈23.8%, 45.5%, and 61.3%, respectively (Zommiti et al, 2016).
Enterocins are another example of bacteriocins, derived from Enterococci spp., which in combination with herbal extracts can be applied as a new ecological approach to Campylobacter reduction in livestock commodities, therefore improving human health safety (Ščerbová et al, 2022). Lactobacillus reuteri could also produce inhibitory compounds, under anaerobic fermentation conditions, such as reuterin, which has broad-spectrum antimicrobial activity against various foodborne pathogens (Asare et al, 2020). A chicken-synthesized reuterin, produced by L. reuteri PTA5_F13, demonstrated a more significant inhibition of C. jejuni N16-1419 when 28 mM glycerol, added in the in vitro experimental conditions, was used as a reuterin precursor (Asare et al, 2020).
Furthermore, the study results showed that during coculturing of L. reuteri PTA5_F13 with 28 mM glycerol, C. jejuni counts drastically dropped from 7.3 logs CFU/mL to just above the detectable limits (1 log CFU/mL). The study concluded that such efficient anti-Campylobacter outcomes might be valuable in slaughterhouse poultry equipment decontamination processes.
Biosurfactants, another category of antiadhesive molecules (e.g., fengycin, iturin, and surfactin, produced by Bacillus subtilis) (Erega et al, 2021a), have the ability to significantly reduce C. jejuni NCTC11168 (by ≈4.2 log10) growth, and biofilm formation and adhesion to abiotic polystyrene surfaces, through a mechanism dependent on diffusible factors (e.g., nonribosomal/polyketide bacillaene) (Erega et al, 2021a). Promising anti-Campylobacter effects of B. subtilis PS-216 were also outlined in several other studies (Erega et al, 2021a; Šimunović et al, 2022). First, B. subtilis PS-216 demonstrated significant antagonism against C. jejuni through growth inhibitory effects (4.2 log10) and reduced biofilm formation and adherence (2.4 log10) to abiotic surfaces. These effects were attributed to the production of the antimicrobial compound nonribosomal/polyketide bacillaene (Erega et al, 2021a). The authors specified that wild-type B. subtilis, carrying the sfp gene encoding for phosphopantetheinyl transferase, is linked to the production of antimicrobial molecules and to the expression machinery of bacillaene.
Second, it was reported that under coculture conditions, B. subtilis reduced C. jejuni growth by 3.87–4.07 logs, and dropped below the detection limits after 48 h (Šimunović et al, 2022). Given that all these effects were observed at 42°C, which is the optimal body temperature in poultry, B. subtilis PS-216 becomes a strong probiotic candidate for Campylobacter reduction in poultry (Šimunović et al, 2022).
Efficient probiotics will also have to be able to overcome and survive the GIT environment (Grispoldi et al, 2020). Probiotic candidates in poultry, such as Lactobacillus spp., have shown adhesion and invasion inhibitory properties, against C. jejuni K49/4, and persistence in the chicken small-intestine cells and pig enterocytes (Šikić Pogačar et al, 2020). Coculturing of C. jejuni with Lactiplantibacillus plantarum PCS20, PCS22, PCS25, and PCK9 considerably declined the number of campylobacters that adhered and invaded the polarized intestinal epithelial cells. Compared with other strains, Lactobacillus rhamnosus LGG and L. plantarum PCS25 statistically reduced the invasion of C. jejuni below the limit of detection (Šikić Pogačar et al, 2020). L. plantarum was also efficient in broilers when part of a microencapsulated symbiotic was combined with fructooligosaccharides, having a positive impact on growth performance, immune and antioxidant parameters, and the digestibility of calcium and phosphorus (Song et al, 2022).
In vivo has been confirmed that lactic acid bacteria can reduce C. jejuni NCTC 11168 pathogenicity, first in a C. elegans nematode model followed by subsequent extrapolation into a mouse and chicken model (Jin et al, 2021). When nematodes were infected with C. jejuni and administered Lactobacillus spp. (13-7, N9, and Z5 strains), a significant increase in the expression of MAPK signaling pathway genes (pmk-1, nsy-1, sek-1, and tir-1), antioxidant genes (skn-1 with bar-1), defence immune genes (daf-16, age-1, and dbl-1), and antibacterial peptide genes (spp-1, clec-85, abf-2, clec-60, and lys-7) was observed (Jin et al, 2021). Strain Z5 also demonstrated increased inhibitory activity of C. jejuni colonization in the chicken ceca (below 104 CFU/g), in contrast to the untreated control and the infected group where counts were above 108 CFU/g.
Less efficient, but significant, decreasing trends were also reported for the 13-7 and N9 isolates, which lowered the ceca counts by ∼103 CFU/g. The combined effects of all the above examples strengthen the general view that the C. jejuni-antagonistic effects of probiotics are articulated by the overexpression of the immune-associated genes, as proven and previously described in nematode, mice, and chicken models of infection.
Conclusion
The findings of this review describe the latest antivirulence, broad antibacterial, and immunomodulatory effects of various bacterial probiotic species against emerging Campylobacter spp. infections. The anti-Campylobacter probiotics will potentially possess other desired features (e.g., acidic/bile resilience, bile salt hydrolase capacity, efficient colonization of the gut properties) indicating wider health beneficial effects and potent immunomodulatory effects. More efforts should be made to further elucidate and decipher the pathogenic mechanisms of enteropathogenic bacteria such as Campylobacter, especially in poultry, to accelerate the validation and accreditation of new probiotic strains.
Footnotes
Acknowledgment
We would like to acknowledge Environtech, Dublin, Ireland, for funding the PhD program of Eugenia Butucel.
Authors' Contributions
All authors contributed equally to the preparation of this article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.