
Abstract
Acinetobacter baumannii is a highly antibiotic-resistant bacterial pathogen known to cause severe life-threatening infections, including pneumonia, meningitis, and sepsis. Recent emergence of this bacterium as a serious nosocomial pathogen has led to categorization of A. baumannii as a “high-priority” pathogen by the World Health Organization (WHO), for which research efforts are urgently required to develop therapeutic interventions. Some of the properties that make A. baumannii a serious pathogen include its capacity to tolerate high levels of stress and enhanced expression of efflux pumps that enable high degrees of antibiotic resistance. Virulence mechanisms employed by A. baumannii to establish successful infection and host responses elicited against A. baumannii to counter the infection are discussed in detail in this article.
Introduction
A
Another major attribute leading to widespread persistence of A. baumannii in nosocomial environments is its ability to resist severe conditions that are detrimental for other pathogens, thus conferring A. baumannii with a survival advantage. It can endure disinfection (Hassan et al., 2013) and extended periods of desiccation (Antunes et al., 2011). Moreover, A. baumannii is known to form robust biofilms both within the host (Thompson et al., 2014) and also on abiotic surfaces such as hospital devices, which can further contribute to infection (Greene et al., 2016b). Bacterial colonization in these biofilms increases extracellular stress tolerance and thus enhances bacterial persistence (Greene et al., 2016a). Association of A. baumannii with hospital devices and equipment has become a major contributor to infection and has been linked to ventilator-associated pneumonia, catheter-associated urinary infections etc. (Wong et al., 2017). Over the last few years many studies have been directed toward understanding the pathobiology of A. baumannii. A number of virulence mechanisms and host responses to infection have been uncovered, thus advancing our understanding of A. baumannii infection as detailed in the following sections.
Major Virulence Strategies Employed by A. baumannii
Multiple virulence factors and mechanisms have been described that A. baumannii employs to induce host damage. One of the major classes of proteins that has evolved from several studies has been porins. Porins are outer membrane proteins that modulate cellular permeability. OmpA is a major porin protein present in the outer membrane of A. baumannii. It serves multiple functions, including adhesion to host epithelial cells, resistance against complement killing, and biofilm formation (Schweppe et al., 2015). OmpA protein is released through outer membrane vesicles (OMVs) of A. baumannii and targets mitochondria in host cells, leading to mitochondrial fragmentation and apoptosis through the release of proapoptotic molecule cytochrome C (Choi et al., 2005; Tiku et al., 2020, 2021). OmpA further increases reactive oxygen species (ROS) levels and decreases mitochondrial polarization and ATP levels (Fig. 1). Concordantly A. baumannii OmpA mutant is significantly attenuated in its ability to induce host cell death (Choi et al., 2005; Tiku et al., 2021).
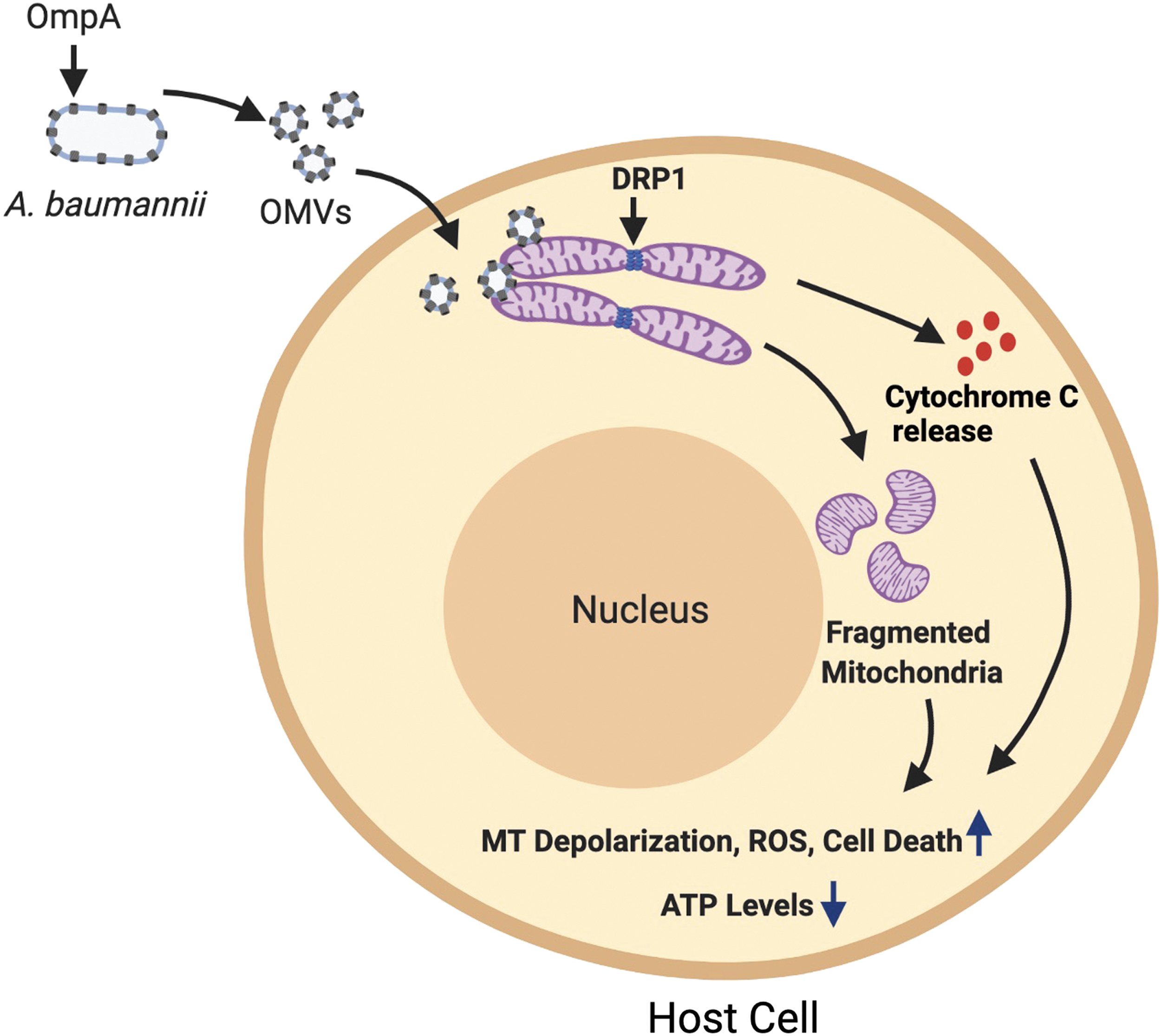
Acinetobacter baumannii secretes OMVs containing OmpA that induce mitochondrial damage. Cytotoxic OMVs containing OmpA are secreted by A. baumannii, which deliver OmpA to mitochondria. OmpA induces fragmentation of the mitochondrial network driven by the host protein DRP1, which is the master regulator of mitochondrial fission. OmpA-mediated mitochondrial fragmentation is accompanied with enhanced ROS levels, decreased ATP levels, and increased mitochondrial depolarization. OmpA-mediated damage to host cell mitochondria also leads to the release of the mitochondrial protein cytochrome C. Release of cytochrome C induces apoptotic cell death in host cells. Figure created with
OmpA also helps in adherence and subsequent invasion of epithelial cells by interacting with extracellular matrix proteins such as fibronectin (Smani et al., 2012). OmpA binds to factor H in human serum thereby assisting A. baumannii to escape complement-mediated killing (Kim et al., 2009). Moreover, OmpA loss lowers the minimal inhibitory concentrations or MICs of multiple antibiotics, including chloramphenicol and aztreonam, which suggests that OmpA plays a key role in expelling antibiotics out of bacterial cells thereby leading to multidrug-resistant (MDR) phenotype of A. baumannii (Smani et al., 2014). Another important porin protein linked with A. baumannii pathogenesis is the 33- to 36-kDa Omp called Omp33-36. This protein is also important for host cell adherence and its loss leads to attenuated host cell adhesion and lowered cytotoxicity in human lung epithelial cells and macrophages.
Omp33-36 loss reduces lethality, bacterial dissemination, and bacterial load in the lungs and spleen of mice (Smani et al., 2013). Omp33-36 induces apoptosis by activating caspases and perturbs autophagy by causing an accumulation of the selective autophagy receptor p62/SQSTM1 and the autophagosomal membrane protein LC3B-II in immune and connective tissue cells. Modulation of autophagy enables A. baumannii to persist intracellularly and reside in autophagosomes (Rumbo et al., 2014). Yet another important outer membrane porin protein Omp22 has been suggested to be an efficient target for vaccine development against A. baumannii infections. Immunization of mice with recombinant Omp22 increased survival rates of mice, reduced bacterial load in different organs, and lowered the levels of inflammatory cytokines and chemokines in the serum (Huang et al., 2016).
The exact mechanism of how Omp22 functions in the pathogenesis of A. baumannii infections still remains elusive; nevertheless, Omp22 serving as a potential vaccine target holds promise in the race for developing therapeutics against A. baumannii. Additionally, OMVs, which are spherical vesicles secreted by the outer membrane of many Gram-negative bacteria, can also serve as a potent strategy by A. baumannii to deliver toxins. These toxins include OmpA, certain cytotoxic proteases and phospholipases (Kwon et al., 2009; Tiku et al., 2021).
Another important determinant of A. baumannii pathogenicity is the bacterial capsule. Mutants deficient in capsular exopolysaccharides are more susceptible to peptide antibiotics. Moreover, mutations that affect sugar precursors that lead to the formation of the capsule and lipopolysaccharide (LPS) sensitize A. baumannii to multiple antibiotics (Geisinger and Isberg, 2015). A transposon mutagenesis screen to identify genes required for proper growth of A. baumannii in inflammatory human ascites fluid revealed ptk and epsA as essential genes. Both ptk and epsA play important roles in capsule formation and assembly, and mutations in these genes lead to compromised capsule formation and growth defects of bacteria in human serum (Russo et al., 2010). PglC is required for the formation of O-pentasaccharide found on the glycoproteins. Loss of PglC in A. baumannii prevented proper synthesis of the capsule, consequently leading to perturbed biofilm formation, and reduced virulence in mice (Lees-Miller et al., 2013).
In addition to the components of the capsule, LPS, an immunoreactive molecule present in the outer membrane of Gram-negative bacteria, also leads to virulence of A. baumannii. LpsB glycotransferase is an essential factor required for the production of LPS in A. baumannii, and a mutation in this gene renders the bacterium defective in LPS, which causes decreased resistance to human serum and attenuated survival in rats (Luke et al., 2010). A number of other studies have also highlighted the role of LPS in survival and pathogenicity of A. baumannii (Lin et al., 2012; McQueary et al., 2012).
Protein secretion systems form another potential virulent strategy used by A. baumannii. The bacterium possesses Type I, II, IV, V, and VI secretion systems, which have not been studied in great detail so far. Recently it was shown that A. baumannii uses its type 2 secretion system (T2SS) to secrete effector lipases LipA and LipH as well as the protease CpaA (Harding et al., 2016). Another study from the same group reported the virulence potential of CpaA in both invertebrate and mouse model of pneumonia (Kinsella et al., 2017). Furthermore A. baumannii utilizes T6SS for bacterial competition (Weber et al., 2015), although its role in cytotoxicity is unknown yet.
Host Responses Elicited by A. baumannii
Multiple studies have uncovered specific host responses against A. baumannii infections. At the initial recognition of the bacteria, epithelial cells secrete antimicrobial peptides like human β-defensins that restrict A. baumannii growth, forming the first line of defense against the pathogen (Feng et al., 2014). As with most infections, pattern recognition receptors (PRRs) at the cell membrane recognize pathogen-specific molecular signatures known as the pathogen-associated molecular patterns. A key PRR, membrane-bound toll-like receptor 4 (TLR4) has been implicated to play an important role in mounting an immune response and assisting bacterial clearance in A. baumannii infections in mice (Knapp et al., 2006) (Fig. 2). Another PRR TLR2, which is important for recognizing peptidoglycan of the bacterial membrane, has a minimal role in host defense in curbing A. baumannii infections (Knapp et al., 2006; Kim et al., 2014).
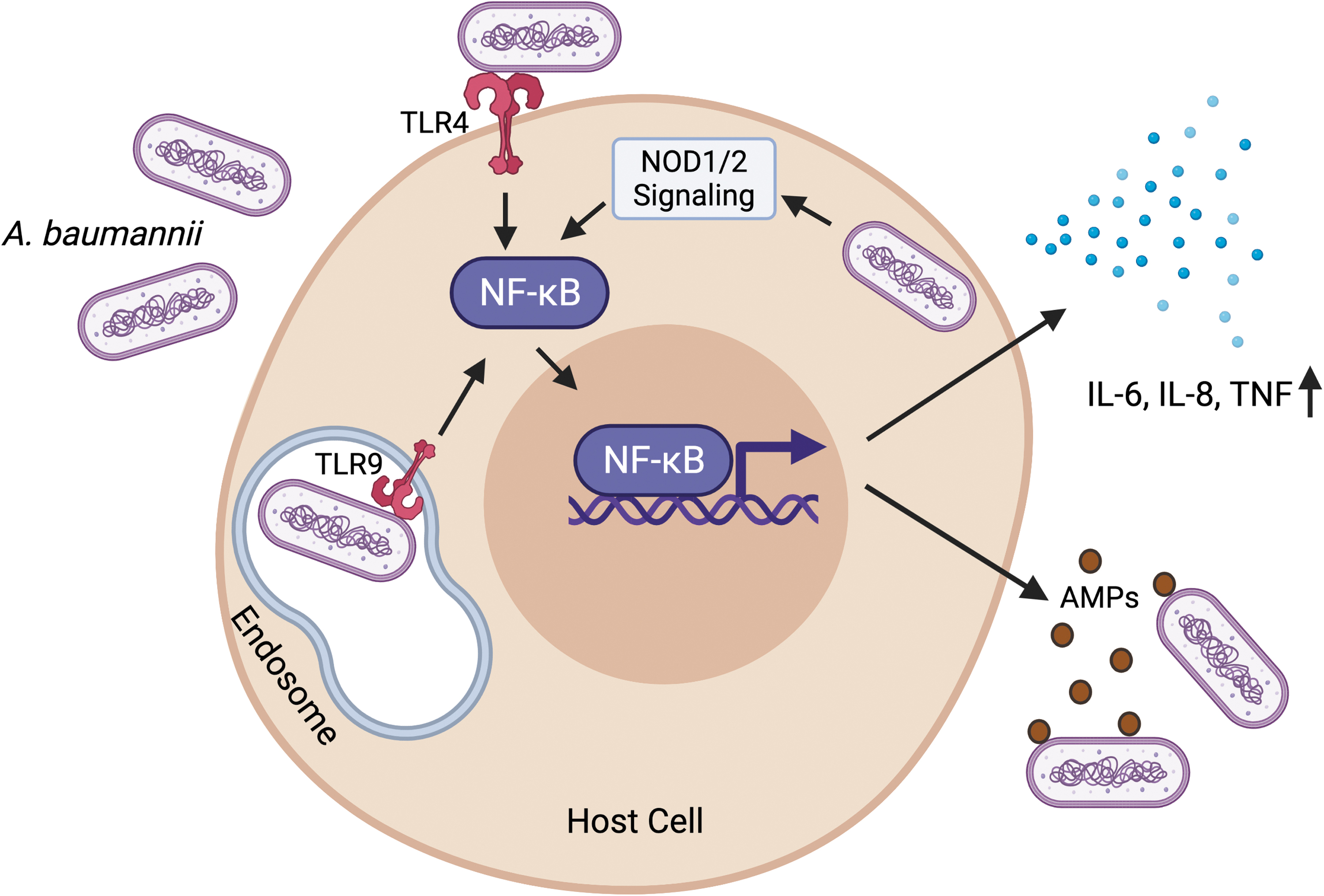
Host responses against A. baumannii. Infection with A. baumannii activates the plasma membrane-bound TLR4 and the endosome-bound TLR9. Activated TLR signaling leads to the activation and nuclear translocation of the transcription factor NF-κB. Intracellular A. baumannii also contributes to NF-κB activation through the intracellular PRRs—NOD1 and NOD2. Activated NF-κB drives the expression of proinflammatory cytokines, including TNF, and IL-6 and IL-8. NF-κB also stimulates the expression of AMPs like human β-defensins, which restrict A. baumannii growth. Figure created with
Traditionally A. baumannii has been considered to be an extracellular pathogen, but recent evidence has shown that the bacterium can persist intracellularly as well (Sycz et al., 2021). The pathogen invades epithelial cells and internalizes in a zipper-like mechanism associated with microtubule and microfilament-dependent uptake (Choi et al., 2008). Internalized bacteria reside in membrane-bound vacuoles in the cytoplasm (Choi et al., 2008), thereby imposing the need of detection by intracellular PRRs. Interestingly, TLR9, which is an endosomal PRR that detects viral/bacterial DNA and helps in mounting a subsequent immune response, is required for bacterial clearance and induction of proinflammatory cytokines and chemokines in a murine model of A. baumannii pneumonia (Noto et al., 2015) (Fig. 2). Additionally, NOD1 and NOD2, which are also intracellular PRRs, and their serine/threonine adaptor protein kinase RIP2, which is induced upon A. baumannii infection, play central roles in sensing and subsequent clearance of A. baumannii in human airway epithelial cells (Fig. 2).
Loss of NOD1, NOD2, and RIP2 increases intracellular invasion and augments survival of A. baumannii in lung epithelial cells (Bist et al., 2014). Finally sensing of the pathogen by PRRs leads to an activation of extensive downstream signaling, which involves concerted functions of NF-κB and the kinase cascade relayed by the mitogen-activated protein kinase pathway, ultimately culminating in the production of inflammatory molecules, including tumor necrosis factor, and interleukins (IL)-6 and IL-8 (March et al., 2010; Bist et al., 2014) (Fig. 2).
Cellular response of the host against A. baumannii entails recruitment of specialized immune cells to the site of infection. The migration of these cells is directed by cytokines and chemokines. Neutrophils and macrophages are the first cells to arrive (Harding et al., 2017). Neutrophils are the major players in keeping A. baumannii infection under check by phagocytosing and clearing the bacteria from the infection site (Bhuiyan et al., 2016). In accord with this, mice deficient in neutrophils exhibit acute lethality and a significant increase in bacterial burden upon A. baumannii infection (Faassen et al., 2007; Breslow et al., 2011). At the molecular level, neutrophils kill A. baumannii by using the NADPH phagocyte oxidase system, which leads to the production of reactive oxygen species (ROS). NADPH-deficient neutrophils are unable to contain bacterial replication and subsequent dissemination (Qiu et al., 2009). Besides neutrophils, macrophages also help in the clearance of A. baumannii from the infection sites (Qiu et al., 2012), but the role of macrophages is secondary to neutrophils and is not vastly explored.
Future Perspective
Extreme MDR phenotypes associated with A. baumannii have invoked great concern and have paved the way for studies aimed to understand the pathobiology of this bacterium. Future efforts should be focused on trying multiple strategies to combat A. baumannii infection. Clinical interventions like immunotherapy hold great promise for treatments against these infections. Since neutrophils are the major phagocytic killer cells in A. baumannii infections, methods of directing neutrophils toward the sites of infection using chemokines and cytokines maybe a viable therapeutic approach to consider. Additionally, other strategies like phage therapy, small molecules targeting bacterial metabolic pathways, and developing novel vaccine targets could prove beneficial (García-Quintanilla et al., 2013). Since A. baumannii is resistant to most of the antibiotics currently available, clinicians opt for colistin as the last resort. However, recent cases have reported colistin resistance in A. baumannii as well. Therefore, it might be necessary to use combination therapies involving different antibiotics.
Finally, since A. baumannii possesses multiple protein secretion systems, it is of great interest to understand in detail their functions in regulating the virulence mechanisms of the pathogen. Many bacteria use their protein secretion systems to deliver potent virulence factors and toxins to spread infection. Therefore, it is vital to get a deeper understanding of the functioning of the different secretion systems in A. baumannii to develop successful therapeutics against this pathogen.
Footnotes
Disclosure Statement
Varnesh Tiku is an employee of Vir Biotechnology, Inc. and may hold shares of Vir Biotechnology, Inc.
Funding Information
No funding was received for this article.