
Abstract
Abstract
Background and Objectives:
The cannabinoid receptor 2 (CB2) was previously implicated in brain functions, including complex behaviors. Here, we assessed the role of CB2 in selected swimming behaviors in zebrafish larvae and developed an in vivo upscalable whole-organism approach for CB2 ligand screening.
Experimental Approach:
Using CRISPR-Cas9 technology, we generated a novel null allele (cnr2upr1) and a stable homozygote-viable loss-of-function (CB2-KO) line. We measured in untreated wild-type and cnr2upr1/upr1 larvae, photo-dependent (swimming) responses (PDR) and center occupancy (CO) to establish quantifiable anxiety-like parameters. Next, we measured PDR alteration and CO variation while exposing wild-type and mutant animals to an anxiolytic drug (valproic acid [VPA]) or to an anxiogenic drug (pentylenetetrazol [PTZ]). Finally, we treated wild-type and mutant larvae with two CB2-specific agonists (JWH-133 and HU-308) and two CB2-specific antagonists, inverse agonists (AM-630 and SR-144528).
Results:
Untreated CB2-KO showed a different PDR than wild-type larvae as well as a decreased CO. VPA treatments diminished swimming activity in all animals but to a lesser extend in mutants. CO was strongly diminished and even more in mutants. PTZ-induced inverted PDR was significantly stronger in light and weaker in dark periods and the CO lower in PTZ-treated mutants. Finally, two of four tested CB2 ligands had a detectable activity in the assay.
Conclusions:
We showed that larvae lacking CB2 behave differently in complex behaviors that can be assimilated to anxiety-like behaviors. Mutant larvae responded differently to VPA and PTZ treatments, providing in vivo evidence of CB2 modulating complex behaviors. We also established an upscalable combined genetic/behavioral approach in a whole organism that could be further developed for high-throughput drug discovery platforms.
Introduction
The endocannabinoid (eCB) system is a key modulator of excitatory and inhibitory neuronal activity 1 and its dysregulation has been linked to several psychiatric disorders.2–5 The two cannabinoid receptors (CB1 and CB2) belong to the G protein-coupled receptor family. They are both activated by endogenous ligands (eCBs)6,7 and exogenous compounds such as Δ 9 -THC, the main psychoactive component in cannabis. 8 CB1 is highly expressed in the CNS and implicated in numerous neurological diseases (for review, see Marco et al., 3 Kendall and Yudowski, 5 Bilkei-Gorzo, 9 Di Marzo et al., 10 and Pavlopoulos et al. 11 ). By comparison, CB2 expression was initially described in the immune system but more recently also in discreet brain regions where its role is still poorly understood.
A genome-wide association study showed association between specific SNPs in the CNR2 gene encoding CB2 and schizophrenia. 12 Several lines of evidence suggest a role for CB2 in complex and specific behaviors in adult rodents.13–15 CB2-KO mice displayed schizophrenia-related behaviors 16 altered cognitive function,17,18 modified cocaine-reward behaviors, 19 as well as increased aggressiveness. 20 CB2 overexpression showed reduced anxiety-like behaviors 21 and resistance to depression, 22 whereas temporary blockage of CB2 expression exhibited reduced aversion to open space. 15 Conditional CB2-KO demonstrated that CB2 can regulate synaptic transmission in hippocampal pyramidal cells and modulate gamma oscillation. 23 Modulation of CB2 expression in the hippocampus showed a regulatory role in fear and working memory. 18 Suppression of CB2 expression in dopamine neurons inhibited psychomotor behaviors, altered anxiety, and depression measurements, as well as alcohol preferences. 24 So far, few developmental studies were performed for CB225,26 and none was exploring complex behaviors during this critical period.
Zebrafish is a powerful, genetic, and developmental model, which also provides the unique feature of upscalability, allowing high-throughput applicable to pharmacological screens (for review, see Rennekamp and Peterson 27 ). Using CRISPR-Cas9, we created a novel null allele (cnr2upr1) and established a stable loss-of-function (CB2-KO) zebrafish mutant line. 28 Homozygote larvae were viable without an overt phenotype and were raised into fertile and healthy adults over several generations, which were all completely lacking CB2. Next, we tested swimming behaviors in 6-day postfertilization (dpf) wild-type and homozygote larvae monitoring photo-dependent responses (PDR)29,30 and measuring the center occupancy (CO) of wells providing an inverse measure of center avoidance. We found that CB2-KO were swimming significantly less in light and significantly more in dark periods with a decreased CO, when compared with wild-type larvae. When adding a broad-spectrum anxiolytic drug (valproic acid [VPA])30,31 just prior recording, we found that swimming activity and CO were strongly reduced in all animals in a similar manner, but swimming was slightly less and CO more diminished in mutants. When adding a classical anxiogenic drug, pentylenetetrazol (PTZ), a well-characterized GABAA inhibitor in many animal models,32–36 we found that larvae lacking CB2 presented an increase in swimming activity and a decrease in CO when compared with wild type. Taken together, we provide in vivo evidence for CB2 modulating complex behaviors in zebrafish larvae.
Finally, to test the potential of our approach for CB2 ligand screening, we treated wild-type and CB2-KO larvae with two CB2-specific agonists (JWH-133 and HU-308) and two CB2-specific antagonists (AM-630 and SR-144528). 37 We found that two of four treatments elicited detectable PDR alterations, possibly CB2 mediated for the most part. Thus, we described a novel, upscalable behavioral approach for drug screening in a whole organism, providing a complementary alternative to current methods.
Materials and Methods
Zebrafish care and husbandry
We used TAB5 or NHGRI wild-type animals that we raised and maintained in our fish room following standard procedures and IACUC protocol (#A880216).
CRISPR-Cas9 generation of the cnr2upr1 allele
We designed a CRISPR-Cas9 guide targeting 5′-ATGGCGTTTACGGGCTCTGT-3′ in cnr2 (ENSDARG00000039970).28,38 Progeny were screened for insertion/deletion (INDEL) by polymerase chain reaction (PCR) amplification and sequencing with the following primers (F: 5′-GACCACACAAGAGCAGAAAGC-3′; R: 5′-GACGATCCAACCAGGTTTTG-3′), and by fluorescent-based PCR fragment analysis performed with fluorescent primers (F: 5′-CGCCCATCGTACCTGTTTAT-3′; R: 5′-TTGGCTCTAGTGCGTGTCAG-3′ 39 ).
CB2 mRNA and protein expression
Total RNA was prepared from genotyped larvae and the subregion of interest in the cnr2 mRNA transcribed using retrotranscriptase (Sigma) and the following primers (F: 5′-CAGCTGCCACGTGATATAAGTA-3′; R: 5′-ATGCCAGCATTTCTCCCCTC-3′), and subsequently sequenced with the same primers.
For Western blots, adult wild-type and cnr2upr1/upr1 brains were dissected and digested in radioimmunoprecipitation assay buffer (Thermo Fisher Scientific #89900), supplemented with protease and phosphatase inhibitor cocktail (100×) (Thermo Fisher Scientific #78440). A dilution of 1:500 was used for the anti-CB2 antibody (Ab; Cayman Chemicals #101550) and 1:1000 for the anti-alpha-tubulin Ab (Developmental Studies Hybridoma Bank #12G10).
The behavioral swimming assay: the PDR
Five dpf larvae were loaded into 48-well plates (CELLSTAR®) 24 h prior recording, for animals to adapt to the new environment. A single larva was placed in each well in 450 μL system water (SW). Next day, wells were topped off to 500 μL with SW and the desired concentration of the drug at study or with SW, and plates were immediately placed in the recording device (Zebrabox, Viewpoint, France). After a 30-min adaptation/incubation in the dark, animals were subjected to 10 min of light (L) at maximum intensity (=385 Lux) followed by 10 min of dark (D) in four successive cycles. All results were binned into 1-min intervals.
Center occupancy
We virtually defined an inner central diameter (0.5 cm) within the whole well (1.0 cm) and recorded inner and total traveled distances. The percentage of CO was calculated as follows: CO=inner traveled distance/total traveled distance×100.
Drug preparation
Stock solutions were prepared in SW at 20 mM for VPA (Sigma #P4543) and 75 mM for PTZ (Sigma #P6500), 30 or in dimethyl sulfoxide at 10 mM for all CB2 ligands (Tocris bioscience: JWH-133 #1343, HU-308 #3088, AM-630 #1120, and SR-144528 #5039). Further dilutions were done in SW.
Toxicity/survival assay
CB2 ligands Kis are in the nM range. However, to overcome diffusion problem and permeability issues, we expected to use them in the μM range. To exclude aberrant swimming patterns induced by overexposure compromising health and survival, we treated healthy 5 and 6 dpf larvae with CB2 ligands at 1, 10, and 50 μM (n=10/ligand/concentration) for 24 h and assessed the following criteria: overall larval morphology, spontaneous swimming, and responses to sound and mechanical stimuli. Concentrations resulting in abnormal morphology or responses were considered not well-tolerated and excluded from further experiments.
Statistical analysis
We analyzed averaged total traveled distances per larva (with a minimum of triplicate experiments and 24 animals/treatment) in GraphPad Prism (v.7). All results were binned into 1-min intervals and error bars represent mean±standard error of the mean. Statistical differences between direct comparisons were calculated using multiple t-tests controlling the effect of the correlation among the number of fixed repeated measures. We performed two-way analysis of variance in graphs when two or more groups were compared simultaneously. Differences with p<0.05 were considered significant (*).
Results
Generation of a CB2 loss-of-function stable mutant line (null allele: cnr2upr1)
In zebrafish, a single cnr2 gene with two transcripts (Fig. 1A) resulting in identical translated exons (solid red blocks) encodes a 383 amino acid (aa) CB2 protein, which is slightly longer than the human homologue (360 aa). To generate loss-of-function alleles, we designed guide RNAs targeting the 5′ end of the second translated exon (Fig. 1A, T in light blue).
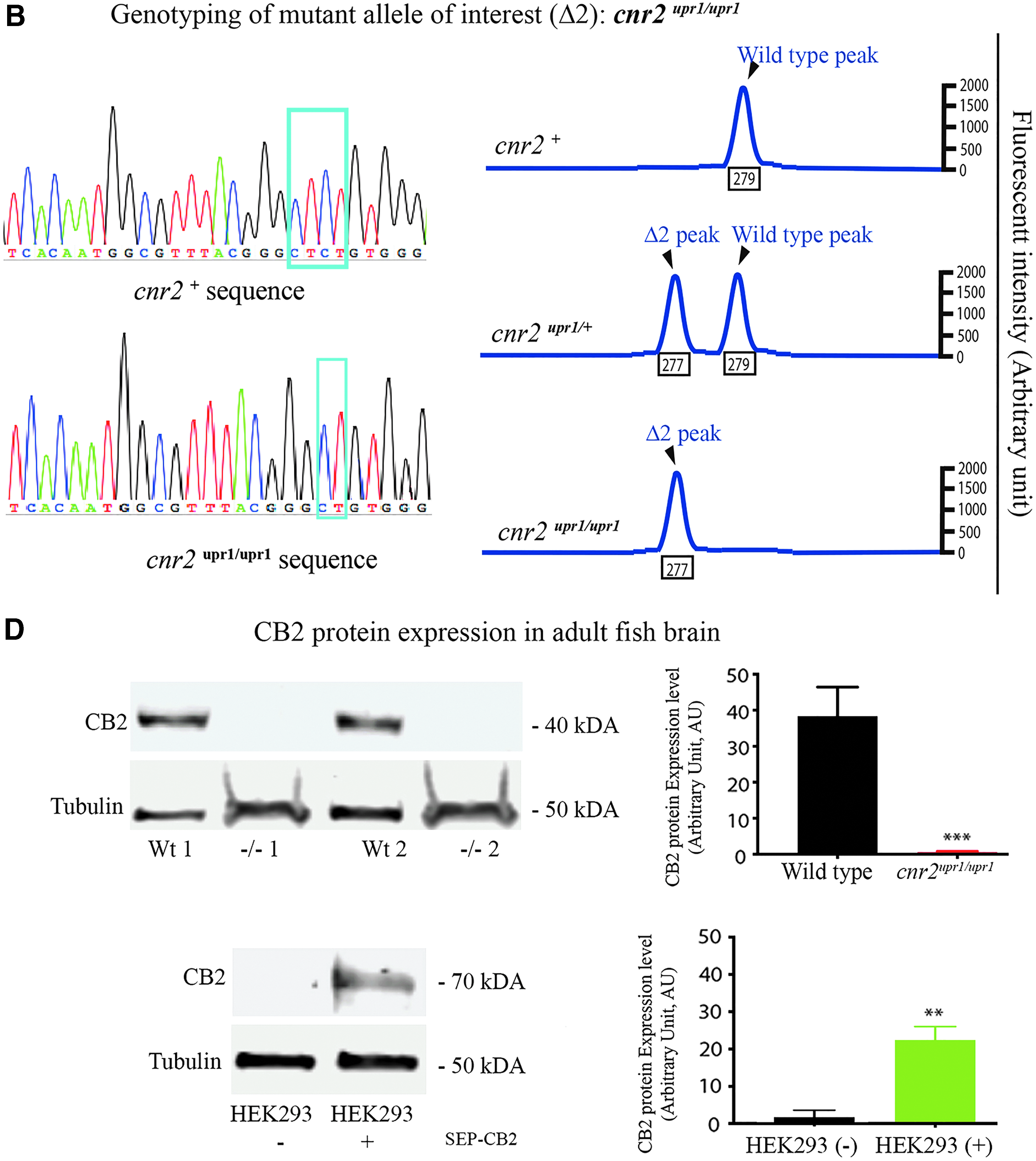
Generation of a CB2 loss-of-function stable mutant line (null allele: cnr2upr1).
We outcrossed adult founders (F0) and genotyped the offspring (F1) for germ line transmission of INDEL in the target site. We identified a two-nucleotide deletion (Δ2: CT) just 3 base pair upstream of the protospacer adjacent motif site (purple square in bottom of Fig. 1A) introducing a translation frameshift, which would predictably create an early stop codon in aa position 159. The resulting truncated protein, if not degraded, would have only two transmembrane domains, thus very likely a loss-of-function mutation. We grew this allele (cnr2upr1) to homozygosity and genotyped animals by classical sequencing (Fig. 1B, left panels) or by fluorescent PCR 40 (Fig. 1B, right panels). We closely monitored heterozygote or homozygote larvae development and morphology between 2- and 9-dpf and found no obvious phenotype. Likewise, adult genotyped cnr2upr1/upr1 animals were healthy, fertile, and inbred into a stable F3 generation, from which we obtained cnr2upr1/upr1 larvae used in all the behavioral studies.
To confirm that we had generated a loss of function, we analyzed the cnr2 gene products. First, we synthetized and sequenced the cDNA obtained from 6 dpf wild-type (n=3) and genotyped cnr2upr1/upr1 (n=3) single larvae, all of which carried the deletion (Δ2: CT, Fig. 1C). Next, we performed Western blots with an anti-CB2 Ab raised against an epitope located before the first transmembrane domain of human CB2, to allow detection of a putative truncated protein. We prepared protein extracts from dissected adult brains of wild-type (n=3), and genotyped cnr2upr1/upr1(n=3) animals. As predicted, we found a CB2-specific band at ∼40 kDa in all wild-type (Fig. 1D, top left panels, Wt1 and Wt2 are shown) but not in any cnr2upr1/upr1 extracts (−/−1 and −/−2 are shown). Notably, no shorter CB2-KO-specific product was found, arguing that the truncated protein was unstable. We further validated specificity of the CB2-Ab in HEK293 cells (Fig. 1D lower left panels), which we transfected with a tagged CB2 construct (SEP-CB2) detected at 70 kDA. We probed tubulin expression in all samples (Fig. 1D, lower bands in left panels) and quantified it to determine the relative CB2 expression (graphs on the right). Taken together, we demonstrated that CB2 was expressed in adult fish brain but absent in cnr2upr1/upr1, and that it was a null allele resulting in viable homozygote larvae and adult totally devoid of CB2.
Absence of CB2 affects the swimming PDR
To determine if CB2 could have a role in complex behaviors, we assessed swimming behaviors using a previously established PDR assay, 29 in which we measured traveled distance and CO during four successive 10-min-long light (L) and dark (D) periods. We recorded simultaneously wild-type and mutant larvae individually distributed in 48-well plates. L/D cycling was started after 30 min of adaptation to darkness. We graphed the averaged traveled distance/larva/min for the entire recording time (Fig. 2A, Wt black and KO red, N=5, n=120 larvae/genotype). A highly reproducible swimming pattern (=PDR) emerged: lower swimming activity in L, which strongly increased in D periods. Most drastic changes were always occurring immediately after a light change and leveling out over the remainder of the period. Thus, we decomposed the analysis into successive (left panels) and cumulative (right panels) post-transition (=first min after an L/D or D/L change, Fig. 2B), and nontransition (=remainder of a period, Fig. 2C).
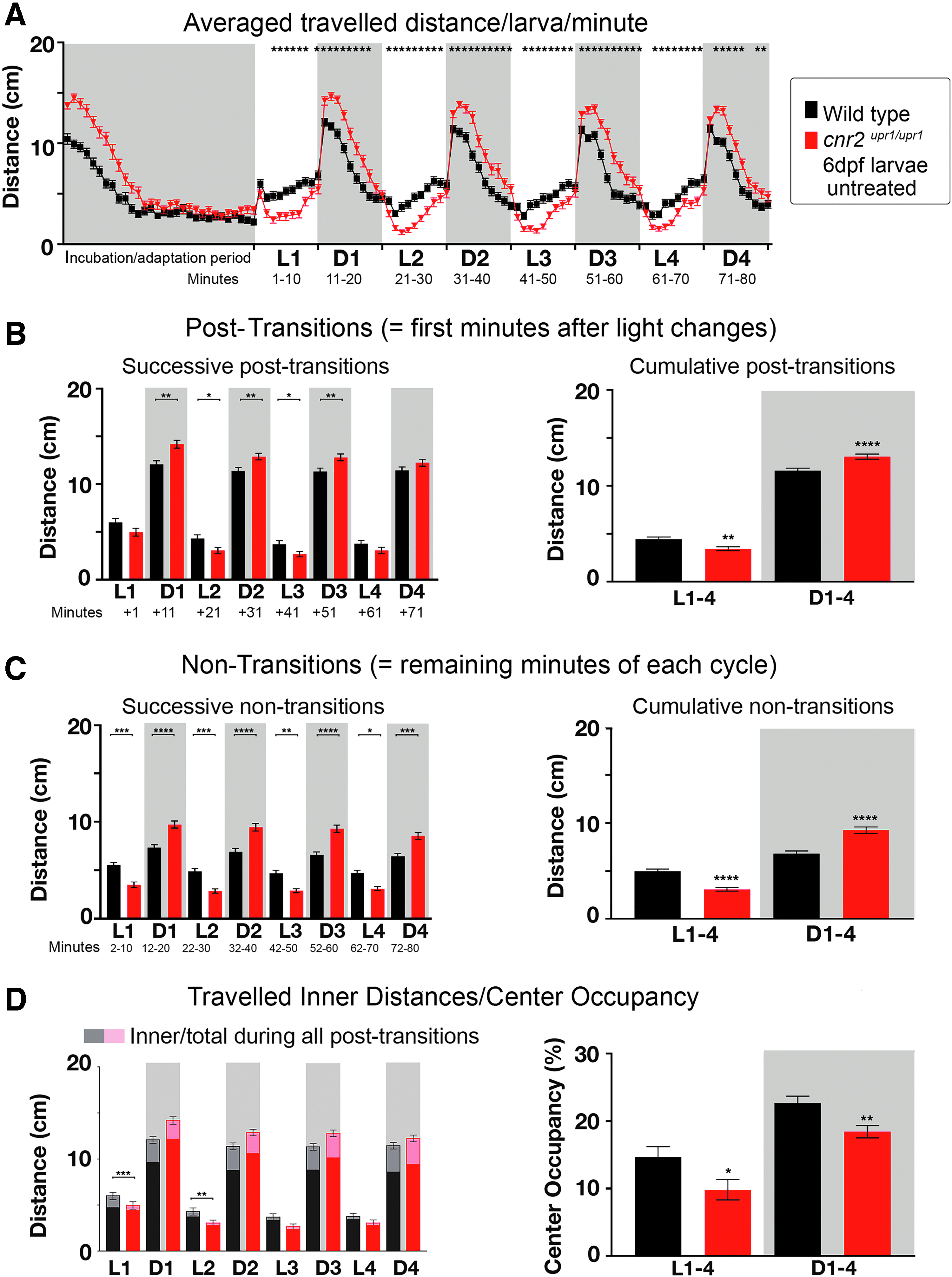
PDR in untreated 6 dpf wild-type and cnr2upr1/upr1 larvae.
Notably, cnr2upr1/upr1 traveled significantly less in L and significantly more in D periods than wild-type animals (* in Fig. 2A), displaying a clear genetic-dependent PDR. In L periods, mutant larvae traveled slightly, but significantly less in cumulative post-transitions (right panel, white Wt=4.45 cm/min vs. KO=3.43 cm/min, p<0.001) as well as in all nontransitions (Fig. 2C, successive: left panel white, p<0.05; cumulative: right panel white Wt=4.97 cm/min vs. KO=3.08 cm/min, p<0.0001). In D periods, mutant traveled significantly more in post-transitions (Fig. 2B, successive: left panel gray, p<0.01 except in D4; cumulative: right panel gray Wt=6.80 cm/min vs. KO=9.25 cm/min, p<0.001) and in all nontransitions (Fig. 3C, successive: left panel gray, p<0.001; cumulative: right panel gray Wt=11.57 cm/min vs. KO=13.04 cm/min, p<0.0001). Thus, mutant animals were significantly less active in L and more active in D periods, suggesting a possible CB2 modulation of light-dependent hypo/hyperactivity.
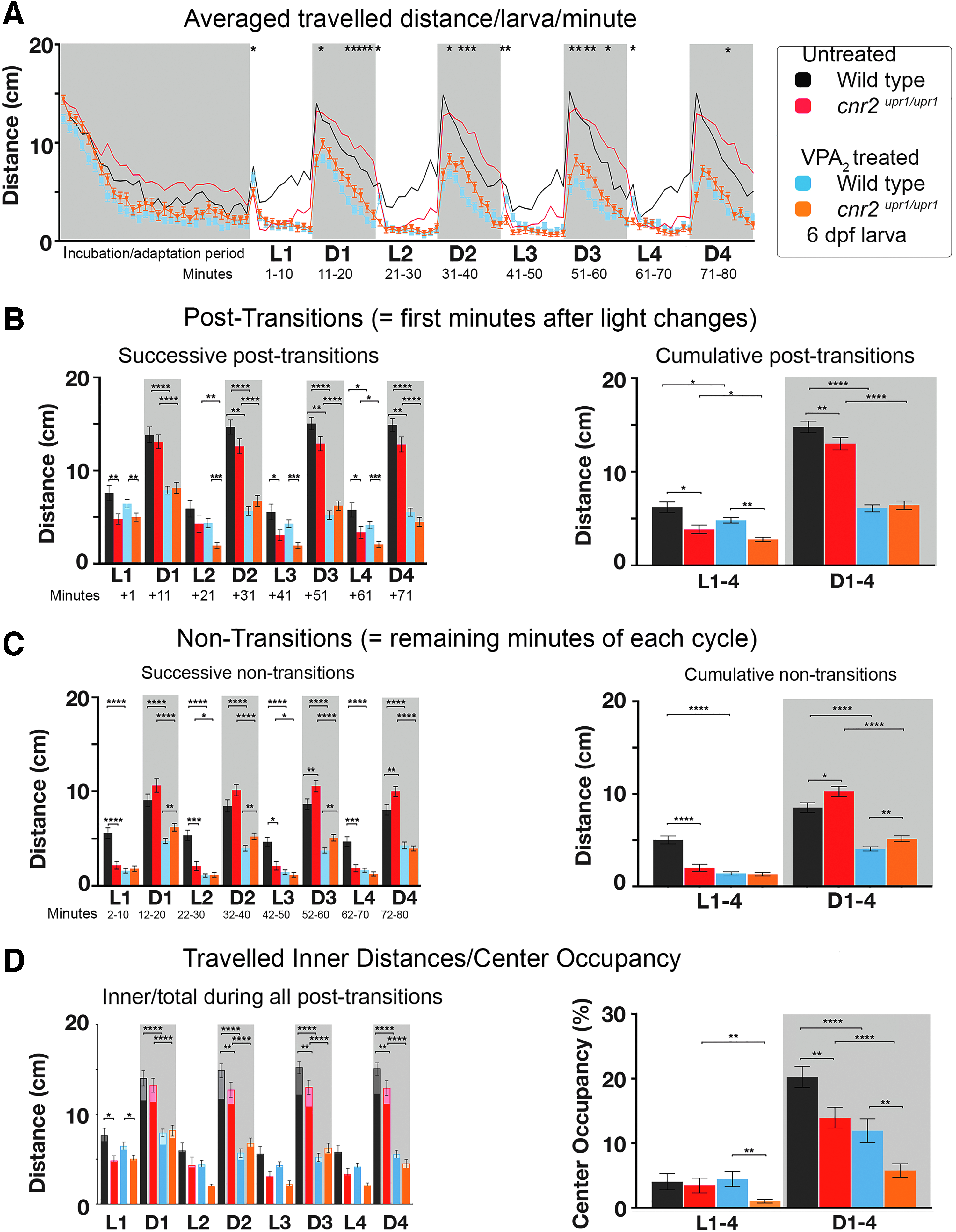
PDR in untreated and VPA [2 mM] (VPA2)-treated wild-type and cnr2upr1/upr1 6 dpf larvae.
Inner traveled distances were not significantly and consistently different between wild-type and mutant larvae (Fig. 2D left panel), but when we calculated the CO, it was significantly lower in mutants in D/L (right panel white COWt=14.70% vs. COKO=9.80%, p<0.05) and L/D post-transitions (gray COWt=22.70% vs. COKO=18.40%, p<0.001). Thus, CB2-KO animals spent less time in the center independently of the total distance traveled, suggesting that mutant larvae were avoiding open spaces more than wild-type animals. Taken together, animals lacking CB2 were hypoactive in L, hyperactive in D periods, and had decreased CO. Those results led us to postulate that CB2 was modulating complex behaviors.
Animals lacking CB2 respond differently to anxiolytic drug VPA
We previously showed that larvae treated with VPA [2 mM] (VPA2) had an altered PDR with overall lower swimming activity. 30 To assess a possible involvement of CB2, we set up parallel VPA2 treatments of wild-type and mutant larvae. After recording, we graphed the averaged traveled distance/larva/min (Fig. 3A, N=5, untreated Wt black n=40; untreated KO red n=40; WtVPA2 blue n=80; and KOVPA2 orange n=80). All VPA2-treated larvae were exhibiting a strong overall decreased activity similar at most recorded time points, independently of the genotype. We found a few significant differences with mutant traveling less than wild-type animals in D/L post-transitions (Fig. 3B, successive: left panel white, p<0.01; cumulative: right panel white WtVPA2 blue=4.82 cm/min vs. KOVPA2 orange=2.77 cm/min, p<0.0001). Also, in D periods, treated mutant traveled slightly but significantly more than treated wild-type larvae in nontransitions (Fig. 3C, successive: left panel gray, p<0.01 except D4; and cumulative: right panel gray WtVPA2 blue=4.06 cm/min vs. KOVPA2 orange=5.16 cm/min, p<0.005). Thus, these results suggested that VPA-triggered decreased swimming activity was partially modulated by CB2.
Inner traveled distances in D periods were reduced in both treated wild-type and mutant (Fig. 3D, left panel gray, p<0.01) and the CO was reduced (right panel gray box COWt black=20.26% vs. COWt-VPA2 blue=13.94%, p<0.001; COKO red=13.94% vs. COKO-VPA2 orange=5.75%, p<0.0001). In L periods, the CO was also significantly reduced in treated versus nontreated mutants (white, COKO red=3.42% vs. COKO-VPA2 orange=0.99%, p<0.0001). Thus, CO was strongly decreased in all treated animals and even more so in CB2-KO animals.
Animals lacking CB2 respond differently to anxiogenic drug PTZ
To assess a possible involvement of CB2 in modulating the previously described inverted PDR induced by PTZ7.5,30,34,40 we set up parallel PTZ7.5 treatments in wild-type and CB2-KO larvae. We graphed averaged traveled distance/larva/min (Fig. 4A, N=6, untreated Wt black n=56; untreated KO red n=56; WtPTZ7.5 green n=96; and KOPTZ7.5 khaki n=96). All PTZ7.5-treated larvae displayed the expected inverted PDR: stark hyperactivity in L, which was momentarily decreased in D periods independently of genotype. However, in L periods, hyperactivity was significantly stronger in mutants in all D/L post-transitions (Fig. 4B, successive: left panel white, p<0.01; and cumulative: right panel white WtPTZ7.5 green=13.56 cm/min vs. KOPTZ7.5 khaki=16.55 cm/min, p<0.001) and also in nontransitions (Fig. 4C, successive: left panel white, p<0.001; and cumulative: right panel white WtPTZ7.5 green=13.56 cm/min vs. KOPTZ7.5 khaki=16.55 cm/min, p<0.001). Thus, in the absence of CB2, the PTZ-induced hyperactivity in L was enhanced. In D periods, all treated larvae strongly reduced hyperactivity immediately after a light change. However, the reduction was smaller in mutants in post-transitions (Fig. 4B, successive: left panel gray p<0.01 except D3; and cumulative: right panel gray WtPTZ7.5 = 9.93 cm/min vs. KOPTZ7.5 = 11.98 cm/min, p<0.0001), and in nontransitions (Fig. 4C, successive: left panel gray p<0.05 except in D2; and cumulative: right panel gray Wt PTZ7.5 = 9.79 cm/min vs. KOPTZ7.5 = 11.49 cm/min, p<0.01). Thus, in the absence of CB2, the PTZ-induced hypoactivity in D was decreased.
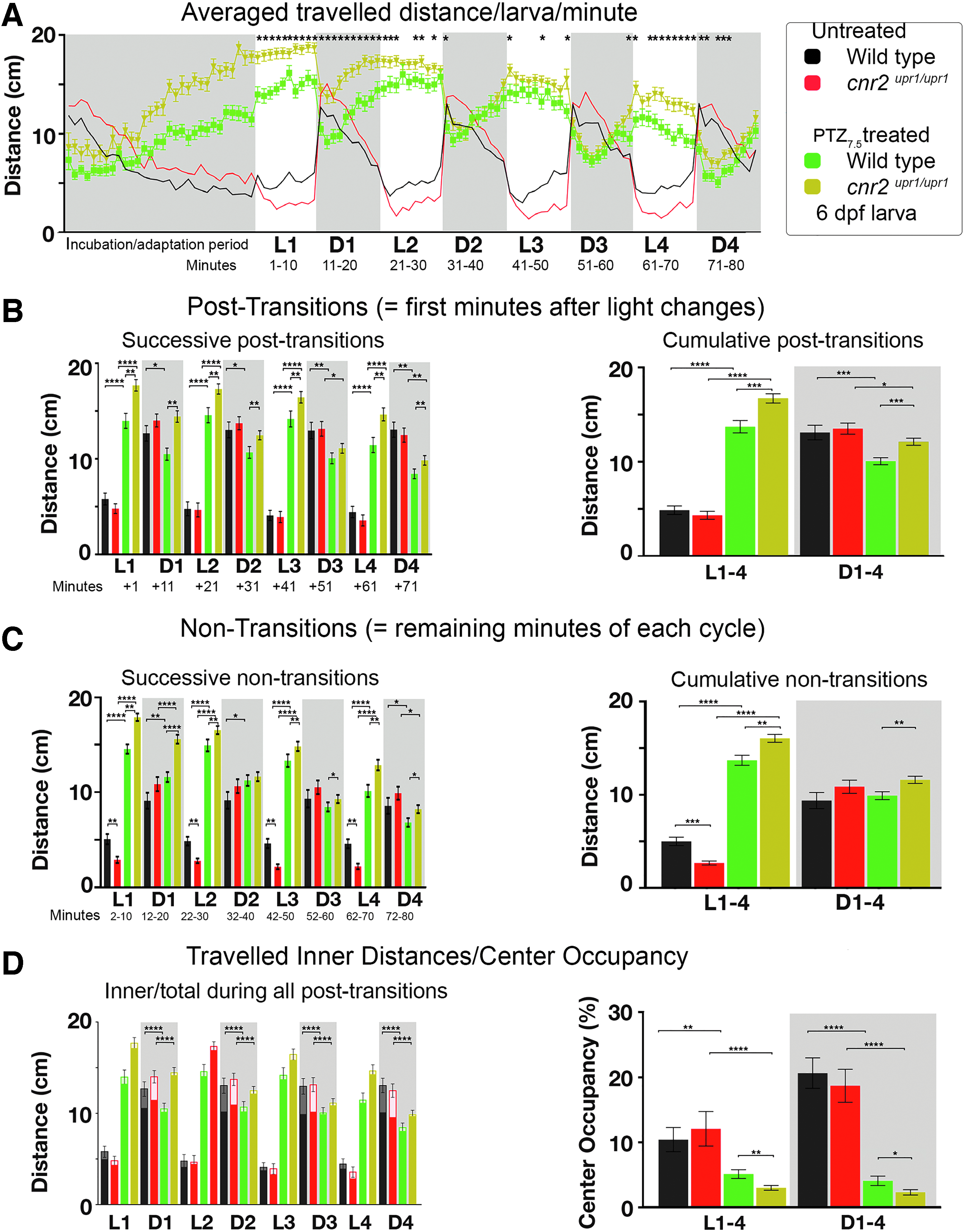
PDR in untreated and PTZ [7.5 mM] (PTZ7.5)-treated wild-type and cnr2upr1/upr1 6-dpf larvae.
Inner traveled distances were strongly reduced in all PTZ7.5-treated animals, but only in D periods (Fig. 4D, left panel, gray boxes, p<0.0001). The CO was strongly reduced in L (right panel white box COWt black=10.38% vs. COWt-PTZ green=5.04%, p<0.0001; and COKO red=12.06% vs. COKO-PTZ khaki=2.91%, p<0.0001) and in D periods (gray box COWt black=20.68% vs. COWt-PTZ green=4.00%, p<0.0001; and COKO red=18.72% vs. COKO-PTZ khaki=2.21%, p<0.0001). Remarkably, the CO reduction was even more pronounced in CB2-KO animals. Taken together, PTZ-triggered inverted PDR and decreased CO were significantly altered in the absence of CB2, suggesting that CB2 was a modulator of the PTZ anxiogenic effects.
A subset of CB2 ligand activity alters the PDR differently in wild-type and CB2-KO larvae
To assess if the PDR could detect CB2 ligand activity, we treated wild-type and mutant larvae with agonists (JWH-133 and HU-380) and antagonists (AM-630 and SR-144528). All ligands were pretested for overexposure-induced side effects (as detailed in the Materials and Methods section), and we analyzed further the PDR after treatment with the highest well-tolerated concentration (Fig. 5).
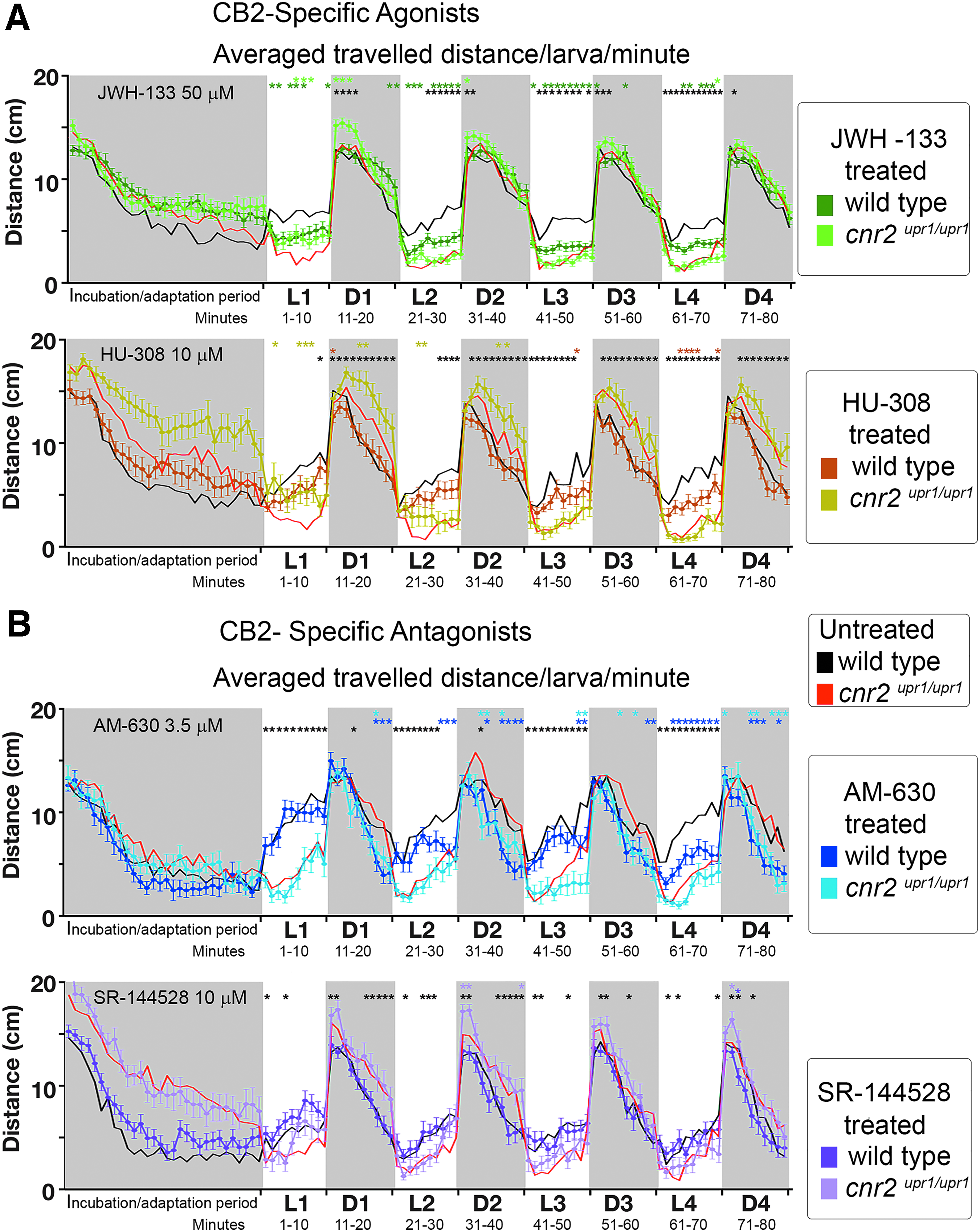
PDR in untreated and CB2 agonist (JWH-133 and HU-308)- and CB2 antagonist (AM-630 and SR-144528)-treated wild-type and cnr2upr1/upr1 6-dpf larvae.
With JWH-133 (top panel in Fig. 5A, Wt-JWH-13350 dark green n=24, and KO-JWH-13350 light green n=24), when we compared untreated versus treated wild type (black vs. dark green), we found that treated animals traveled significantly less in L (dark green* middle row, 30/40 in L and 4/40 in D) possibly as a result of CB2 binding and activation. Furthermore, when comparing untreated versus treated mutants (red vs. light green), we found few significant differences (light green* top row, 4/40 in L and 4/40 in D), meaning there was little activity detected in the absence of CB2, suggesting a good ligand specificity and low off-target effects. Thus, we concluded that JWH-13350 altered PDR, possibly in a CB2-specific manner.
With HU-308 (bottom panel in Fig. 5A, Wt-HU-30810 brown n=24, and KO-HU-30810 khaki n=24), when we compared untreated versus treated wild type (black vs. brown), we found very few significant differences (brown*middle row, 6/40 in light and 1/40 in dark) and likewise when comparing untreated versus treated-CB2-KO larvae (red vs. khaki and khaki* top row, 6/40 in L and 4/40 in D) pointing to a weak in vivo effect at well-tolerated concentrations.
With AM-630 (top panel in Fig. 5B, Wt-AM-6303.5 dark blue n=24, and KO-AM-6303.5 light blue n=24), when we compared untreated versus treated wild type (black vs. dark blue), we found that treated animals traveled less during nontransitions (dark blue* middle row 13/40 in L and 14/40 in D), possibly as a result of CB2 binding and activation. When comparing untreated versus treated mutant larvae (red vs. light blue), we found a few significant differences mostly in D periods (light blue*, top row 2/40 in L and 12/40 in D), suggesting that the detectable AM-6303.5-induced in vivo effect might be CB2 specific in L, but be off-target effects in D periods.
With SR-144528 (bottom panel in Fig. 5B, Wt-SR-14452810 purple n=24 and KO-SR-14452810 pink n=24), when we compared untreated versus treated wild type (black vs. purple), we found no significant differences (purple* middle row 0/40 in L and 1/40 in D) and likewise with untreated versus treated mutant larvae (red vs. pink, and pink* top row 2/40 in L and 12/40 in D) pointing to a weak in vivo effect at well-tolerated concentrations. Taken together, we elicited detectable in vivo effects with two of four tested CB2-ligands, which altered the PDR significantly at a subset of time points in wild type but not in mutants, arguing for CB2 specificity of the observed effects.
Discussion and Conclusions
To assess CB2 involvement in complex behaviors during vertebrate development, we generated with CRISPR-Cas9 technology CB2-KO animals and tested homozygote (cnr2upr1/upr1) larvae in a PDR swimming behavior assay. We showed that mutant animals were swimming significantly less in light, more in the dark, and avoiding open spaces more than wild type. Thus, we provide evidence for CB2 involvement in complex larval behaviors.
Hyperactivity and hypoactivity associated with, but not limited to, light changes are well-accepted measures of anxiety-like behaviors in rodents and have been also explored in adult fish41–43 although not yet extensively in larvae.44,45 We tested 6-dpf larvae because at this developmental stage, animals swim upright and exhibit complex behaviors comparable with adults. Using larvae presents major experimental advantages such as enabling upscalability. The small size (∼2 mm) and relative permeability of young larvae simplify chemical treatments that can be simply added to the water and will penetrate the animal by simple diffusion. Weekly spawning (∼100 eggs/couple/week) can provide ample number of animals for parallel testing of various concentrations of compounds. Center avoidance is another classical measure of anxiety-like behavior.46,47 As described previously, wild-type larvae swim mostly near the walls but travel more in the center during L/D post-transitions. 29 So, we measured inner traveled distances in post-transitions. However, variation of inner distances might simply reflect variation of the total activity, so we expressed relative distances traveled as a ratio: inner traveled distance/total traveled distance×100 to obtain the percentage of CO, providing an inverse reading of center avoidance. Therefore, we established anxiety-like in vivo parameters for fish that open new experimental avenues.
Next, we showed that when treating larvae with the anxiolytic drug VPA, the PDR was strongly reduced similarly in wild-type and mutant larvae, but with stronger hypoactivity in D/L post-transitions and more activity in dark nontransitions in the latter. VPA (or valproate) is a broad-spectrum anxiolytic drug
31
that increases gamma-aminobutyric acid (GABA) turnover, inhibits glutamate/N-methyl-
PTZ is commonly used in animal models to induce anxiety and seizure-like activity that is principally mediated via GABAA inhibition.35,51 We and others have previously shown that with a fixed concentration of PTZ [7.5 mM] (PTZ7.5), a strong inverted PDR could be induced in wild-type larvae, namely hyperactivity in light periods and hypoactivity in dark periods.30,34,40 Treated CB2-KO larvae had consistent heightened inverted PDR in light periods. Involvement of CB2 in the PTZ-elicited GABAA inhibition was previously shown in rodents 52 and offers a potential explanation. However, a greater sensitivity to treatments of mutant larvae, as well as possible additive effects occurring in parallel signaling pathways, cannot be excluded at this point. Testing of different doses of VPA and PTZ as well as cotreatments will help elucidate CB2 involvement.
We also measured the effect on the PDR with four known CB2 ligands and showed that we elicited with two of them, PDR alterations that were possibly CB2 mediated. Our data are proof of principle that such an approach could be further developed into an effective means to screen ligand-binding efficacy, specificity, as well as drug safety in a whole organism. However, a few major drawbacks must be addressed before exploiting this approach on a large scale. First, possibly because of the mode of administration of the ligands (directly into the water), we had to use very high concentrations of ligands to elicit a detectable response. This significantly narrowed the testable range of concentrations before reaching toxic levels. Alternative means of drug administration should be explored such as food additives. Second, we found an internal variation of the PDR across experiments in untreated wild-type and mutant larvae alike, rendering phenotypic differences in dark periods less consistent across experiments with different treatments especially when using smaller sample size (n<40). However, we found that the robustness of the phenotype could be easily strengthened by augmenting the number of tested animals, and by always using nontreated wild-type and mutant animals from the same clutches in parallel runs to provide solid internal controls.
In summary, we present an innovative upscalable approach that can be coupled to automated readout for significant differences and applied to drug discovery pipeline to test new CB2 ligand lead compounds. Likewise, mutant lines for CB1, opioid receptors, or GABA subunits could be established and double or even triple-KO lines used as screening tools. Finally, the indispensable preclinical safety and efficacy testing needed for bringing new drugs to the market could be performed in zebrafish larvae, offering a cost-effective, fast, and easy alternative or complement to the more classical preclinical models.
Footnotes
Acknowledgments
This work was supported by grants from NIDA (DA040920 and R01DA037924) to G.Y. and from NSF (1736019) to M.B. A.A.-C., L.C.-C., and R.R.-M. received support from NIGMS-RISE (R25 GM061838). Statistical analysis was supported by NIMHD (2U54MD007587). The authors thank Aranza Torrado, Alexis Santana, Juan Cantres, and Tiffany Tossas for help with fish room maintenance and husbandry.
Author Disclosure Statement
No competing financial interests exist.