
Abstract
Abstract
The endocannabinoid system (ECS) has been shown to be of great importance in the regulation of numerous physiological and pathological processes. To date, two Class A G-protein-coupled receptors (GPCRs) have been discovered and validated as the main therapeutic targets of this system: the cannabinoid receptor type 1 (CB1), which is the most abundant neuromodulatory receptor in the brain, and the cannabinoid receptor type 2 (CB2), predominantly found in the immune system among other organs and tissues. Endogenous cannabinoid receptor ligands (endocannabinoids) and the enzymes involved in their synthesis, cell uptake, and degradation have also been identified as part of the ECS. However, its complex pharmacology suggests that other GPCRs may also play physiologically relevant roles in this therapeutically promising system. In the last years, GPCRs such as GPR18 and GPR55 have emerged as possible missing members of the cannabinoid family. This categorization still stimulates strong debate due to the lack of pharmacological tools to validate it. Because of their close phylogenetic relationship, the Class A orphan GPCRs, GPR3, GPR6, and GPR12, have also been associated with the cannabinoids. Moreover, certain endo-, phyto-, and synthetic cannabinoid ligands have displayed activity at other well-established GPCRs, including the opioid, adenosine, serotonin, and dopamine receptor families. In addition, the cannabinoid receptors have also been shown to form dimers with other GPCRs triggering cross-talk signaling under specific conditions. In this mini review, we aim to provide insight into the non-CB1, non-CB2 cannabinoid-related GPCRs that have been reported thus far. We consider the physiological relevance of these molecular targets in modulating the ECS.
Introduction
The Class A G-protein-coupled receptors (GPCRs), cannabinoid receptor type 1 (CB1) and cannabinoid receptor type 2 (CB2), have been widely confirmed as cannabinoid targets. These receptors have been shown to be involved in numerous physiopathological processes, including pain, inflammation, cancer, metabolic syndromes, hypertension, and neurodegenerative disorders. 1 Nonetheless, the complex pharmacology of the endocannabinoid system (ECS) and its wide implication in numerous biological functions suggest the existence of other receptors playing important physiological roles. Consequently, extensive research is currently focused on the identification of potential missing cannabinoid receptors.
Diverse Class A orphans or lately deorphanized GPCRs have been proposed and evaluated as possible ECS members. Nonetheless, the lack of selective ligands for these receptors along with their intricate signaling pathways is delaying a clear elucidation of their relationship with the ECS. Therefore, thus far no other GPCR has been categorized as the cannabinoid receptor type 3 by the International Union of Pharmacology. 2
Herein, we intend to provide an overview of the GPCRs that have been postulated as cannabinoid molecular targets and the current available evidence of their relationship with the ECS. Non-GPCR targets of the cannabinoids such as the peroxisome proliferator-activated receptors, ligand-gated ion channels, or transient receptor potential channels have been revised elsewhere and are beyond the scope of this review.3,4
GPR55 and GPR18
Several GPCRs have been postulated to be putative cannabinoid receptors, but so far, only GPR18 and GPR55 have been demonstrated to be targets of a wide variety of endogenous, phytogenic, and synthetic cannabinoid ligands. 4 Despite this fact, inconsistencies in pharmacological data in the literature are hampering their categorization.5,6
The cannabinoid-related class A GPCR GPR55 displays low sequence identity with CB1 and CB2 (∼13% and 14%, respectively). GPR55 is widely expressed in the brain, as well as in the peripheral system, co-localizing with the cannabinoid receptors in diverse tissues.7–9 This receptor displays G-protein coupling promiscuity associating with Gα13,8,10 Gαq/11, 11 Gα12, 11 or Gα12/138,12 depending on the cell line or tissue. GPR55 has been implicated in different physiopathological conditions such as cancer,13–15 pain,11,16,17 metabolic disorders,18,19 vascular functions,20,21 bone physiology, 22 and motor coordination. 23
The phospholipid lysophosphatidylinositol (LPI) is considered the endogenous GPR55 ligand.8,24,25 In fact, GPR55 has also been named the LPI1 receptor. 26 Numerous CB1 and CB2 ligands have also been reported to act as GPR55 modulators.6,27–29 However, significant pharmacological discrepancies have been found depending on the tested functional outcome. 6 For instance, the well-known phytocannabinoid Δ 9 -tetrahydrocannabinol (Δ 9 -THC) displayed activation of GPR55 according to certain reports,10,11 while it was unable to exert any effect in other functional assays.24,30 Cannabinoid ligands reported to be recognized by GPR55 and their intriguing pharmacology have been recently reviewed elsewhere. 31
Although its sequence presents low identity with CB1 and CB2 (∼13% and 8%), GPR18 has also been tightly associated with the ECS.4,32 High expression of GPR18 has been found in the lymphoid tissues, while it is moderately expressed in other organs such as lungs, brain, testis, or ovary.33,34 Initially, GPR18 was found to couple to Gαi/o; however, subsequent findings suggested the participation of the Gαq/11 transduction pathway as well.34–36 Different reports have shown the therapeutic potential of this target in the treatment of pathologies such as intraocular pressure, 37 cancer, 38 or metabolic disorders 39 among others.
N-arachidonoyl glycine (NAGly) has been suggested to be the endogenous GPR18 ligand by several research groups.32,34 However, other researchers were not able to confirm these data. 40 Recent investigations point to the existence of another endogenous GPR18 activator: the polyunsaturated fatty acid metabolite, Resolvin D2 (RvD2), which is mainly involved in inflammatory processes. 41 In addition, and despite the pharmacological divergences observed among some reports, GPR18 has been shown to recognize an array of CB1 and/or CB2 ligands of endogenous, phytogenic, or synthetic nature (reviewed by others).39,42
The pharmacological discrepancies on the appraisal of cannabinoids in these two receptors, as well as the lack of selective ligands targeting them, are delaying an insightful understanding of the relation of GPR55 and GPR18 with the ECS. These inconsistencies, which may rely on intrinsic properties of these GPCRs, or on the cell type or functional assay, need to be further studied. Intensive efforts are also focused on the structural understanding of these receptors, 43 as well as the development of more potent and selective pharmacological tools for the study of these promising therapeutic targets.
GPR3, GPR6, and GPR12
GPR3, GPR6, and GPR12 are three orphan Class A GPCRs that exhibit a very close phylogenetic relationship with the cannabinoid receptors CB1 and CB2 (Fig. 1). Indeed, they belong to the same cluster of receptors, the so-called MECA cluster (which consists of the melanocortin receptors, the endothelial differentiation GPCRs, the cannabinoid receptors, the adenosine binding receptors, and the orphan receptor subset GPR3, −6, and −12).44,45 Because of their phylogenetic proximity, these orphan receptors share common conserved residues and unique sequence motifs with CB1 and CB2. 46 According to Fredriksson et al. these orphan receptors may share a common ancestor with the cannabinoid receptors since they share the same chromosomal positions. 45
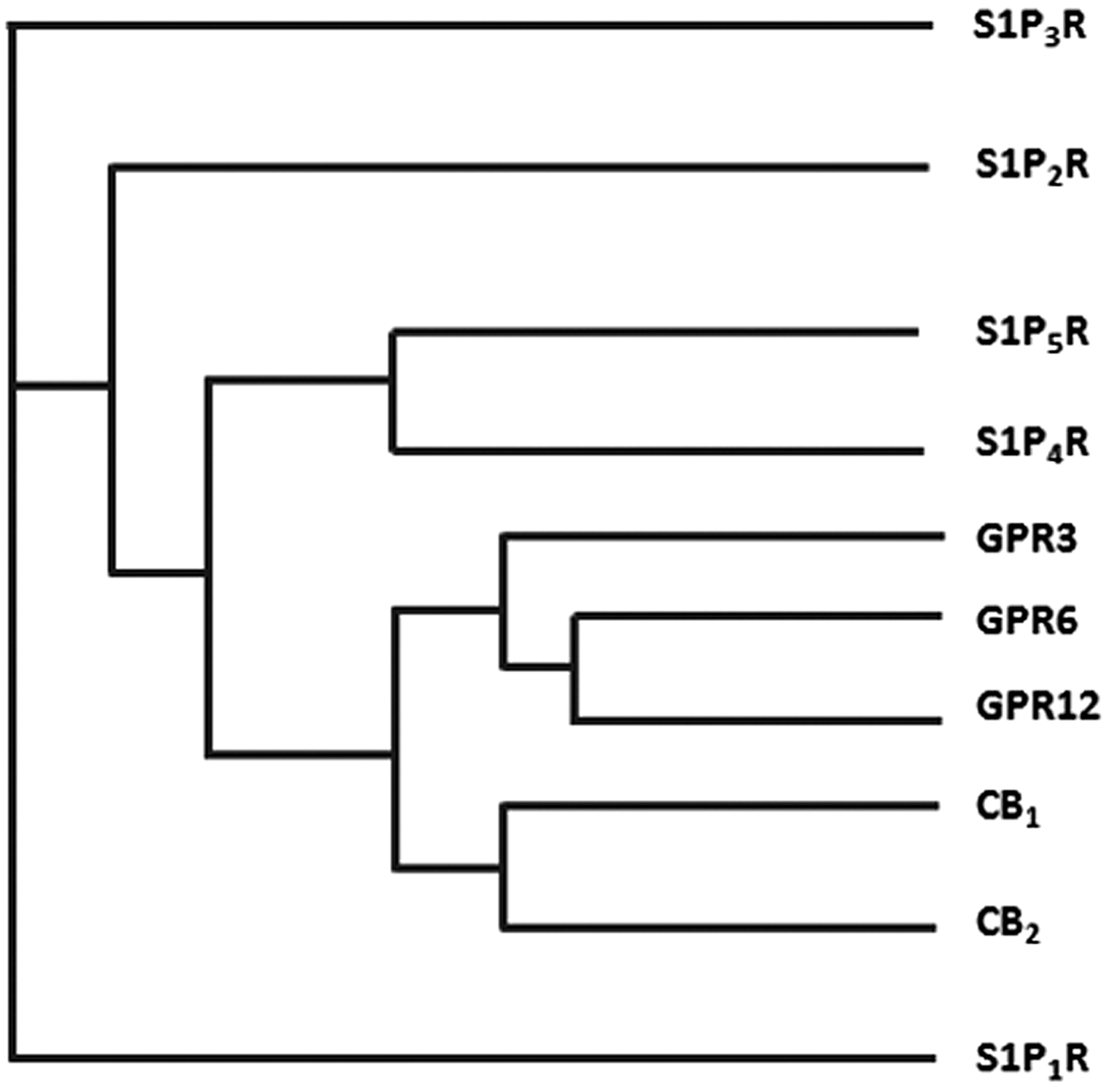
Phylogenetic tree of cannabinoid receptors and the closely related Class A GPCRs (S1PR family and the orphan receptors GPR3, GPR6, and GPR12). Data were obtained from GPCRdb.org. CB1, cannabinoid receptor type 1; CB2, cannabinoid receptor type 2; GPCR, G-protein-coupled receptor; S1P, sphingosine-1-phosphate.
GPR3, GPR6, and GPR12, which share over 60% of sequence similarity, were first cloned in 1995.47,48 These receptors constitutively activate adenylate cyclase by coupling to Gαs proteins. In fact, different groups have reported that when expressed in diverse cell lines, they can stimulate adenylate cyclase to levels similar in amplitude to agonist-activated GPCRs.47,49,50 In addition to Gαs, GPR6 and GPR12 have also been suggested to couple to Gαi/o,51,52 but further investigations are required to confirm this G-protein promiscuity.
GPR3, GPR6, and GPR12 are predominantly expressed in the brain and the reproductive system. 49 This family of constitutively active GPCRs is involved in neuronal differentiation and growth, as well as in the formation of synaptic contacts. 49 Therefore, their role in different neurological processes such as neurite outgrowth, 49 Alzheimer's disease,53–57 development of cerebellar granule neurons,58,59 neuropathic pain, 60 early phases of cocaine reinforcement, 61 emotional-like responses, 62 instrumental learning, 63 or Parkinson's disease64,65 has been studied. Other pathophysiological conditions such as oocyte maturation,66,67 dyslipidemia, 68 and cell proliferation 69 may also be impacted by the modulation of some of these receptors.
The bioactive lipids, sphingosine-1-phosphate50,52 and/or sphingosylphosphorylcholine, 51 have been proposed as endogenous ligands of these receptors (Fig. 2). However, other groups were not able to confirm this claim, and consequently, GPR3, GPR6, and GPR12 are still categorized as orphans.30,70,71 Interestingly, among the very few ligands discovered so far for these receptors, the nonpsychoactive phytocannabinoid cannabidiol (CBD) stands out as being able to target GPR3 and GPR6, 72 acting as a β-arrestin2 inverse agonist of both receptors. This functionality is of high interest in the GPR3 field because β-arrestin2 signaling at GPR3 has been directly linked to the manufacture of beta-amyloid plaque (Aβ1–40 and Aβ1–42) in Alzheimer's disease through complex formation with γ-secretase.56,57 Because CBD is an inverse agonist of this signaling pathway at GPR3, it may represent a potential tool for the reduction of amyloid pathology. Other phytocannabinoids and several endocannabinoids were also tested but so far none of them were found to modulate this family of orphan receptors.30,72
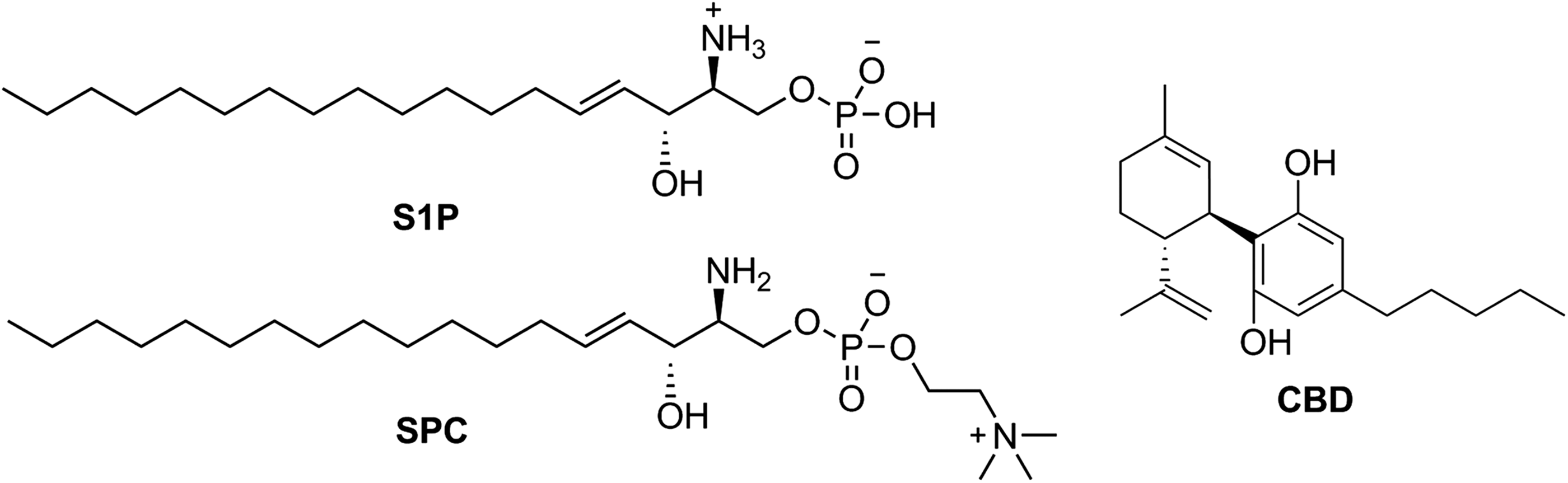
Structures of the putative GPR3, GPR6, and GPR12 endogenous ligands S1P and SPC and the GPR3 and GPR6 inverse agonist CBD. SPC, sphingosylphosphorylcholine.
So, a relationship between the cannabinoids and the orphan receptors GPR3, GPR6, and GPR12 has been evidenced. Nonetheless, extensive research and more pharmacological tools are needed to extract significant conclusions about the association of these receptors with the ECS and its ligands.
Alkylindole-Sensitive Receptors
As reported by different research groups, the well-known aminoalkylindole cannabinoid agonist WIN55,212-2 (Fig. 3) displays pharmacological functions independent of the cannabinoid receptors CB1 and CB2.73–75 This fact led to the identification of novel targets commonly referred to as the alkylindole (AI)-sensitive receptors.74,76,77 These cannabinoid-related receptors are modulated by AI derivatives, but not by other classes of cannabinoid ligands. 76 Diverse evidence suggests that the AI-sensitive receptors are Gαs-protein coupled receptors that are mainly expressed in microglia and astrocytomas.76–79 However, their biological functions, pharmacology, and therapeutic value remain to be unraveled due to the lack of selective pharmacological tools.
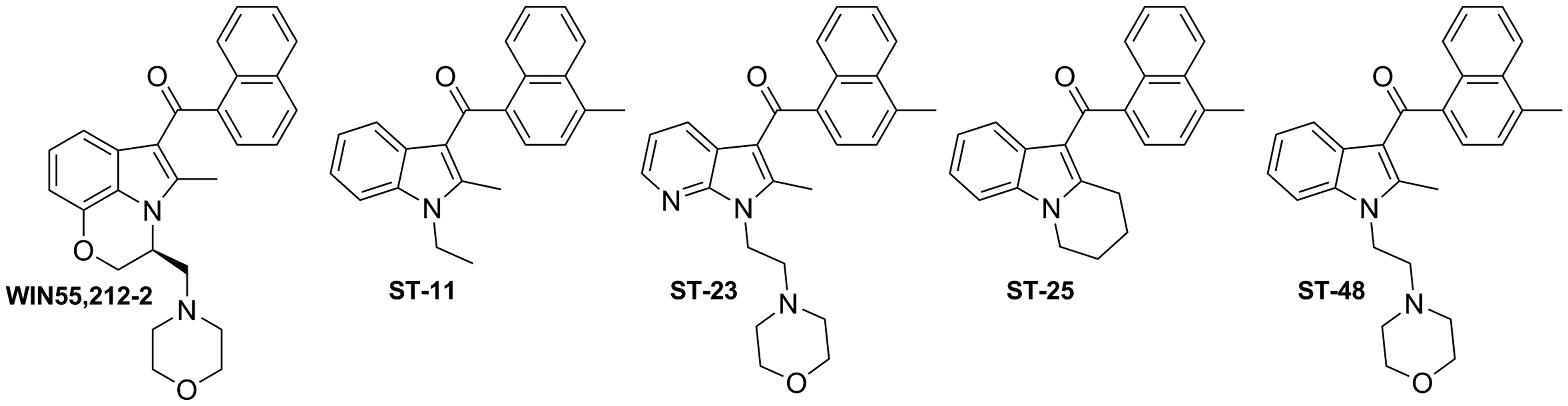
Alkylindole derivatives WIN55,212–2, ST-11, ST-23, ST-25, and ST-48.
Recent studies from Stella and coworkers revealed the role of AI-sensitive receptors in the modulation of microglial cell migration and proliferation highlighting their potential in the treatment of gliomas.77,78 Moreover, these authors have identified a series of naphthoyl AI derivatives, ST-11, ST-23, ST-25, and ST-48 (Fig. 3) among them, that bind to the AI-sensitive receptors. 78 These compounds display affinities in the nanomolar range when competing with [ 3 H]WIN55,212-2 in DBT (Delayed Brain Tumor) cells which endogenously express AI-sensitive receptors, while lacking CB1 and CB2 receptors. 80 Compound ST-11 stands out from this study because of its potency at AI-sensitive receptors, while not interacting with CB1 and CB2 receptors. In addition, in vitro assays revealed that this compound inhibits cell migration and proliferation in the aforementioned mouse glioma cell line, DBT. Further studies revealed that ST-11 can reduce glioblastoma growth in a syngeneic mouse model. 81
Even though extensive research is clearly needed to understand the pathophysiological function of these receptors, reported data suggest that AI-sensitive receptor agonists could represent a novel class of potential brain cancer antitumor drugs.
Cannabinoid-Related Oligomers
Numerous studies have shown that GPCRs, cannabinoid receptors among them, can exist and function as dimers or complexes of higher order.82–85 This oligomerization may affect receptor signaling, receptor trafficking, and ligand binding. The physiological relevance of such dimerization has not yet been fully established for the cannabinoid receptors; nonetheless, the presence of cannabinoid homo- and heterodimers in specific tissues has been intensely reported over the last years.
For the CB1 receptor, heteromers have been suggested to exist under certain physiological conditions with serotonin, 86 angiotensin, 87 opioid,88–90 GPR55, 91 somatostatin, 92 orexin,93,94 dopamine,95–97 and adenosine 98 receptors among others (Table 1). Although CB2 has been less investigated, recent research revealed that it may form heterodimers with CB1, 99 with GPR55,100,101 with the serotonin receptor 5HT1A, 102 or with the chemokine receptor CXCR4. 103 The expression of these heterodimers has been associated with different pathologies. For instance, the CB2−CXCR4 and the CB2−GPR55 dimers have been associated with cancer progression, while the CB1−A2A and the CB1−D2 heteromers have been suggested to have physiological implications in neurodegenerative disorders such as Huntington's or Parkinson's diseases. All these data suggest that the ECS interacts in a significant manner with several other endogenous systems.

CB1, cannabinoid receptor type 1; CB2, cannabinoid receptor type 2.
With regard to cannabinoid receptor homodimerization, more data have been published on CB1 homodimers than on their CB2 counterparts. The presence of CB1 receptor homodimers has been reported in different biological tissues,104–106 but their functional role has not been determined. In contrast, CB2 homodimers have been evidenced,107–109 but their pharmacological potential has not been explored yet.
In this field, bivalent ligands have emerged as promising new pharmacological entities and potential tools for the biological study of their respective dimeric receptors.110–113 Despite their poor pharmacokinetic properties, 114 bivalent ligands can exhibit enhanced activity and selectivity over their respective corresponding parent ligands offering unique pharmacological strategies. Bivalent ligands have been synthesized and evaluated for several GPCRs. Opioid,115,116 dopamine,117,118 and histamine 119 are some of the receptors for which a bivalent compound provided higher activity than their monomer counterparts. CB1 homobivalent120–122 and heterobivalent123–125 ligands have been also reported and explored. However, currently available receptor structural information challenges the fact that bivalent ligands can simultaneously bind to both receptors within the dimer, especially in the case of lipid receptors as the cannabinoids. 126 Therefore, novel drug design approaches to target dimers, as well as new techniques to determine bivalent binding, remain to be explored.
Homo- and heterodimerization likely influences the manner in which the ECS responds to ligands. Nevertheless, unambiguous data about their physical association in native tissues, as well as their pharmacology, are needed to clearly identify what biological functions are impacted by cannabinoid dimers.
Well-Established GPCRs Related to the Cannabinoids
Certain endo-, phyto-, and synthetic cannabinoid ligands have been shown to modulate well-known GPCRs. These GPCRs include members of established families such as the opioid, serotonin, muscarinic, dopamine, and adenosine families. For instance, the endocannabinoid anandamide has been shown to act at the adenosine receptor A3, 127 the muscarinic acetylcholine receptors M1 and M4, 128 and the serotonin receptors 5-HT1A and 5-HT2A 129 among others. In addition, phytocannabinoids such as Δ 9 -THC and CBD have been shown to modulate the μ− and δ−opioid receptors, 130 while other plant-derived compounds such as CBG (cannabigerol) and Δ 9 -THCV (tetrahydrocannabivarin) display activity at the 5-HT1A receptor.131,132 Likewise, synthetic cannabinoids, such as the CB1 inverse agonists taranabant (MK-0364) and rimonabant (SR141716), have also displayed activity in well-established targets. These include the adenosine A3 and the tachykinin NK2 receptors. 133
Some of these cannabinoid ligands have been proposed to interact allosterically with the aforementioned targets. It is worth mentioning that the efficacy that most of these cannabinoids exhibit toward these GPCRs is lower than the one displayed at the CB1 and/or CB2 receptors. Therefore, there is no evidence indicating a necessary recategorization of these receptors.
Other GPCRs
Because of their ability to recognize lipids and their relatively close phylogenetic relationship with CB1 and CB2, several other Class A orphan or recently deorphanized GPCRs such as GPR40, GPR43, GPR41, GPR120 (currently classified as free fatty acid receptors FFA1, FFA2, FFA3, and FFA4, respectively), GPR23, GPR92 (recently categorized as lysophosphatidic acid receptors LPA4 and LPA5), GPR84, GPR119, or GPR35 have been postulated as possible cannabinoid receptor candidates. 4 However, there is no available evidence since they do not meet some of the criteria established by the International Union of Pharmacology.4,70
Conclusions
Two cannabinoid receptors, CB1 and CB2, have been validated at the molecular level as the main targets of the ECS. These two GPCRs have been widely explored in the development of numerous pathophysiological processes, and their therapeutic potential for the treatment of different diseases has been extensively confirmed. 1 Great efforts are being made to structurally understand these receptors; in fact, CB1 in its inactive134,135 and active 136 states has been recently crystallized. Despite possible crystallization artifacts, these structures will help shedding light into the complex pharmacology of the cannabinoid receptors.
Growing evidence suggests that other cannabinoid or cannabinoid-like receptors remain to be identified as important players of the ECS. Different endogenous, phytogenic, and/or synthetic cannabinoid ligands have been reported to modulate GPCRs such as GPR18, GPR55, GPR3, GPR6, or the AI-sensitive receptors, among others. Pharmacological discrepancies and the lack of selective ligands for these receptors are delaying the characterization of their relationship with the ECS. Consequently, no CB3 receptor has yet been confirmed. 2
Adding more complexity to the ECS scenario, molecular interactions of the cannabinoid receptors with other GPCRs have been reported. Co-localization or co-immunoprecipitation data suggest the presence of cannabinoid homo- and heterodimers in specific native tissues. Cannabinoid receptor dimerization may not only influence the pharmacology of these receptors but also it may provide new signaling pathways through the interacting protomers. However, due to the lack of appropriate tools, there is still limited in vivo information about the expression of cannabinoid dimers. Hence, it remains a challenge to elucidate their therapeutic relevance under specific physiological conditions.
Currently, appropriate characterization of cannabinoid ligands should take into account the activity at the aforementioned GPCRs. Possible biased agonism of ligands, allosterism, or cross-talk signaling could be determining the intricate GPCR pharmacology. In addition, differential coupling and regulation of G-proteins or the formation of oligomers are among GPCR intrinsic properties that might be delaying the validation of novel potential cannabinoid targets. Therefore, further research is needed to fully understand the physiopathological role of these non-CB1, non-CB2 GPCRs in the modulation of the ECS.
Footnotes
Acknowledgments
The authors acknowledge research support from NIH/NIDA grants R01 DA003934 and K05 DA021358 (P.H.R.).
Author Disclosure Statement
No competing financial interests exist.