
Abstract
Aortic valve stenosis is one of the most common cardiovascular diseases in western countries and can only be treated by replacement with a prosthetic valve. Tissue engineering is an emerging and promising treatment option, but in-depth knowledge about the microstructure of native heart valves is lacking, making the development of tissue-engineered heart valves challenging. Specifically, the basement membrane (BM) of heart valves remains incompletely characterized, and decellularization protocols that preserve BM components are necessary to advance the field. This study aims to characterize laminin isoforms expressed in healthy human aortic valves and establish a small animal decellularized aortic valve scaffold for future studies of the BM in tissue engineering. Laminin isoforms were assessed by immunohistochemistry with antibodies specific for individual α, β, and γ chains. The results indicated that LN-411, LN-421, LN-511, and LN-521 are expressed in human aortic valves (
Introduction
Aortic valve stenosis is one of the most prevalent cardiovascular disorders in the western world.
1
To date, no medical therapies are available, making treatment options limited to surgical or endovascular replacement with a biological or mechanical prosthetic valve. However, biological valves have limited durability due to gradual structural deterioration eventually leading to graft failure, and mechanical valves necessitate life-long treatment with potent anticoagulants.
2
The ideal heart valve replacement is considered to be an off-the-shelf valve that does not exert an immune response and has the capacity to grow and be repopulated by living cells. Therefore, tissue-engineered heart valves, both decellularized and
Despite the widely recognized importance of the extracellular matrix (ECM), and the significant interest in heart valve tissue engineering, the important basement membrane (BM) of heart valves remains poorly characterized. The BM plays key roles in cell adhesion, migration, function, and differentiation. 14 Laminins, consisting of an α, β, and γ chain, are the first BM components to be expressed in mammalian tissues. 15 Their importance in cell function and tissue formation is illustrated by the fact that several laminin chain isoforms are vital for normal embryonic development. 14 However, the laminin isoforms and their expression pattern in heart valves have not been characterized, and the role of laminins in tissue engineering remains largely unknown.
Decellularized scaffolds provide an opportunity to investigate the effects of individual ECM components in a regenerative context relevant for tissue engineering. They have previously been used to study how heparan sulfate proteoglycans affect the differentiation of endoderm-derived stem cells on a decellularized lung scaffold. 16 However, it is well known that different tissues require different methods for decellularization17–20 as the process inevitably has detrimental effects on the ECM. 21 Therefore, optimization of a decellularization protocol for the specific tissue in question to achieve effective cell removal while limiting detergent exposure is critical.
The aim of the present study was two-fold. First, to characterize the laminin isoforms present in human and rat aortic valves, and their distribution patterns. Second, to develop a decellularized rat aortic valve scaffold that preserves laminin and other BM components for future studies on how the BM influences cell behavior in heart valve regeneration.
Materials and Methods
Human tissues
Whole human hearts were obtained from Legacy Donor Services Foundation, Florida. Legacy Donor Services Foundation receives donor referrals from hospitals and medical examiners serving the state of Florida. Written informed consent was obtained from the donor through registration as a tissue and organ donor with Donate Life America, Donate Life Florida, or registering to be a donor when renewing or obtaining a driver's license. Alternatively, written informed consent was obtained from the donor's next-of-kin by a Donor Management Coordinator from Legacy Donor Services Foundation. Hearts intended for biomedical research were offered to Nova Southeastern University by the Legacy Donor Services Foundation. Inclusion criteria were donor age 18–60 years and no history of valvular heart disease. Institutional Review Board approval was obtained from the Regional Ethics Review Committee in Stockholm, Sweden (Protocol Number: 2017/2268-31). Hearts (
Animals
Rat aortic roots were procured from female Sprague–Dawley rats 8–12 weeks of age (Charles River). All animals were cared for in accordance with the rules and regulations of Karolinska University Hospital and Karolinska Institutet. Institutional Review Board approvals for animal experiments were obtained from the Regional Committee on the Ethics of Animal Experiments, Linköping, Sweden (Protocol Numbers: ID 6-17 and S190-12).
Rat aortic valve procurement
Rats were anesthetized through isoflurane inhalation and then immediately euthanized by inhalation of CO2 or decapitation. The thoracic cavity was accessed by dividing the ribs and sternoclavicular joints and excising the sternum. All intrathoracic organs were removed en bloc. The aortic valve was dissected from the heart under a microscope as a U-shaped aortic conduit as described by others.22,23 Briefly, a myocardial cuff of ∼1–2 mm and part of the anterior mitral valve leaflet were preserved, whereas the brachiocephalic artery, left common carotid artery, and left subclavian artery were ligated with 6-0 sutures (Ethicon) at their origin. The descending aorta was divided at the level of the aortic valve. For normal controls, the ascending aorta was divided at the origin of the brachiocephalic artery and the aortic root was then embedded in OCT and snap frozen.
Decellularization
A perfusion decellularization method based on earlier protocols5,17 was used. In brief, the aortic conduit was cannulated with a 1.1 mm intravenous catheter (B. Braun Medical), which was connected to a tubing pump (Ismatec) through a custom-built stainless steel bottle cap to enable recirculation of reagents in a 250-mL bottle (Supplementary Fig. S1A). Aortic conduits were perfused with 1% sodium dodecyl sulfate (SDS; Merck Sigma-Aldrich) in deionized water for 3 h at a flow rate of 2 mL/min, followed by 15 min with deionized water, 30 min with 1% Triton X-100 (Merck Sigma-Aldrich) in deionized water and 12–24 h with phosphate-buffered saline (PBS), changed twice. The aorta was divided at the origin of the brachiocephalic artery to facilitate enzymatic treatment. Removal of residual nucleic acids was achieved by processing aortic roots with 90 U benzonase endonuclease (E1014; Merck Sigma-Aldrich) on a see-saw rocker (Cole-Parmer) overnight at room temperature (RT). The remaining aortic roots were processed on the see-saw rocker overnight in PBS and then snap frozen in OCT or used for DNA quantification.
Histology and immunohistochemistry
Human valve cusps (
DNA quantification
DNA content per decellularized aortic valve (
Statistics
One-way analysis of variance with Bonferroni corrections for multiple comparisons was used to identify differences between groups.
Results
Healthy human aortic valves express laminin α4, α5, β1, β2, and γ1 chains
Aortic valves were from male donors 50–60 years of age. Upon gross morphologic assessment, one of the hearts was hypertrophic and had significant atherosclerosis in the main coronary arteries (Table 1). All three aortic valves were tricuspid and macroscopically normal. A single aortic valve cusp was used from each donor, that is, one right coronary cusp, one left coronary cusp, and one noncoronary cusp. H&E staining and double staining for elastin and collagen I confirmed the absence of plaques and an overall normal histology with an intact three-layered structure (Fig. 1). The noncoronary cusp had a markedly vascularized interstitium, the left coronary- and right noncoronary cusp had limited or no vascularization.

Histology of donor aortic valve cusps.
Donor Demographics and Heart Morphology
M, male; F, female; CAD, coronary artery disease; NCC, noncoronary cusp; RCC, right coronary cusp; LCC, left coronary cusp.
The endothelial BM of the human aortic valve cusps stained positive for laminin α4, α5, β1, β2, and γ1 chains (Fig. 2). All chains were uniformly present in all three cusps, forming a continuous monolayer. The same chains were also expressed in the BM of the interstitial vessels observed in two of the cusps. The interstitium itself was mostly negative for the laminin chains, except in minor regions. Laminin α1, α2, α3, β3, and γ2 could not be detected in any of the valve cusps. This suggests that the human aortic valve endothelial BM consists of LN-411, LN-421, LN-511, and LN-521.

Laminin chain expression in human aortic valves. The endothelial basement membrane covering the valve cusps were positive for laminin α4, α5, β1, β2, and γ1, as well as the endothelial lining of interstitial vessels and minor patches in the interstitium. Images of α1, α2, α3, β3, and γ2 show absence of immunoreactivity. A representative image of ≥3 tissue sections from each of the three donor valve cusps is shown. All images shown at 20 × magnification. Scale bar represents 100 μm.
To assess the relevance and utility of the rat model, three rat aortic roots were stained for laminin α4, α5, and γ1. Rat aortic valve cusps also stained positive for α4, α5, and γ1, indicating the presence of the same laminin isoforms as in human valves (Fig. 3). The distribution pattern was uniform for all three laminin chains, which formed a convergent monolayer, continuous with the endocardium and aortic endothelium.

Expression of laminin α4, α5, and γ1 in rat aortic valves. The endothelial basement membrane stained positive for all three laminin chains. Minimal immunoreactivity was seen in the interstitium. A representative image of ≥4 tissue sections from three animals per group is shown. All images captured with 20 × magnification. Scale bar represents 100 μm.
Decellularization with SDS and Triton X-100 preserves BM components
To establish a small animal model for studies of cell–laminin interactions, we sought to assess the effectiveness and effects on the BM of a decellularization protocol for rat aortic valves adapted from earlier studies.5,17 Aortic roots were exposed to 1% SDS for 1, 3, 6, or 12 h (
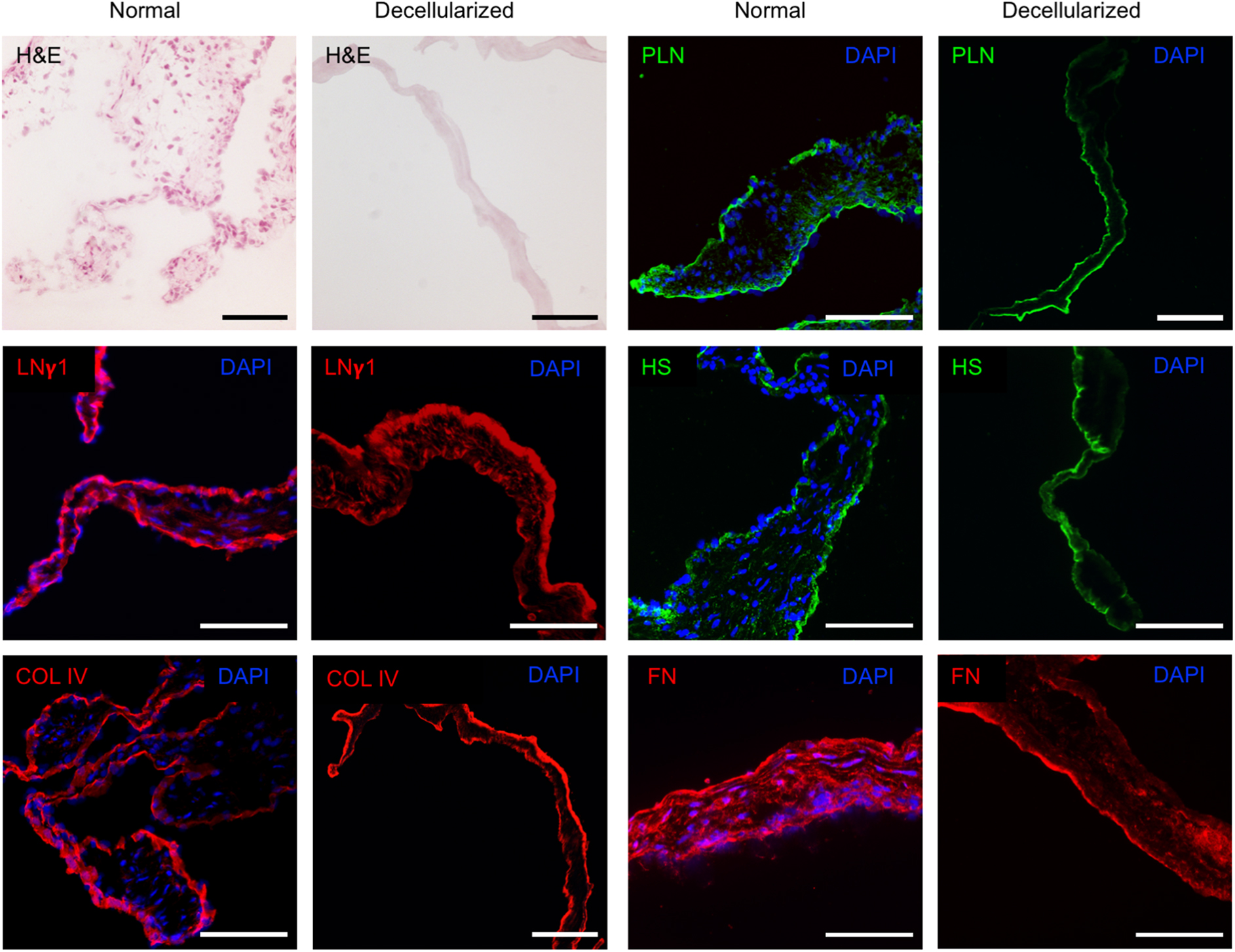
Histology of normal and decellularized rat aortic valves. H&E images and DAPI staining of cell nuclei reveal absence of intact nuclei after decellularization. Laminin γ1, collagen IV, perlecan, heparan sulfate, and fibronectin was visualized in both normal controls and decellularized valves, with a largely preserved distribution pattern after decellularization. A representative image of ≥3 tissue sections from three animals per group is shown. All images were captured at 20 × magnification. Scale bar represents 100 μm. LNγ1, laminin γ1; COL IV, collagen IV; PLN, perlecan; HS, heparan sulfate; FN, fibronectin.
To confirm the integrity of the BM, tissue sections of decellularized rat aortic valves (
Limited detergent exposure combined with benzonase endonuclease results in minimal residual DNA
Due to their extremely small mass, we were unable to obtain reliable weight measurements from the valve cusps. However, by using fine dissection tools, the three valve cusps could be excised from the aortic root in a highly reproducible fashion, enabling assessment of DNA content per valve, that is, all three cusps excised from the same specimen. Normal control aortic valves (
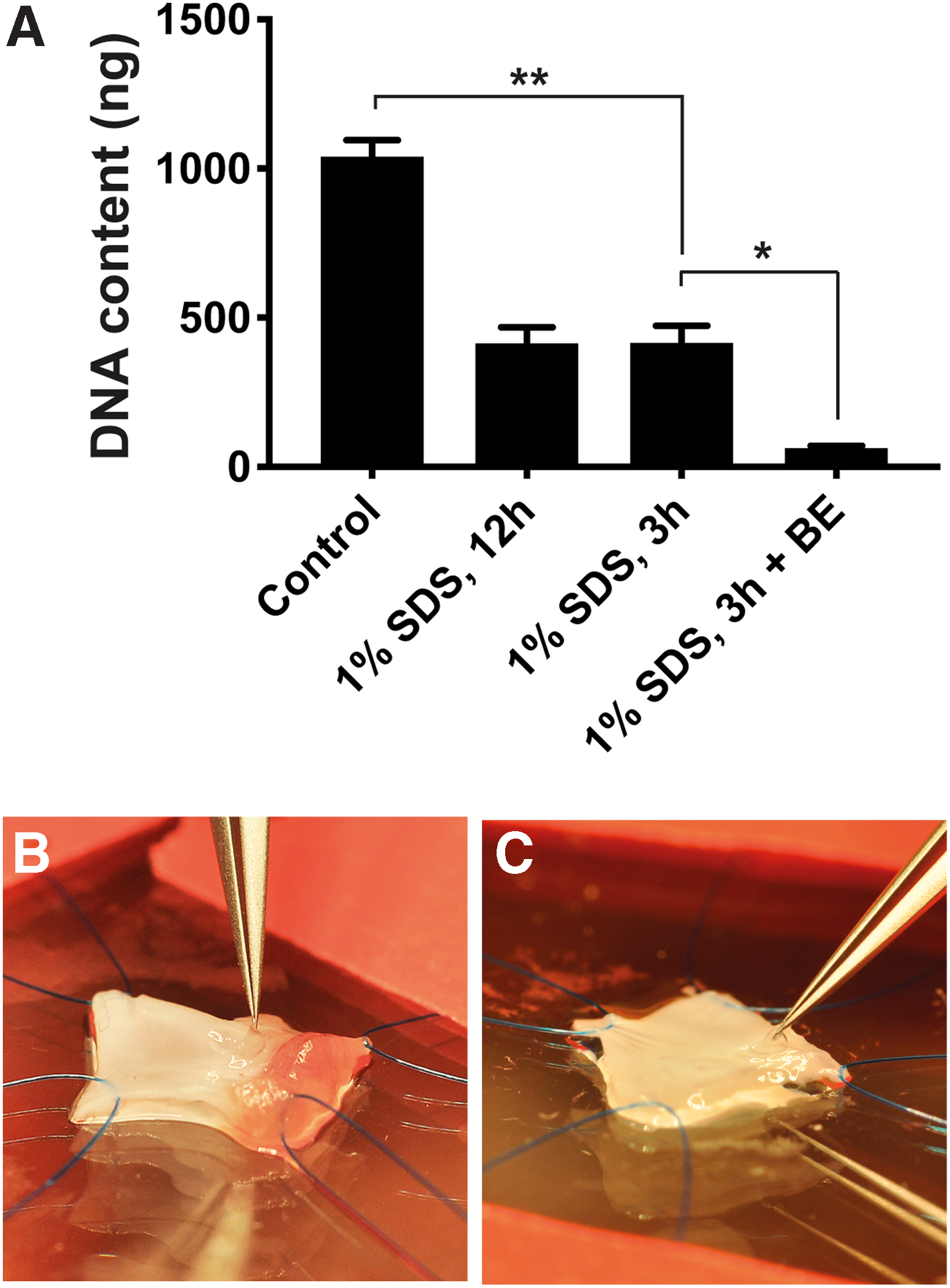
Quantification of DNA in rat aortic valves before and after decellularization.
Discussion
The present study describes, for the first time, the specific laminin chains present in healthy human aortic valves. We show that the main laminin isoforms expressed in the endothelial BM of heart valves are LN-411, LN-421, LN-511, and LN-521, and that the isoforms are conserved across mammalian species. The same laminin isoforms were also expressed in small interstitial vessels that may exist in valve cusps.26,27 Interestingly, small traces of laminin that appeared to be located in the interstitium rather than in the endothelial BM were observed. These findings are consistent with previous reports on laminin isoforms expressed in vascular endothelial BMs.28–33 However, cardiac valvular cells and their ECM are unique in structure and function,34–36 and our study provides critical data on the BM structure and microenvironment of valvular cells necessary for recreating the native BM in tissue-engineered heart valves.
Laminins are key components of vascular BMs and have important, isoform-specific functions.
37
Laminin α4 is ubiquitous in the BM of blood vessels and most endothelial BMs also express α5.29–31
However, their distribution pattern and relative expression levels vary considerably between different vessel types, which affects the properties of the BM. For example, the expression patterns of α4 and α5 are known to determine the tightness of endothelial junctions and thus the permeability of the endothelium in the context of lymphocyte extravasation, with α4 isoforms being permissive and α5 isoforms being restrictive.38–41
Furthermore, evidence suggests that α4 has a role in angiogenesis as well as the regulation of endothelial cell survival,42,43 indicating a role in endothelialization of tissue-engineered valves and vascular grafts. Indeed, laminin coating, although of an undefined isoform, has been shown to promote endothelialization while decreasing intimal hyperplasia in aortic grafts
In conclusion, laminins play key roles in the cardiovascular system, and the presence or lack of specific laminin isoforms in tissue-engineered heart valves may thus have a profound impact on their viability. However, the role of laminins in tissue engineering needs to be defined to assess their true relevance in this context, and further studies on small animal scaffolds are an appropriate next step. While decellularized rat aortic valves have been described by others,5,23,47–49 we present a protocol with reduced detergent exposure, which results in both effective cell removal and preservation of laminin and other key BM components. Scaffolds produced through limited detergent exposure, and thus with a better preserved BM,20,21,50 will be a useful tool in future mechanistic studies.
The number of donated heart valves in our study was limited due to the sparse availability of healthy human heart valves. We did not obtain clinical data, such as cause of death, comorbidities, and clinical risk factors, which could have added insights into whether valves were at risk of calcific degeneration. However, all valves had a normal gross morphology and histology. Furthermore, the laminin expression in all human aortic valves was fully consistent between specimens, and data are supported by current literature on laminins in the cardiovascular system.28–33,37
To the best of our knowledge, this is the first report to describe the laminin chains expressed in aortic valves and their localization. Given the importance of laminins for a number of cellular functions, this provides crucial data for future attempts to recreate the aortic valve through tissue engineering. The presence or absence of laminins, and other ECM components such as perlecan,51–54 heparan sulfate, 16 and fibronectin4,55 is bound to affect the viability of tissue-engineered heart valve grafts, but other factors that influence graft viability also need to be considered. Aside from a lack of appropriate ECM components, structural deterioration of tissue-engineered heart valve grafts may be due to (i) immunogenicity, 56 (ii) thrombogenicity, 57 or (iii) incomplete or inappropriate valvular endothelial or interstitial cell repopulation.3,4,8 The relative importance of laminins and other ECM components, as well as other factors that may affect the long-term viability of a tissue-engineered heart valve, will need to be determined in future studies.
Conclusion
We have shown that the aortic valve BM contains LN-411, LN-421, LN-511, and LN-521. We also describe a modified decellularized rat scaffold with a largely preserved BM for future mechanistic studies. Further studies are required to quantify the laminin isoforms and determine their role in heart valve tissue engineering. Additional studies are also necessary to assess the effect of detergents used during decellularization on the functional properties of ECM components in heart valve regeneration.
Footnotes
Acknowledgments
The authors would like to thank H. Thomas Temple and Chris Agle at the Legacy Donor Services Foundation, as well as Richard Jove, former director, and Vladimir Beljanski at the Cell Therapy Institute, Nova Southeastern University, Florida, for excellent collaboration. They would also like to thank Ewa Ellis (Division of Transplantation Surgery, Karolinska Institutet), Stephen Strom (Division of Pathology, Karolinska Institutet), and Alejandro Soto-Gutierrez (Dept. of Pathology, University of Pittsburgh) for sharing their protocols and equipment with them, and John Couchman (Biotech Research and Innovation Center, University of Copenhagen) for kindly providing the anti-perlecan antibody.
Author Disclosure Statement
K.-H.G. and S.R. are cofounders of the company IsletOne AB. The other authors declare no competing interests.
Funding Information
This study was supported by grants from the Swedish Research Council, Young Investigator (2013-03590), the Magnus Bergvall Foundation (2017-02416), the Wallenius Foundation and a private donation from Mr. Fredrik Lundberg. None of the funding sources had any role in study design, data collection or analysis, decision to publish, or preparation of the article.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.