
Abstract
Tissue-engineered bone grafts seeded with mesenchymal stem cells (MSCs) have been sought as a replacement for bone grafts currently used for bone repair. For production of osteogenic constructs, MSCs are isolated from bone marrow (BM) or other tissues, expanded in culture, then trypsinized, and seeded on a scaffold. Predifferentiation of seeded cells is often desired. We describe here bone progenitor cells (BPCs) obtained by direct osteogenic differentiation of unprocessed BM bypassing isolation of MSCs. Human BM aspirates were incubated for 2 weeks with a commonly used osteogenic medium (OM), except no fetal calf serum, serum substitutes, or growth factors were added, because responding stem and/or progenitor cells were present in the BM milieu. The adherent cells remaining after the culture medium and residual BM were washed out, expressed high levels of bone-specific alkaline phosphatase (ALP) on their surface, demonstrated high ALP activity, were capable of mineralization of the intercellular space, and expressed genes associated with osteogenesis. These parameters in BPCs were similar and even at higher levels compared to MSCs subjected to osteogenic differentiation for 2 weeks. The yield of BPCs per 1 mL BM was 0.71±0.39×106. In comparison, the yield of MSCs produced by adhesion of mononuclear cells derived from the same amount of BM and cultured in a commercial growth medium for 2 weeks was 0.3±0.17×106. When a scaffold was added to the BM-OM mixture, and the mixture was cultured in a simple rotational bioreactor; the resulting BPCs were obtained already seeded on the scaffold. BPCs seeded on scaffolds were capable of proliferation for at least 6 weeks, keeping high levels of ALP activity, expressing osteogenic genes, and mineralizing the scaffolds. Autologous rat BPCs seeded on various scaffolds were transplanted into critical-size calvarial defects. Six weeks after transplantation of polylactic acid/polyglycolic acid scaffolds, 76.1%±18.3% of the defects were filled with a new bone, compared to 37.9%±28.4% in the contralateral defects transplanted with the scaffolds without cells.
Introduction
In the case of the loss of large bone segments as a result of trauma or a tumor resection, the bone defect is usually filled with a porous scaffold or a bone graft is used. 1 With recent progress in regenerative medicine, attempts have been made to seed scaffolds with adult mesenchymal stem cells (MSCs) found in the bone marrow (BM), fat, and other tissues (reviewed in Refs.2–5 ). Transplanted MSCs are assumed to undergo osteogenic differentiation in the bone environment or alternatively are predifferentiated before the transplantation. 6
The low frequency of MSCs in BM requires expansion of isolated cells in vitro, which depends on preselected fetal calf serum (FCS). The differentiation potential of MSCs at later passages (40–50 population doublings) is often low due to MSC senescence, 7 or tumor transformation. 8
We hypothesized that osteogenic progenitors sought for tissue engineering bone grafts could be derived directly from BM without isolation of MSCs, but rather by treating of unprocessed BM with osteogenic factors. We assumed that osteogenic agonists would target all the cells in BM that are capable of osteogenic differentiation, whereas a part of these bone progenitor cells (BPCs) is lost during isolation of MSCs. In addition, osteogenic differentiation within unprocessed BM would be supported by growth factors and cytokines that are present in the BM milieu and by signals from adjacent cells.
We present here a method for derivation of BPCs directly from BM, and confirm the osteogenic character of these cells. When BPCs are produced in a rotational bioreactor in the presence of a scaffold, they populate the scaffold. We show that scaffolds seeded with BPCs during differentiation of unprocessed BM promote bone repair in vivo.
Materials and Methods
Production of human BPCs
Human BM aspirates were purchased from Lonza Inc. BM was mixed with an osteogenic medium (OM) [alpha-MEM (Sigma), 0.2 mM L-ascorbic acid-2-phosphate (Mg salt, n-hydrated; Fluka), 100 nM dexamethasone (Sigma), 2 mM GlutaMAX™-I Supplement (Invitrogen), 100 units/mL penicillin, 0.1 mg/mL streptomycin, and 0.25 mg/mL amphotericin B] at the ratio 1:5, plated onto 24-well plates or cell culture dishes (all from NUNC) precoated with 10 μg/mL bovine fibronectin (Biological Industries), and cultured for 2 weeks in a cell incubator. No FCS was added to OM. In experiments with scaffolds, 0.2 mL BM was combined with 2.5 mL OM and with a fibronectin-precoated scaffold in a 50-mL polypropylene tube with a filter cap (Techno Plastic Products AG). The tubes with the BM-OM mixture were placed on a HUG tube rotator (MRC) and rotated in the cell incubator at 6–10 rpm/min at an angle of 30° for 2 weeks. After 1 week, half of the volume of each culture was replaced with a fresh OM, and incubation continued for another week. At 2 weeks, all cultures were washed 3 times with MEM-alpha, and BPCs adherent to the culture vessels or scaffolds (BPCs p0) were used for analysis. All BPC batches used in the experiments were derived from different BM donors.
Production of rat BPCs
BM from one femur/animal (∼1 mL) of 14-week-old Fischer344 inbred rats was flushed with 5 mL of OM. About 0.6 mL of the BM/OM mixture was either seeded into a 24-well plate or combined with 2.5 mL OM and rotated with a fibronectin-precoated scaffold disc as described for human cells. Rat BPC cultures were completely washed after 1 week. At 2 weeks, the scaffolds were washed and used for transplantation.
Isolation of human MSCs
Human MSCs were isolated as described elsewhere, 9 adopting the commonly used protocol. 10 MSCs expressed all the main MSC markers, CD105, CD29, CD73, CD44, CD90, CD166, CD9, CD106, and CD146, and were negative for the hematopoietic cell marker CD45. 9 MSCs were capable of trilineage differentiation into osteoblasts, adipocytes, and chondrocytes. 9 All MSC batches used in the experiments were derived from different BM donors.
Osteogenic differentiation of human MSCs
Cells were plated at 3000 cells per 1 cm2 in GM overnight. Then, GM was replaced by the OM containing 10% heat-inactivated FCS (Biological Industries). The OM was changed twice a week.
Scaffolds
Polylactic acid (PLA) (70/30 L, DL-PLA), polylactic/polyglycolic acid copolymer (PLA/PGA) 50/50%, and PLA/ceramics (70/30 L,DL-PLA compounded with 80% beta-tricalcium phosphate by mass) were purchased from Kensey Nash Corporation. PLA/PGA 75/25% scaffolds used for animal studies and translucent PLA/PGA 50/50 scaffolds used for microscopic evaluation were a generous gift from Dr. Shulamit Levenberg, Technion, Haifa, Israel. All the scaffolds used in in-vitro studies were of 10-mm diameter and 1–2-mm thickness; the scaffolds used for animal studies were of 5-mm diameter and 2-mm thickness.
Quantitation of MSCs and BPCs with Calcein-AM
Cells were incubated with 5 μM Calcein-AM (Invitrogen) in Dulbecco's modified Eagle's medium (DMEM) without phenol red in 24
Quantitation of BPCs growing on scaffolds with Alamar Blue
Scaffolds seeded with cells were incubated with 10% Alamar Blue (Invitrogen) in the OM at 37°C for 2 h. Fluorescence of the incubation medium was measured at Ex/Em 530/590 nm. For a standard curve, serial dilutions of BPCs were plated in a 24-well plate and allowed to adhere overnight. Adherent cells were incubated with Alamar Blue in the presence of empty scaffolds. The exact cell numbers were determined with Calcein-AM after the Alamar Blue assay.
Measurements of alkaline phosphatase activity
Cells growing in a 24-well plate or on a scaffold were incubated with 1 mL alkaline lysis buffer for 1 h on ice. 100 μL lysate combined with 400 μL phosphatase substrate solution [20 mg/mL of p-nitrophenol phosphate in 5 mL alkaline buffer solution diluted 1:3 with ddH2O] was incubated at 37°C for 10 min. The reaction was stopped with 500 μL EDTA-NaOH stop solution [20 g NaOH plus 37.22 g Na2EDTA in 500 mL ddH2O]. The absorbance of the samples was read at 404 nm. Serial dilutions of the dephosphorylated product, p-nitrophenol, were prepared for a standard curve. The results were expressed as nmol p-NP/mL/min/10,000 live cells. All the reagents were from Sigma-Aldrich.
Flow cytometry analysis of ALP expression
BPCs or MSCs were incubated with saturating concentrations of anti-ALP antibodies (clone B478; R&D) in an FACS buffer [2% BSA (Sigma-Aldrich) and 2% human blocking serum (Chemicon) in DMEM without phenol red (Sigma), pH 7.4] for 30 min on ice and then analyzed on FACSAria (BD).
Quantitation of calcium deposition
Cells in a 24-well plate or on a scaffold were mixed with 250 μL 0.5N HCl. The lysates were shaken overnight at 4°C to extract calcium and then centrifuged at 13,000 rpm for 2 min. The assay was set up in 96-well plates using the Calcium Liquicolor kit (Stanbio Labs), according to the manufacturer's instructions.
Fast Blue RR staining for ALP activity
Cells growing in 24-well plates or on scaffolds were fixed with a mixture of 1:50 dilution of citrate solution (Sigma-Aldrich) and acetone at 2:3 ratio for 30 sec and then stained with an alkaline phosphatase staining kit (Sigma-Aldrich), according to the manufacturer's instructions.
Alizarin red staining for calcium deposits
Cells growing in 24-well plates or on scaffolds were fixed with 4% paraformaldehyde for 15 min at room temperature, washed, and stained with 5 mg/mL Alizarin Red S (Sigma-Aldrich) for 1 min.
Microscopic evaluation
Stained cells were analyzed using either bright-field or fluorescent microscopy (Zeiss AxioVert 40) and photographed with AxioCam MRC5.
RNA isolation
The RNeasy kit (Qiagen) was used to isolate RNA. BPCs growing on scaffolds were lyzed directly. RNase-free DNase (Qiagen) was used for DNase digestion. RNA was frozen at −80°C. Aliquots of RNA were thawed on ice, and RNA integrity was confirmed by electrophoresis in 2% agarose gel. The RNA yield from BPCs produced on PLA and PLA/ceramic scaffolds was 9.8±5.0 and 11.5±4.1 μg per scaffold, respectively (mean±SD; n=4).
Real-time polymerase chain reaction
Real-time polymerase chain reaction was performed using Applied Biosystems 7500 machine. TaqMan-based primers for alkaline phosphatase (ALP; Hs 00758162_m1; NM_000478.3), bone sialoprotein (BSP; Hs 00173720_m1; NM_004967.3), matrix Gla protein (MGP; Hs 00179899_m1; NM_000900.2), and hyaluronan-mediated motility receptor (RHAMM; Hs 00234864_m1; NM_012484) were purchased from Applied Biosystems. Thirty-eight nanograms of RNA was used per each cDNA sample. Delta-Ct was calculated by the subtraction Ct value of the TATA box-binding protein (TBP) gene (NM_003194.3). The results for BPCs and differentiating MSCs were normalized to gene expression in undifferentiated MSCs.
Animal studies
Animal studies were performed under the approval of the Institutional Animal Care and Use Committee of the Tel Aviv University. An ∼5-mm-diameter defect was produced with a high-speed, water-cooled, diamond wheel-shaped bur in each side of the calvarial bone of inbred Fischer 344 rats. Care was taken not to injure the underlying brain tissue. Syngeneic BPCs seeded on scaffolds were transplanted into the right side. The left side was transplanted with the same type of scaffold, but without cells. Eight animals per each type of scaffold were transplanted. A control group of 4 animals was transplanted with the scaffold alone without cells on the right side, whereas the contralateral side remained untreated. The animals were euthanized at 6 weeks after transplantation. Skull tissues were subjected to Faxitron contact microradiography, then embedded in methyl methacrylate, and sectioned coronally at 4-μm thickness roughly through the middle of the 2 defects. Sections were stained with Masson's TriChrome to identify the new bone in the defect area (dark blue). A rectangle of 1.5-mm width was superimposed on the defect, and bone and total areas were measured. Percentage of the new bone in the defect was calculated. The study was double blinded.
Statistical analysis
Data are presented as mean±SD. Comparisons were made by using a two-tailed Student's t-test or a one-way analysis of variance for experiments with more than two subgroups. Probability values were considered statistically significant at p<0.05.
Results
Human BPC yields
The yield of BPCs p0 was 0.71±0.39×106 per 1 mL BM (n=23). When the BPC culture was continued for another week, the yield of BPCs was 1.05±0.36 (n=13). The same BM batches were used for production of MSCs. MSCs were produced by plating 10×106 MNCs (corresponded to ∼1 mL BM) in GM containing 20% FCS and by culturing adherent cells for 2 weeks (MSCs p0). The yield of MSCs p0 was 0.3±0.17×106 (n=21). Significantly, a lower yield of MSCs (p=0.00043) suggested that conditions of the described BM culture are more favorable for stem cell/progenitor cell proliferation than commonly used MSC culture conditions.
Osteogenic differentiation of human BPCs p0
When MSCs were induced to differentiate in an OM, the level of enzymatic activity of ALP gradually increased within 2 weeks (Fig. 1A). For production BPCs p0, unprocessed BM was also cultured in the OM for 2 weeks. ALP activity of the resulting BPCs was significantly higher than ALP activity of MSCs subjected to osteogenic differentiation for 1 week (p<106) and for 2 weeks (p<0.0025) (Fig. 1A). Gradual increase of ALP enzymatic activity in MSCs subjected to osteogenic differentiation paralleled the kinetics of its surface expression analyzed by flow cytometry. As illustrated by histograms in Figure 2B, most undifferentiated MSCs did not express ALP, although a small population was positive, probably due to spontaneous differentiation. At 1 week after addition of the OM, a significant number of MSCs expressed ALP, but a clear-cut population of ALP-negative cells still remained, and only by 2 weeks of differentiation, most of MSCs became positive. The flow cytometry analysis of ALP expression in BPCs p0 demonstrated a homogeneous ALP expression (a histogram at the bottom of Fig. 1B has practically a normal bell-shape distribution). The mean fluorescence intensity (MFI) of BPCs was higher than MFI of MSCs differentiated for 2 weeks; however, this difference was not statistically significant (Fig. 1C).

Comparison of mesenchymal stem cells (MSCs) and bone progenitor cells (BPCs) derived from human bone marrow (BM).

Proliferation of human BP cells produced on scaffolds.
Production of human BPCs on scaffolds
To demonstrate proliferation of BPCs on scaffolds, the scaffolds with BPCs p0 were washed and rotated in a fresh OM for additional 1–4 weeks (3–6 weeks from the initiation of culture). During the third week, the number of cells per scaffold practically doubled, although the proliferation rate of BPCs seeded on PLA/ceramic scaffolds was somewhat lower than that of BPCs seeded on PLA and PLA/PGA scaffolds (Fig. 2A). Extended culturing of BPCs on scaffolds resulted in practically an exponential growth (Fig. 2B). ALP activity of BPCs p0 growing on PLA and PLA/ceramic scaffolds was 8.8±2.4 and 10.7±0.8 nmol p-NP/min/10,000 cells, respectively (n=3 for each scaffold). BPC growth and differentiation are illustrated by Figure 3A, showing BPCs seeded on PLA/PGA 50/50 translucent scaffolds. Individual live cells stained with Calcein-AM could be seen in greater quantities at 6 weeks than at 2 weeks after initiation of BM culture. Histochemistry for ALP activity and for calcium deposits confirmed that proliferated BPCs retained their osteogenic character.
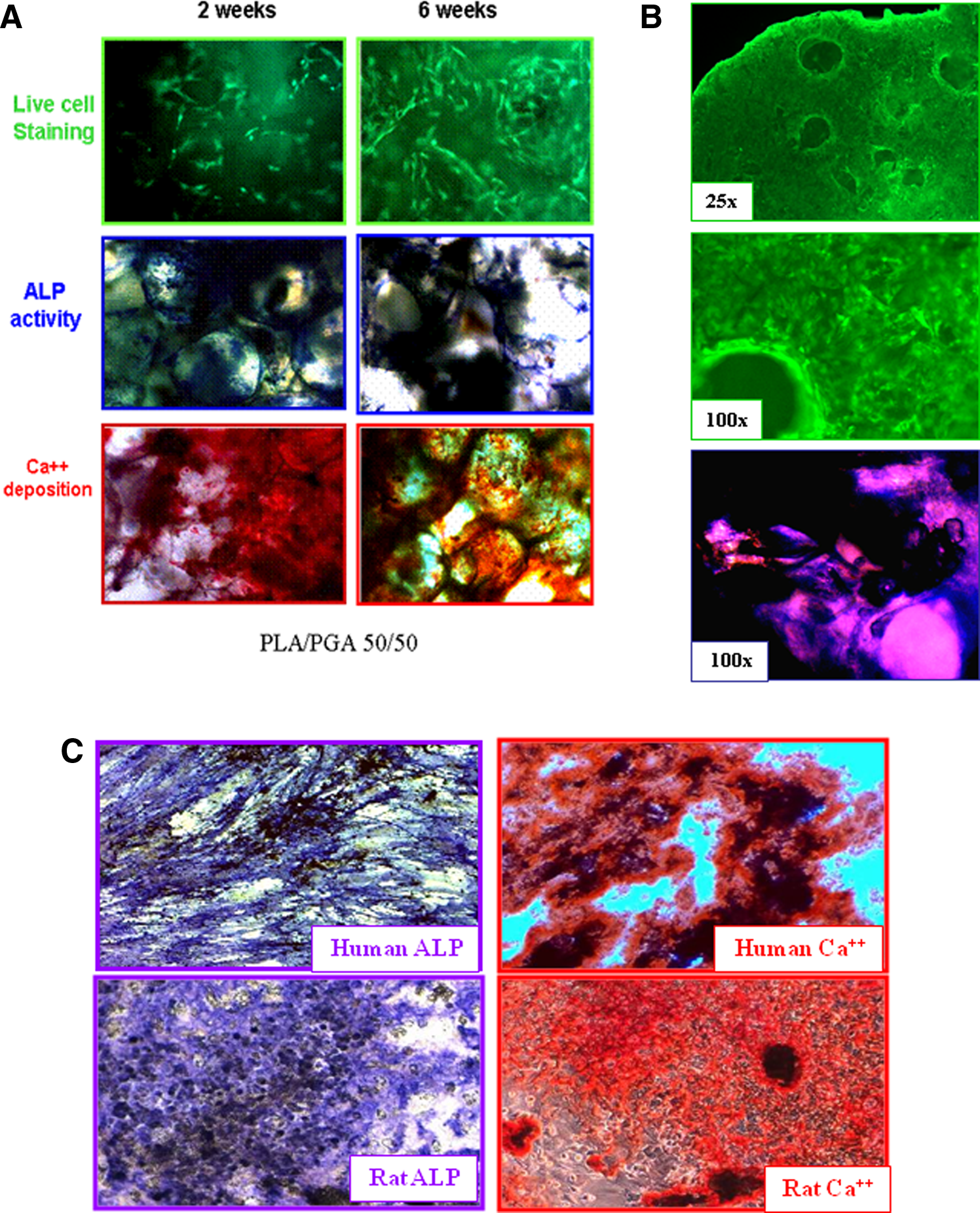
Assessment of osteogenic differentiation of human and rat BPCs.
The osteogenic status of BPCs growing either on plastic or on scaffolds was also demonstrated on a gene level. All BPC samples were cultured for 2 weeks from the moment of plating of unprocessed BM in an OM. For comparison, MSCs derived from the same batches of BM were also differentiated in the OM for 2 weeks, although this time period might not be optimal for maximum expression of the studied genes. Both BPCs and differentiated MSCs had similar patterns of expression of osteogenic markers, although some of the genes (e.g., ALP) were upregulated to the higher extent in BPCs, whereas others genes such as bone sialoprotein (BSP) and matrix Gla protein (MGP) had higher levels of expression in MSCs differentiated for 2 weeks (Fig. 4A–F). The levels of expression of all tested genes were many folds higher in BPCs than in undifferentiated MSCs or in MSCs differentiated for 1 week. The fold increase of each gene over undifferentiated MSCs varied from batch to batch. A representative of three experiments is shown in Figure 4. The error bars show variability of triplicates of the same RNA sample. Expression of the hyaluronan-mediated motility receptor (RHAMM) decreased many folds in MSCs upon differentiation and also in BPCs. (Fig. 4G, H).

Osteogenic gene expression in human BPCs and in differentiating MSCs.
Characterization of rat BPCs
The data given in Table 1 demonstrate that while the yield of rat BPCs was significantly higher than the yield of human BPCs, their ALP activity and Ca2+ deposition were of the same order (also illustrated by microphotographs in Fig. 3C). Similar to human BPCs, rat BPCs populated the scaffolds. Rat BPCs seeded on translucent scaffolds are visualized with Calcein-AM (Fig. 3B, two upper images). Histochemistry staining demonstrated high ALP activity (Fig. 3B, bottom image).
Characterization of Human and Rat Bone Progenitor Cells Derived from Intact Bone Marrow
BPC, bone progenitor cell; BM, bone marrow; ALP, alkaline phosphatase.
Effect of rat BPCs on bone repair in vivo
According to the Alamar Blue assay, the number of BPCs on scaffolds used for transplantation was 95,140±8,955 for PLA/PGA (75/25); 71,607±14,003 for PLA/PGA (50/50); and 86,269±4,172 for PLA/ceramic scaffolds (n=5). As illustrated in Figures 5B and C, significant bone fill was seen in the right defect area in the majority of animals, 6 weeks after transplantation with PLA/PGA 50/50 and PLA/PGA 75/25 scaffolds.

Transplantation of rat BPCs in calvarial defects. Representative radiographs (top) and corresponding histological sections of the defect area (bottom). R, right side transplanted with a scaffold seeded with cells; L, left side transplanted with the same scaffold with no cells. The numbers in the images represent the identification numbers of the animals. Red circles identify the bone area used for the OD calibration of Faxitron; OD of the no bone area (black) was calibrated on the empty X-Ray film (not shown).
Scattered calcified tissue on the left side is seen at the edge of the defect and not in the center. Defects transplanted with PLA/ceramic scaffolds had a radio-opaque material on both sides due to the presence of beta-tricalcium phosphate in the scaffolds (Fig. 5D). No bone formation was observed in control animals (Fig. 5A), confirming that the defect of this size could not been naturally healed (critical-sized defect). The X-ray results are summarized in Figure 6A. There was a statistically significant difference in the amount of bone in the defects transplanted with cells and those transplanted with a scaffold alone for both PLA/PGA 75/25 and 50/50 scaffolds, although somewhat less bone formation was observed in PLA/PGA 50/50 scaffolds; however, one animal demonstrated complete closure of the defect (animal 1, Fig. 5C). The results of histomorphometry (see examples of histological sections corresponding to X-ray images in Fig. 5) are summarized in Figure 6B. Histological examination of the defects transplanted with PLA/ceramic scaffolds demonstrated a significant difference between the defects transplanted with BPCs and the defects transplanted with the scaffold alone. The difference between the defects transplanted with PLA/PGA 50/50 scaffolds seeded with BPCs and those without cells was not statistically significant due to the high variability between the animals.

Summarized results of BPC transplantation.
Discussion
It has been hypothesized that adult stem cells are cells that emerge during development and then are somehow conserved in the adult organism for tissue/organ maintenance and repair.11,12 If such conservation occurs at various stages of development, then the resulting stem/progenitor cells should differ in the degree of maturity and differentiation potential. Heterogeneity of stem cells in the BM is reflected by the fact that various methods of isolation of BM-derived stromal cells yield stem cells with a different phenotype and differentiation potential.13–17 Perhaps, none of the current methods of MSCs isolation and expansion 18 targets all BM stem cell resources. The most common method of isolation of MSCs from BM relies on the ability of MSCs to adhere to plastic. However, adherent cells are not homogenous and represent different precursor populations. 19 For example, although osteogenic differentiation considered the default lineage of MSCs, only 60% of colonies obtained through adhesion selection form bone after transplantation in vivo. 20 Humoral and intercellular signals within the BM microenvironment, such as vascular cells, pericytes, and hematopoietic stem cells, constituting a stem cell niche, control fate decisions of the mesenchymal progenitors.21–23 Destruction of this niche during MSC isolation might result in lower yields and suboptimal differentiation potential of isolated stem and progenitor cells.
On the basis of this knowledge, we attempted to produce predifferentiated osteoprogenitors from unprocessed BM bypassing isolation of MSCs. Human BM aspirates were incubated for 2 weeks with a commonly used OM, except no FCS, serum substitutes, or growth factors were added, because responding stem and/or progenitor cells were present in the BM milieu. Indeed, these culture conditions were more favorable for proliferation of stem/osteoprogenitor cells than culture conditions commonly used for derivation and expansion of MSCs, achieving higher yields of BPCs. Besides biological factors mentioned above, dexamethasone present in the OM could also have a mitogenic effect. 24
Differentiation and seeding of BPCs on a scaffold occur simultaneously in a simple rotational bioreactor system. 25 Based on the results shown in Figure 2A, the yield of BPCs on scaffolds could be estimated as 100,000–150,000 cells per 1 mL BM. Because BPCs were produced by incubation in the OM for 2 weeks, their osteogenic status was compared to that of MSCs differentiated in a conventional OM for 2 weeks. MSCs were derived from the same batches of BM. BPCs produced by the new method were compatible with osteoprogenitors derived from MSCs as demonstrated by surface expression of ALP, ALP activity, Ca2+ deposition, and the levels of expression of osteogenic genes. Two genes known to be expressed during osteogenesis ALP and BSP 26 were upregulated many folds in both BPCs and differentiated MSCs, although the absolute levels of upregulation varied as the time of their maximal expression was not optimized, but rather standardized to 2 weeks. Two other genes, MGP, a calcium-binding protein similar to osteocalcin, 27 and RHAMM, a hyaluronan receptor similar to the main hyaluronan receptor, CD44, 28 were studied in our laboratory as potential new markers of osteogenic differentiation. Indeed, MGP was upregulated up to 50-fold upon differentiation of MSCs. A 10-fold upregulation of MGP was also demonstrated in BPCs. Surprisingly, expression of RHAMM drastically decreased upon osteogenic differentiation of MSCs. Similarly, RHAMM expression in BPCs was many folds lower than in undifferentiated MSCs.
Osteogenic potential of BPCs seeded on scaffolds was tested in vivo in an autologous critical-sized calvarial defect model. 29 Quantitative radiographic (Faxitron) analysis and histomorphometry demonstrated larger amount of bone in the defects transplanted with scaffolds seeded with BPCs compared to scaffolds without cells. No direct comparison with MSCs was made because of the variability of the methods for making tissue-engineering grafts seeded with MSCs. Cell-plating time, seeding density, and preseeding or postseeding differentiation influence the bone repair efficacy of such constructs, making their comparison to BPC-seeded scaffolds unreliable. However, the percentage of a new bone (76.1%±18.3%) in the defects transplanted with PLA/PGA scaffolds seeded with about 100,000 BPCs was compatible to or even higher than in published studies of bone repair by MSCs in the calvarial model in mice (50,000 MSCs transplanted on PLA/PGA scaffolds 30 or 107 MSCs in platelet-rich plasma [PRP] 31 ), in rats (500,000 MSCs in agarose 32 or umbilical cord blood-derived predifferentiated MSCs mixed with a demineralized bone matrix 33 or 300,000 MSCs in chitosan 34 ), and in rabbits, where 2×107 MSCs were transplanted in PRP gel. In the latter study, MSCs predifferentiated with dexamethasone produced better results than undifferentiated MSCs. 35
The amount of bone observed by X-ray evaluation was higher than shown by the histological analysis, when compared on a per-animal basis. This could be explained by several factors: (1) the X-ray quantitation refers to the whole defect, whereas the histology refers only to a 5-μm-thick band roughly at the center of the defect. Therefore, any radio-opaque substance at the lower or upper parts of the defect (as seen on the radiograph) will not be measured by the histomorphometry. This inherent discrepancy will result in underestimation of the mineralized material by the histomorphometry and a better, more global assessment by the radiographic analysis; (2) on the other hand, the radiographic analysis does not discriminate between new bone formed as part of the healing process, bone chips that were left within the defect during surgery, and mineralized scaffolds (all 3 forms were found in the histological sections). Therefore, the radiographic data will usually give an overestimation of the amount of new bone. The histological inspection is the only means to ascertain the formation of true new bone, and thus a healing process. Combination of the two methods yielded a quite accurate report of the outcome of the animal studies and demonstrated that BPCs produced by direct osteogenic differentiation of intact BM are capable of generation of a new bone in vivo.
Footnotes
Acknowledgments
The experimental work presented in the article was performed while I.G., N.A., and M.S. were working at Teva Pharmaceutical Industries, Petach Tikva, Israel. The authors want to thank Teva administration for their financial support of the study.
Author Disclosure Statement
No competing financial interests exist.