
Abstract
Purinergic signaling may represent an effective target in cancer therapy because the expression of purinergic receptors is altered in many forms of cancer and extracellular nucleotides modulate cancer cell growth. We examined the effect of extracellular ATP on the growth of the metastatic breast carcinoma cell line MDA-MB-435 relative to an immortalized breast epithelial cell line, hTERT-HME1. We also investigated whether the metastasis suppressor gene BRMS1 alters the sensitivity of breast cancer cells to ATP. Exposure to ATP for 24 h decreased proliferation and induced apoptosis in hTERT-HME1. However, exposure to ATP did not decrease proliferation or induce apoptosis in MDA-MD-435 cells until 48 h of exposure and only at higher doses than were effective with hTERT-HME1, suggesting MDA-MB-435 cells were resistant to the antiproliferative and apoptosis-inducing effects of ATP. Exposure to ATP for 24 h induced a decrease in proliferation of MDA-MB-435 cells expressing BRMS1, similar to hTERT-HME1, but did not induce an increase in apoptosis. MDA-MB-435 cells expressed low levels of the purinergic receptor P2Y2, as well as decreased ATP-induced cytosolic calcium mobilization, relative to hTERT-HME1. However, expressing BRMS1 in MDA-MB-435 cells restored P2Y2 levels and ATP-induced cytosolic calcium mobilization such that they were similar to hTERT-HME1. These data suggest that BRMS1 increases the sensitivity of breast cancer cells to the antiproliferative, but not apoptosis-inducing effects of ATP and that this is at least partly mediated by increased expression of the P2Y2 receptor.
Introduction
In addition to serving as an intracellular energy store, ATP also operates as an extracellular signaling molecule. 1 Many cell types release ATP into the extracellular environment where it acts as an autocrine or paracrine factor by activating two classes of cell surface receptors. P2X purinergic receptors form nonselective cation channels, while P2Y receptors are metabotropic G-protein–coupled receptors. 2 The activation of these receptors by ATP and other extracellular nucleotides induces a diverse set of cellular responses in normal tissues and cells, including but not limited to proliferation, 3 migration, 4 and apoptosis. 5
Accumulating evidence suggests that purinergic signaling may be important in the pathophysiology of many types of cancer and a valuable target in the development of cancer therapies. Alterations in the expression profile of purinergic receptors has been noted in leukemia 6 and pancreatic cancer 7 as well as other cancers. Additionally, extracellular nucleotides have been shown to modulate cancer cell growth.8,9 In normal breast tissue, the activation of P2 receptors regulates fluid secretion by mammary epithelial cells. 10 Since the growth of many epithelial cell types has been shown to be regulated by extracellular nucleotides,11,12 it is likely that ATP also regulates mammary epithelial and breast cancer cell proliferation.
MDA-MB-435 (435) is a human metastatic breast carcinoma cell line. Breast-cancer metastasis suppressor 1 (BRMS1) is a gene that reduces cancer cell metastatic capacity and is widely expressed in almost all normal human tissues, 13 but is absent in metastatic cancer cells such as MDA-MB-435. Interestingly, MDA-MB-435 cells that have been engineered to express BRMS1 (435-BRMS1) are significantly less metastatic to lungs and regional lymph nodes. 13 BRMS1 also blocks lung and regional lymph node metastases in experimental melanoma and ovarian cancer models.13–17 While the exact mechanisms by which BRMS1 inhibits metastasis remains unknown, 18 it is now recognized that BRMS1 acts as a transcriptional repressor via an interaction with histone deacetylase complexes. 19 In particular, BRMS1 suppresses the activity of nuclear factor (NF)-κB 20 and expression of anti-apoptotic genes. 21 These findings led us to hypothesize that BRMS1 expression would increase the sensitivity of breast cancer cells to ATP-induced growth inhibition and apoptosis. In this study, we examined the effect of extracellular ATP on the growth of the metastatic breast carcinoma cell line MDA-MD-435 and whether expression of affects sensitivity to ATP.
Methods
MDA-MB-435 cells were maintained in Dulbecco's modified Eagle's medium (DMEM):F12 media (Invitrogen) supplemented with 5% fetal bovine serum (Hyclone) and 1% penicillin/streptomycin. 435-BRMS1 cells, transfected with the expression vector pcDNA3 containing a BRMS1 cDNA 13 and 435-pcDNA3 (from Dr. Danny Welch, Department of Cancer Biology, University of Kansas Medical Center), containing an empty vector, were maintained in the same media supplemented with 500 μg/mL geneticin. hTERT-HME1, an immortalized breast epithelial cell line, was utilized as a normal control and maintained in mammary epithelial growth medium (Clonetics) supplemented with 52 μg/mL bovine pituitary extract, 0.5 μg/mL hydrocortisone, 10 ng/mL hEGF, 5 μg/mL insulin, and 50 μg/mL gentamicin.
The effect of ATP on cellular proliferation was assessed using the 3-(4,5-dimethyl-2-thiazolyl)-2,5diphenyl-2H tetrazolium bromide (MTT) assay (Roche). hTERT-HME1, MDA-MD-435, and transfected 435 cell lines were seeded to 24-well plates at 1.5×104 cells/cm2 and cultured for 24 h. Cells were treated with media containing 0, 10, 100, or 1000 μM ATP for 24 or 48 h, with media being replaced after the first 24 h. Because of the action of nucleotidases, the half-life of ATP is on the order of seconds. 22 Therefore, it is necessary to add relatively high concentrations of ATP under experimental conditions to expose cells to physiologically relevant concentrations of ATP.
To assess proliferation, ATP-containing media was removed and replaced with 600 μL of fresh growth media containing 60 μL of MTT reagent. In viable, metabolically active cells, MTT is converted to an insoluble, purple formazan product, the absorbance of which is directly proportional to cell number. Cells were labeled with MTT reagent for 4 h at 37°C. An equal volume of sodium dodecyl sulfate (SDS) solubilizing solution was then added to each well and plates incubated at 37°C for an additional 2.5 h. Two hundred microliters of each sample was then transferred to a 96-well plate in duplicate and the absorbance measured at 560 nm (Dynex). Results are expressed relative to vehicle controls.
The effect of ATP on apoptosis was assessed via fluorescent-activated cell sorting (FACS). Cells were treated with ATP for 24 or 48 h, washed with phosphate-buffered saline and collected in 0.25% trypsin. Cells (1×105) were suspended in 0.1 M HEPES (pH 7.4), 1.4 M NaCl, and 25 mM CaCl2, and stained with Annexin V-FITC (BD Pharmingen) for 15 min. Cells that lost plasma membrane asymmetry and were stained by Annexin V-FITC were quantified by FACS (FACScan, BD Pharmingen). Gating was set up using fluorescence intensity and set to exclude 99% of an unstained cell population.
For Western blots, cell lysates were collected in 0.1% Triton X-100, 10 mM Tris, pH 8, and 1 mM EDTA, supplemented with a protease inhibitor cocktail (Calbiochem). Twenty-five micrograms of protein was resolved by SDS-PAGE (Gradipore) and transferred to polyvinylidene difluoride membranes (Bio-Rad) and probed with antibodies specific for P2Y2 and P2X7 (Alamone Laboratories) and GAPDH (Accurate Chemical and Scientific Corp.). Immunoreactive bands were visualized by enhanced chemiluminescence detection (Amersham Biosciences) and densitometric analysis carried out with Quality One image analysis software (Bio-Rad). Data from optical density readings of protein were normalized to GAPDH.
Intracellular calcium concentration ([Ca2+]i) was determined using the ratiometric calcium indicator Fura-2 (Molecular Probes). hTERT-HME1, MDA-MB-435, and 435 transfected cell lines were cultured for 24 h in 35-mm dishes containing glass cover-slips. On the day of the experiment, cells were incubated in a 10 μM Fura-2 solution for 30 min at 37°C and then positioned on an inverted fluorescent microscope (Nikon). Metafluor image analysis software (Universal Imaging) was used to compare consecutive images at 340 and 380 nm recorded once every 2 sec to a calibration curve provided by the manufacturer. Basal [Ca2+]i levels were sampled for 60 sec at which point cells were treated with 10 μM ATP, UTP, ADP, UDP, or vehicle controls and [Ca2+]i was monitored for an additional 60 sec.
Statistical analysis
All studies were replicated in two to four independent experiments with an n=4–10. Data are expressed as means±standard error (SE). One-way analysis of variance (ANOVA) and Dunnett's multiple comparisons tests were used to compare multiple groups (Prism; GraphPad Software); p<0.05 was considered statistically significant.
Results
Extracellular ATP inhibits the proliferation of metastatic breast cancer cells
Treating hTERT-HME11 cells with 10, 100, or 1000 μM ATP for 24 h significantly decreased cell proliferation by 11.1±0.9%, 31.7±2.2%, and 41.3±2.6%, respectively, compared to untreated controls (Fig. 1). After 48 h of ATP treatment, hTERT-HME1 proliferation was further decreased by 25.0±2.9%, 68.9±1.9%, and 78.6±1.2% compared to untreated controls. In MDA-MB-435 cells, 24 h of ATP treatment had no effect on cellular proliferation. However, we observed a dose-dependent decrease in cellular proliferation after 48 h of culture. Whereas 10 μM ATP did not significantly affect MDA-MB-435 cell proliferation when compared to untreated controls, 100 and 1000 μM ATP decreased cell proliferation 29.3±3.2% and 36.4±1.1%, respectively.

Effect of ATP on cell growth. Normal human mammary epithelial cells (hTERT-HME1) or metastatic breast cancer cells (MDA-MB-435) were treated with 0, 10, 100, or 1000 μM ATP for 24 or 48 h and cellular proliferation was assessed by 3-(4,5-dimethyl-2-thiazolyl)-2,5diphenyl-2H tetrazolium bromide (MTT) assay. While hTERT-HME1 cell growth was inhibited by all doses of ATP after 24 and 48 h, ATP had no effect on MDA-MB-435 cell growth after 24 h. ATP at concentrations of 100 and 1000 μM significantly decreased MDA-MB-435 cell proliferation after 48 h. n=9–10, **p<0.01 compared to 0 μM ATP.
BRMS1 increases the sensitivity of breast cancer cells to ATP
To examine the effect of the metastasis suppressor gene BRMS1 on the sensitivity of breast cancer cells to ATP-induced growth suppression, we treated MDA-MB-435 cells expressing BRMS1 (435-BRMS1) 13 or vector control (435-pcDNA3) with ATP for 24 or 48 h. Like the parental cell line, cellular proliferation was not affected by ATP after 24 h in 435-pcDNA3 cells (Fig. 2), but 100 and 1000 μM ATP decreased cell proliferation after 48 h by 22.8±3.9% and 37.4±2.7%, respectively. Interestingly, we observed a significant decrease in 435-BRMS1 cell growth after 24 h. ATP at 10, 100, and 1000 μM decreased 435-BRMS1 cell proliferation by 13.8±1.9%, 33.7±2.5%, and 43.2±3.7%, respectively. Cell growth was similarly depressed after 48 h of ATP treatment.

Effect of BRMS1 expression on ATP-induced growth suppression. MDA-MB-435 cells transfected with an expression vector containing a cDNA of BRMS1 (435-BRMS1) or empty vector control (435-pcDNA3) were treated with 0, 10, 100, or 1000 μM ATP for 24 or 48 h and cellular proliferation assessed by MTT assay. Twenty-four hours of ATP treatment did not inhibit 435-pcDNA3 proliferation, but 100 and 1000 μM ATP decreased proliferation after 48 h. In cells expressing BRMS1, ATP significantly decreased cell proliferation after 24 and 48 h. n=9, **p<0.01 compared to 0 μM ATP.
ATP induces breast cancer cell apoptosis
To examine whether the decrease in cell growth following ATP treatment results from an increase in apoptosis, cells were treated with ATP for 24 or 48 h; stained with Annexin V-FITC, which indicated a loss of plasma membrane asymmetry; and assessed via FACS analysis. Treating hTERT-HME1 cells with 100 or 1000 μM ATP for 24 h increased Annexin V staining 2.0±0.1-fold and 2.6±0.3-fold, respectively, compared to untreated controls (Fig. 3A). However, we did not observe a significant effect of ATP on Annexin V staining in any of the breast cancer cell lines. After 48 h Annexin V staining was increased 3.2±0.2-fold and 3.3±0.5-fold by 100 and 1000 μM ATP, respectively, in hTERT-HME1 cells compared to untreated controls (Fig. 3B). Similar increases were also observed in MDA-MB-435 and 435-pcDNA3 cells. In cells expressing BRMS1, Annexin V staining was increased 0.9±0.2-fold and 1.0±0.1-fold after treatment with 100 and 1000 μM ATP, respectively. Although the increase was significant, it was rather modest and this result was likely due to higher basal apoptosis levels in cells expressing BRMS1 (Fig. 3C).

Effect of ATP on mammary epithelial cell and metastatic breast cancer cell apoptosis. Cells were treated with the 0, 10, 100, or 1000 μM ATP for 24 h
BRMS1 rescues expression of the P2Y2 receptor in breast cancer cells
To determine why cell growth is suppressed by 10 μM ATP in normal mammary epithelial cells and breast cancer cells expressing BRMS1, but not others, we examined the expression of the P2Y2 and P2X7 receptor in these cell lines. By Western blot, we detected the expression of both the P2Y2 and P2X7 receptor in hTERT-HME1 cells (Fig. 4A). In MDA-MB-435 and 435-pcDNA3 cells, we detected only a small amount of P2Y2 (19.4±2.8% and 22.2±3.6% of hTERT-HME1, respectively). However, expressing BRMS1 in MDA-MB-435 cells rescued the expression of the P2Y2 receptor (86.6±9.6% of hTERT-HME1). Levels of the P2X7 receptor were similar in all four cell types.
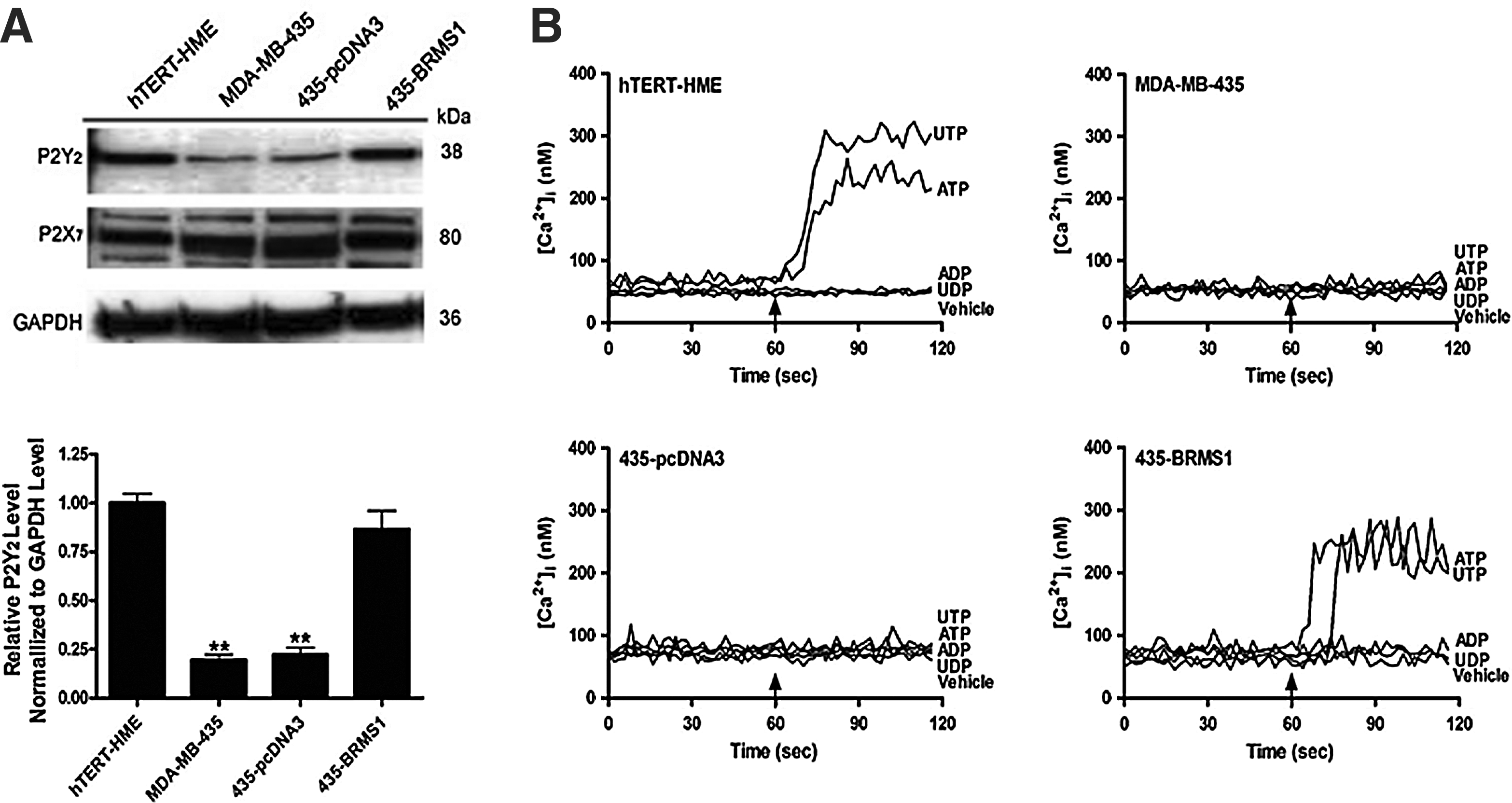
Expression of P2 receptor expression in mammary epithelial cells and effect of nucleotides on intracellular calcium.
To test the functionality of the P2Y2 receptor, we treated these cells with extracellular nucleotides (10 μM) and examined changes in [Ca2+]i (Fig. 4B). Both ATP and UTP increased [Ca2+]i in hTERT-HME1 cells, but had no effect on [Ca2+]i in MDA-MB-435 or 435-pcDNA3. In cells expressing BRMS1, ATP and UTP were able to increase [Ca2+]i, which is consistent with the higher levels of P2Y2 expression. ADP and UDP did not increase [Ca2+]i in any of the four cell types.
Discussion
Purinergic signaling represents a valuable target in the treatment of many forms of cancer. Changes in the expression patterns of P2 receptors in cancer cells have been noted6,7 as has the effect of extracellular nucleotides on cancer cell growth.8,9 The goal of this study was to examine the effect of the metastasis suppressor gene BRMS1 on the sensitivity of breast cancer cells to ATP. In two models of human breast cancer, MCF-7 and MDA-MB-231, ATP suppresses cancer cell growth, 23 but the effect of BRMS1 expression has not been examined. Previous studies indicate that BRMS1, identified via differential display as a metastasis suppressor gene, 13 acts as a transcriptional repressor. 19 For example, BRMS1 interacts with histone deacetylase complexes to inhibit NF-κB activity, thereby suppressing expression of osteopontin, 20 a matrix protein associated with tumor progression, and anti-apoptotic genes including cIAP2 and Bfl-1/A1. 21 Therefore, we hypothesized that BRMS1 expression would enhance sensitivity of breast cancer cells to ATP.
We are unaware of any previous study that has examined the effect of ATP on normal breast epithelial cells. Using hTERT-HME1 as a model for normal mammary epithelial cells, we demonstrated that 24 h of ATP treatment at 10 to 1000 μM significantly decreased epithelial cell growth compared to untreated controls. Furthermore, the decrease in cell growth at 100 and 1000 μM ATP, respectively, was accompanied by a significant increase in apoptosis.
Interestingly, MDA-MB-435 cells were more resistant, relative to hTERT-HME1 cells, to ATP-induced growth suppression. Twenty-four hours of ATP treatment was without effect on breast cancer cell proliferation, and only 100 and 1000 μM ATP decreased proliferation after 48 h. The ATP- induced inhibition of proliferation of breast cancer cells is consistent with previous studies suggesting that ATP decreases cancer cell proliferation. 24 However, it is important to emphasize that MDA-MB-435 cells were more resistant to ATP inhibition of proliferation, relative to normal breast epithelial cells, consistent with their aggressive phenotype.
The MTT assay is an established method of testing the cytotoxicity of many cancer drugs. Unlike other cell count assays, it focuses on cell viability since its readout depends on metabolic activity. A possible limitation of using the MTT assay in this study is that cancer cells both in vivo and in vitro favor glycolysis to a greater extent than their nonmalignant counterparts—a consequence of the Warburg effect. 25 However, in our experiments, the cancer cells acted as their own controls and different ATP doses represented the experimental variable. Therefore, this difference in metabolism between cancer cells and normal cells would not alter interpretation of our results. In addition, since the MTT assay depends on the reducing factors NADH and NADPH, which are generated to a lesser degree by glycolysis- favoring cells, 26 we speculate that this method is more likely to underestimate the number of cancer cells compared to normal cells. This in turn would actually amplify the difference we observed between normal and cancer cells.
Consistent with our hypothesis, expressing BRMS1 in MDA-MB-435 cells increased their sensitivity to ATP. Similar to normal epithelial cells, ATP at 10–1000 μM significantly decreased cell proliferation after 24 h. Unexpectedly, in 435-BRMS1 cells 24 h of ATP treatment did not increase apoptosis. We had expected ATP to increase 435-BRMS1 cell apoptosis since BRMS1 suppresses expression of anti-apoptotic genes. 21 This apparent discrepancy could be related to the higher basal levels of apoptosis in these cells.
The mechanism responsible for decreased ATP-induced apoptosis in breast cancer cells, including those expressing BRMS1, was not investigated in this study. However, we found that expressing BRMS1 in MDA-MB-435 cells restores P2Y2 expression. That it is a functional channel is evidenced by the calcium imaging data presented. These data also demonstrated that the cytosolic calcium response to ATP is intact in the absence of sensitivity to ATP-induced apoptosis. Taken together, this suggests that decreased sensitivity to ATP-induced apoptosis is downstream or independent of P2Y2 mediated increases in [Ca2+]i.
The increased sensitivity of 435-BRMS1 cells to ATP-induced growth suppression is likely due to expression of the P2Y2 receptor. Compared to normal mammary epithelial cells, MDA-MB-435 cells expressed very little P2Y2, but expressing BRMS1 in these cells rescued the expression of this receptor. This result is consistent with the finding that lower concentrations of ATP inhibited growth of hTERT-HME1 and 435-BRMS1 cells relative to MDA-MB-435 and 435-pcDNA3 cells. Whereas 10 μM ATP is sufficient to activate the P2Y2 receptor, higher concentrations of ATP are necessary to activate the P2X7 receptor (100 μM), which was expressed by all four cell types. 2 The observation that 10 μM UTP, which also activates the P2Y2 receptor, increased [Ca2+]i and also suppressed cell growth (data not shown) in hTERT-HME1 and 435-BRMS1 cells, but not MDA-MB-435 or 435-pcDNA3 cells, also supports this hypothesis.
While the exact molecular mechanisms by which BRMS1 suppresses metastasis remains elusive, 18 it is possible that BRMS1 alters the ability of cancer cells to persist in metastatic tissue. Tissues, such as bone and lung, which are preferred metastatic targets for migratory breast cancer cells, release nucleotides in response to a number of stimuli.27,28 Our data suggest that ATP released from these tissues could inhibit growth of breast cancer cells still expressing BRMS1 via activation of the P2Y2 receptor, whereas cells that no longer express BRMS1 would be resistant to low concentrations of ATP. Consistent with this hypothesis, MDA-MB-435 cells and MDA-MB-231 cells stably transfected with BRMS1 still form locally invasive tumors, but fail to colonize other tissues. 13 In this regard, in the context of our data, the characteristic of the host tissue to release nucleotides would represent an endogenous defense by the host tissue against metastasizing cells. Further exploration of this phenomenon could add to the inventory of possible mechanisms that the BRMS1 gene uses to exert its metastasis-suppressing powers in vivo.
It is important to note that the mammary origin of MDA-MB-435 cells has been called into question, with some data suggesting that MDA-MB-435 cells are actually melanocytes.29,30 However, MDA-MB-435 cells have been demonstrated to express milk proteins and metastasize from mammary fat pad but not subcutaneous tissue, consistent with the cell line being a breast carcinoma.31,32 Regardless of the issue of the authenticity of MDA-MB-435 cells as breast cancer cells, the interpretation of the data suggesting that BRMS1 sensitizes cancer cells to ATP-induced growth suppression remains relevant.
In summary, we demonstrated that metastatic breast cancer MDA-MB-435 cells are resistant to ATP-induced growth suppression, while the expression of BRMS1 sensitizes MDA-MB-435 cells to ATP at least partly by altering the expression of the purinergic receptor P2Y2. These data suggest that agonists for this receptor could be used to inhibit the growth of breast cancer cells prior to the onset of metastasis, while agonists of the P2X7 receptor could be used to induce breast cancer cell apoptosis.
Footnotes
Acknowledgments
This work was supported by National Institutes of Health, National Cancer Institute grant CA90991, and the Michael and Myrtle Baker Orthopaedic Research Endowment.
Author Disclosure Statement
No competing financial interests exist.