
Abstract
An origin of Earth life on Mars would resolve significant inconsistencies between the inferred history of life and Earth's geologic history. Life as we know it utilizes amino acids, nucleic acids, and lipids for the metabolic, informational, and compartment-forming subsystems of a cell. Such building blocks may have formed simultaneously from cyanosulfidic chemical precursors in a planetary surface scenario involving ultraviolet light, wet-dry cycling, and volcanism. On the inferred water world of early Earth, such an origin would have been limited to volcanic island hotspots. A cyanosulfidic origin of life could have taken place on Mars via photoredox chemistry, facilitated by orders-of-magnitude more sub-aerial crust than early Earth, and an earlier transition to oxidative conditions that could have been involved in final fixation of the genetic code. Meteoritic bombardment may have generated transient habitable environments and ejected and transferred life to Earth. Ongoing and future missions to Mars offer an unprecedented opportunity to confirm or refute evidence consistent with a cyanosulfidic origin of life on Mars, search for evidence of ancient life, and constrain the evolution of Mars' oxidation state over time. We should seek to prove or refute a martian origin for life on Earth alongside other possibilities.
1. Introduction
Life as we know it utilizes amino acids, nucleic acids, and lipids for the metabolic, informational, and compartment-forming subsystems of a cell. In one current hypothesis for the origin of life, these building blocks may have formed simultaneously from cyanosulfidic chemical precursors in a planetary surface scenario involving ultraviolet light, wet-dry cycling, and volcanism (Patel et al., 2015; Damer and Deamer, 2019; Sasselov et al., 2020). This process can be driven by photoredox chemistry with sulfite (SO3 2-) mediating cycling of ferrocyanide ([FeII(CN)6]4-) and ferricyanide ([FeIII(CN)6]3-) in combination with ultraviolet irradiation (Xu et al., 2018). While this scenario does not rule out other models such as an origin of life at seafloor vents (Martin et al., 2008), it plausibly and simultaneously addresses key challenges including formation and concentration of organic building blocks, their polymerization to yield functional molecules, and compartmentalization to yield protocellular entities (Joyce and Szostak, 2018).
All life as we know it shares a common ancestor (Pace, 2001). The most conserved genome regions occur within genes encoding the translation machinery (16S and 23S ribosomal subunits, transfer RNAs), which are themselves RNA machines involved in translating RNAs to polypeptides via the genetic code (Isenbarger et al., 2008). These regions have changed little over 4 billion years (Bray et al., 2018). The deep evolutionary conservation of these molecular fossils is one piece of evidence for an RNA-protein world preceding the DNA world (Hsiao et al., 2009; Fournier et al., 2010; Goldman et al., 2010; Harish and Caetano-Anollés, 2012). Furthermore, RNA molecules are capable of both storing hereditary information and catalyzing reactions, a dual role that may have been critical before the emergence of translation and the fixation of the genetic code. Protocell-like growth and division (Budin and Szostak, 2011), for example, mediated by feedstock supply and/or photochemical processes (Zhu et al., 2012; Szostak, 2017), could facilitate compartmentalization, selection, and evolution in the context of an RNA-protein world.
Genetic evidence suggests that the last universal common ancestor (LUCA), which shares many features with modern life and was evolutionarily distant from its origin (Cantine and Fournier, 2018), inhabited an anoxic, “geochemically active environment rich in H2, CO2 and iron” (Weiss et al., 2016). However, this setting on its own does not distinguish between seafloor vents and shallow-water hydrothermal habitats, nor between Earth and Mars.
Life as we know it utilizes dehydration synthesis to form the metabolic (protein, carbohydrate), informational (nucleic acid), and compartment-forming (lipid) polymers (Kitadai and Maruyama, 2018). Driving forces for dehydration include evaporation, sublimation, crystallization, or formation of hydrated minerals. Surface conditions thus offer plausible mechanisms to concentrate prebiotic molecules and produce polymers. Dehydration could possibly occur due to nanoconfinement (Muñoz-Santiburcio and Marx, 2017) in metal sulfides at alkaline vents, yet, at present, high water activity does not seem consistent with an origin of life (Frenkel-Pinter et al., 2021). Thus, while some considerations favor a cyanosulfidic model, other models, including an origin of life at seafloor vents, are not ruled out. Here we focus on the potential for evaluating a possible cyanosulfidic origin of life on Mars.
2. Problems with an Origin of Life on Earth
2.1. Early Earth was an ocean world
The cyanosulfidic origin theory is compelling, yet current data suggests that early Earth was a water world (Fig. 1A), with little to no sub-aerial continental crust before 3.5 Ga (Bada and Korenaga, 2018), reaching 1–2% land by 3.0 Ga and 5–8% by 2.5 Ga (Flament et al., 2013; Lalonde and Konhauser, 2015). Consistent with these findings, recent analysis suggests initiation of continental weathering between 3 and 2.5 Ga (Johnson and Wing, 2020). This would have limited the land area suitable for a cyanosulfidic origin of life to regions such as volcanic island hot spots (Bada and Korenaga, 2018).
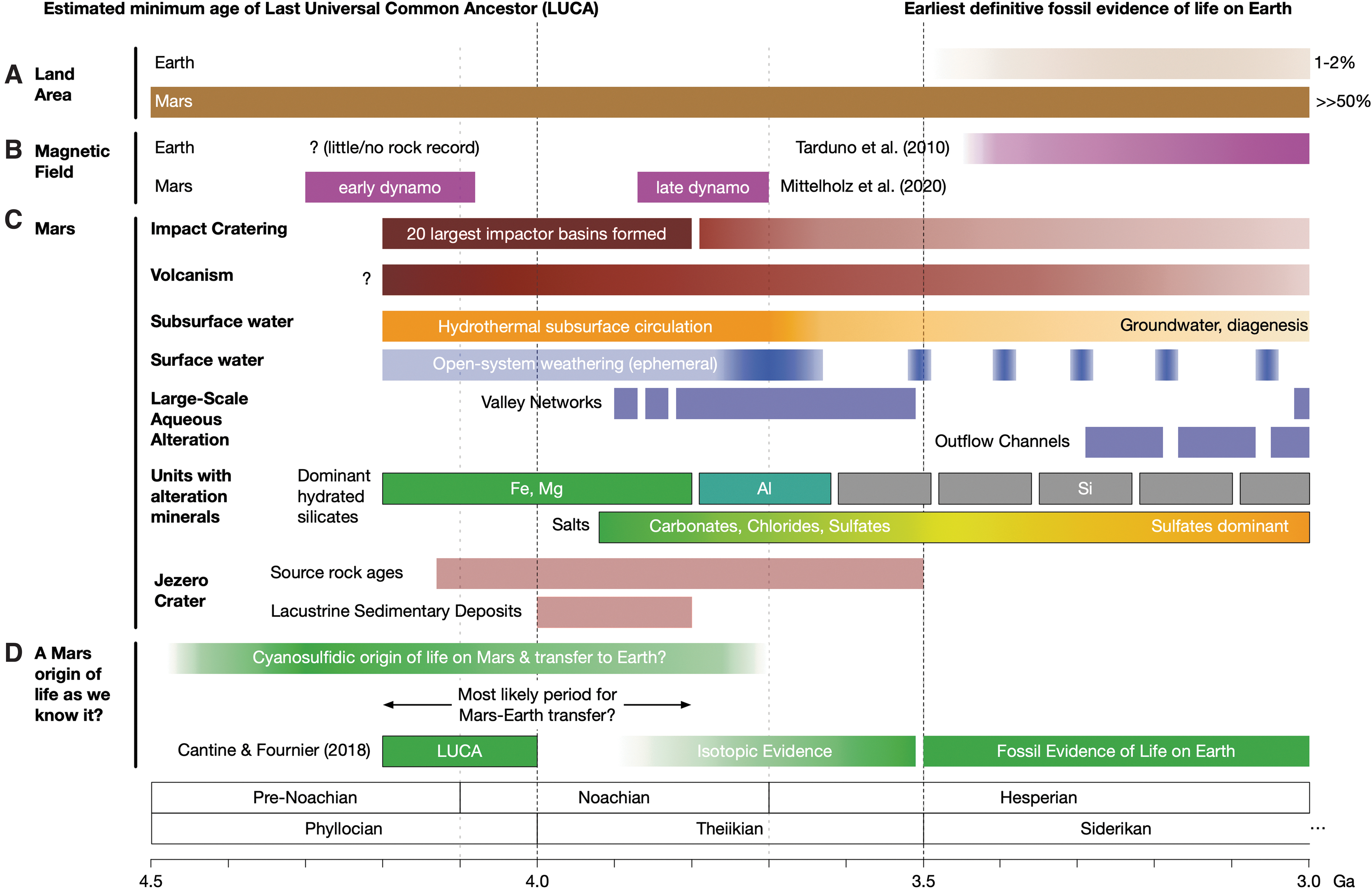
Planetary context for a hypothesized cyanosulfidic origin of life on Mars and its transport to Earth via lithopanspermia. The horizonal axis for all panels is billions of years ago (Ga). (
In contrast, the land area on early Mars was orders of magnitude greater than on early Earth. Estimates of the water inventory on Mars at the Noachian/Hesperian boundary range from tens of meters (Carr and Head, 2015) to ∼1 km global equivalent layer (Kurokawa et al., 2014). The upper bound assumes a putative northern ocean after formation of the martian dichotomy, for example, the Borealis basin resulting from a massive impact event (Andrews-Hanna et al., 2008) at or before 4.2 Ga and possibly as old as 4.5 Ga (Mittelholz et al., 2020). Even in the most extreme scenarios, Mars would have had orders-of-magnitude more land area than early Earth.
A possible resolution for a cyanosulfidic origin of life on early Earth is that closed-basin lakes in volcanic island hot spots might represent ideal locales for concentration of cyanide salts (Toner and Catling, 2019). In these regions, chemical weathering of mafic/ultramafic rocks could produce alkaline lakes and lead to ferricyanide salt precipitation. Under this scenario, favorable conditions could lead to a rapid boot-up of life despite limited land area. However, similar conditions were likely present on early Mars and potentially orders of magnitude more common. Thus, if cyanosulfidic life is easy to start up, then a cyanosulfidic origin would be all the more likely on Mars and might have occurred earlier, all other factors being equal. While the rock record of early Earth has been largely lost, on Mars this record remains, making it feasible to seek direct evidence of cyanosulfidic chemistry in ancient rocks there.
2.2. The atmospheric rise of oxygen on Earth took place more than a billion years after fixation of the genetic code
Additional support for a martian origin of life on Earth comes from amino acids and the evolution of the genetic code itself. Granold et al. (2018) proposed that the genetic code used a simpler set of amino acids and that the final diversification of amino acids happened in response to oxygen, suggesting that the diversification was late, in other words, coincident with the appearance of early local oxygen on Earth. It this hypothesis is correct, it would imply exposure to oxidizing conditions on early Earth.
However, genomic, fossil, and geological evidence suggests that fixation of the genetic code and oxidation on early Earth are separated in time by more than 1 billion years. The geologic record has revealed that Earth was largely devoid of oxygen for its first 2+ billion years, and appreciable quantities only accumulated after cyanobacteria invented oxygenic photosynthesis, resulting in the great oxidation event (GOE) (Harel et al., 2015). While the GOE occurred around 2.33 Ga (Luo et al., 2016), evidence for early local oxidative weathering suggests there could have been transient local oxygen pulses at or before 3 Ga (Crowe et al., 2013; Mukhopadhyay et al., 2014). Nevertheless, the late diversification is problematic because the root of the archaeal phylum Euryarchaeota, which arose after LUCA, has been dated to >4 Ga (Wolfe and Fournier, 2018) (Fig. 1D), and the genetic code must have been fully established before this time (Fournier and Alm, 2015; Weiss et al., 2016), including use of selenocysteine by LUCA (Weiss et al., 2016). Time-calibrated phylogenomics extending this deep into life's history comes with caveats and wide error bars, yet it highlights a >1.5 billion-year inconsistency. Thus, the timing of the fixation of the genetic code does not align with the inferred oxidation state of early Earth. However, on Mars, surface conditions became oxidizing much earlier, as discussed later in Section 3.3.
3. The Potential Origin and Evolution of Life As We Know It on Mars
3.1. A cyanosulfidic origin of life on Mars?
Today Mars is dry and cold (Savijärvi et al., 2019), yet early Mars was habitable for life as we know it, with significantly more water than is present today (Ehlmann et al., 2011; Kurokawa et al., 2014; Alsaeed and Jakosky, 2019). Availability of the main elements used by life (C, H, N, O, P, and S) and energy sources including variable redox states of iron and sulfur minerals are recorded in the stratigraphy of Gale Crater (Grotzinger et al., 2015; Hurowitz et al., 2017). Widespread clay minerals confirm extensive periods of subsurface water-rock interactions before 3.7 Ga (Fig. 1C) yet also suggest even early Mars had mostly cold and relatively arid surface conditions (Ehlmann et al., 2011), which could have aided accumulation of organic molecules through concentration and low hydrolysis rates.
Sasselov et al. (2020) delineated a plausible pathway for a cyanosulfidic origin of life on early Mars. They suggest that igneous intrusions, volcanism, or impacts interacting with cyanide salt deposits could have generated the relevant feedstocks to produce nucleotide, amino acid, and lipid precursors. Here, we also suggest a Mars cyanosulfidic origin of life could have seeded life on Earth, resolving the inconsistencies previously noted for an origin of Earth life on Earth.
Lightning would have provided hydrogen cyanide (HCN), representing a total fixed nitrogen budget on par with that of early Earth (Segura, 2005). Meteorite impacts (Fig. 1C) may also have generated HCN and provided phosphate (Pasek and Lauretta, 2005). Sulfite would likely have been available on early Mars as a consequence of volcanic SO2, a CO2 atmosphere, and low temperatures (Marion et al., 2013); later oxidation would have led to the formation of sulfate minerals, consistent with remote sensing (Flahaut et al., 2010) and in situ measurements (Rapin et al., 2019). The presence of sulfites would also help explain the relative dearth of carbonates on Mars (Marion et al., 2013).
3.2. An RNA world on Mars?
A cyanosulfidic origin of life (Patel et al., 2015; Sasselov et al., 2020) would produce all the building blocks required for an RNA-protein world (Goldman et al., 2010; Harish and Caetano-Anollés, 2012). What we are still lacking is knowledge about whether ancient Mars, especially before the Noachian-Hesperian boundary (Fig. 1), was conducive to the formation, stabilization, and evolution of an RNA-protein world and ultimately cellular life.
Hints are beginning to emerge: Mojarro et al. (2020) evaluated plausible divalent cations (Fe2+, Mg2+, and Mn2+) at different pH conditions and found the lowest rate of metal-catalyzed hydrolysis of RNA for Mg2+ at pH 5.4, rather more acidic than the circumneutral conditions suggested by the cyanosulfidic model (Sasselov et al., 2020). Bray et al. (2018) found that Fe2+ and Mn2+ can replace the modern role of Mg2+ for ribosomal RNA folding and translation in anoxic conditions, relevant to early Earth or early Mars. An improved understanding of Mars' redox history is necessary to understand the potential origin and evolution of an RNA world on Mars.
For all known life, DNA ultimately replaced RNA as the carrier of hereditary information. However, if an RNA-protein world evolved on Mars, we may still be able to sample fossilized or extant traces of it in the subsurface, or may find, like all known Earth life, that its role in heredity has been displaced by DNA.
3.3. Mars surface redox evolution occurred earlier than on Earth
While an oxidizing atmosphere on Earth arose due to biological production of oxygen, oxidation on Mars is thought to have resulted from crustal water sequestration (Tuff et al., 2013; Wade et al., 2017) as well as photolysis of water and subsequent loss of hydrogen (Hartman and McKay, 1995; Kurokawa et al., 2014; Alsaeed and Jakosky, 2019; Fedorova et al., 2020). The present-day surface of Mars is highly oxidizing, and this state has likely persisted for billions of years as inferred from manganese hydroxides identified at Gale and Endeavor Craters (Noda et al., 2019).
On Mars, the transition to more oxidizing conditions is recorded by the deposition of sulfates (SO4 2- with oxidation state S6+) from approximately the Noachian-Hesperian transition (3.7 Ga) onward (Fukushi et al., 2019; Rapin et al., 2019) (Fig. 1C). This is consistent with orbital observations of Noachian-aged Fe3+-bearing clay minerals (e.g., associated with surface oxidation) and in situ detection of their purported ferrous smectite precursors (Chemtob et al., 2017).
As the full range of sulfate deposits on Mars has yet to be identified, oxidation may also have occurred earlier. Recent modeling work demonstrates that Mars may have transitioned between reducing and oxidizing conditions repeatedly early in its history (Wordsworth et al., 2021). Furthermore, the relative absence of pre-Hesperian sulfate may be explained by a combination of dissolution under reducing conditions, as well as widespread thermochemical sulfate reduction due to bolide impacts, which declined over time (Wordsworth et al., 2021).
Thus, the inferred timing of surface oxidation on Mars is consistent with a potential role of oxidation in shaping fixation of the genetic code, followed by meteoritic transfer of life to Earth. This resolves the problematic interpretation (Granold et al., 2018) that the genetic code was not fixed until 1.5 Ga after the timing predicted by genetic evidence (Fournier and Alm, 2015; Weiss et al., 2016; Wolfe and Fournier, 2018) and is consistent with the antiquity of antioxidant enzymes and recent work suggesting a potential role for reactive sulfur species in their evolution (Neubeck and Freund, 2020).
3.4. Phosphate availability was likely higher on early Mars than on early Earth
Early nucleic acid–based life would have required phosphate to build informational polymers and store energy. Release rates for phosphate-bearing minerals are estimated to be 45 times higher for Mars compared to Earth, with ∼2 × higher equilibrium phosphate concentrations (Adcock et al., 2013). In contrast, phosphorus availability was limited on early Earth due to a dearth of oxidizing power that limited recycling until the GOE (Kipp and Stueken, 2017). Cyanide can promote release of phosphate from iron phosphates, enabling generation of organophosphates under desiccating conditions (Burcar et al., 2019); dry conditions were widespread on early Mars but lacking on early Earth.
3.5. Ejecta from large impact events could have transferred viable life from Mars to Earth
The plausibility of lithopanspermia has been theoretically and experimentally tested, including non-sterilizing ejection from planetary surfaces as measured in martian meteorite ALH84001 (<40°C based on residual magnetization) (Weiss et al., 2000) and through experimental studies of microbial survival to hypervelocity shock pressures (Horneck et al., 2008). Modeling suggests that the mass of Mars to Earth viable transfers (Mileikowsky et al., 2000; Worth et al., 2013) is greater than 1012 kg (Worth et al., 2013), and this ignores potential transfers before 3.5 Ga when cratering rates were higher (Fig. 1C). In comparison to the present, the thicker atmosphere (∼0.5 bar at ∼4 Ga) (Kurokawa et al., 2018) on early Mars would have provided substantial shielding. In addition, recent data supporting a martian dynamo at 4.5 and 3.7 Ga (Mittelholz et al., 2020) (Fig. 1B) suggests any life being ejected before 3.7 Ga would have been afforded additional protection from low-energy solar and cosmic radiation while on Mars (direct effect) and could have helped to slow the loss of the atmosphere to space (indirect effect).
Organisms arriving on Earth—putative chemolithoautotrophs—would have adapted to available redox gradients, including hydrothermal systems. While thermophily near the base of the tree of life was originally interpreted to provide evidence of the vent origin model, it can also be explained by parallel adaptation to high temperatures (Boussau et al., 2008), such as selection for heat-resistant species during Mars-Earth meteoritic transfer, or via subsequent heating of Earth's oceans during impact events. Reconstructed optimal growth temperature (OGT) across the tree of life suggests hyperthermophily at the base of the Archaeal and Bacterial domains but a colder OGT for LUCA of ∼45°C (confidence interval 33–68°C) (Groussin et al., 2013), consistent with non-sterilizing temperatures during meteoritic ejection (Weiss et al., 2000). Under this scenario, the transfer of life to Earth (Fig. 1D) need not have taken place before LUCA; if multiple transfers occurred, then LUCA may have existed on Mars. Indeed, large meteorite impacts may have transported life either direction between the planets until the geologically recent past (Worth et al., 2013).
4. A Martian Origin for Life on Earth Is Consistent with Genetic and Geological Evidence
What we know about early Mars is consistent with the cyanosulfidic origin of life, and conditions there would have provided orders of magnitude more land area for prebiotic chemistry to cross the threshold to life. The Noachian-Hesperian transition and commensurate changes in surface redox balance on Mars are also consistent with the timing of fixation of the genetic code (Fournier et al., 2010; Fournier and Alm, 2015) and the antiquity of antioxidant enzymes (Neubeck and Freund, 2020). Theoretical (Mileikowsky et al., 2000; Worth et al., 2013) and experimental (Weiss et al., 2000; Horneck et al., 2008) studies of lithopanspermia imply that Earth, Mars, and Venus would have shared viable microbes (Worth et al., 2013), if present in ejecta, and subsequent genetic evidence is consistent with this timeline.
Other options are possible (McKay, 2010): If cyanosulfidic chemistry and the transition to life is a planetary phenomenon (Sasselov et al., 2020), relatively fast and easy given the right conditions, then we may find that such life evolved independently on both early Mars and early Earth, despite the latter's dearth of sub-aerial crust. If booting up life is harder or rare, this would raise the prior that we may share common ancestry with any life on Mars. Finding no life on Mars would be incredibly revealing, because it would suggest life is a rare event requiring very particular circumstances.
Given the inconsistencies between the inferred history of life as we know it and Earth's geologic history, it is plausible that life on Earth could have originated on Mars (McKay, 2010). Such a history, while incredible, is a story of our past that is consistent with the available genetic and geological evidence. It is also testable.
5. Ongoing and Future Missions Should Search for Life As We Know It
While the Mars 2020 Perseverance rover will focus on habitability and evidence of ancient, and not extant, life (Williford et al., 2018), it can seek to confirm or refute conditions consistent with a cyanosulfidic origin of life (Sasselov et al., 2020), as well as seek chemical precursors to life and identify high-abundance organics.
Jezero Crater is a 45 km diameter impact crater that once harbored an open-basin lake system recorded by sedimentary deposits, deltas, and other features associated with fluvial activity that may have ended by around the Noachian-Hesperian boundary or earlier (Goudge et al., 2018). Analysis of the western fan deposit at the crater inlet has identified clay minerals (Fe/Mg-smectite) and stratigraphy that likely records an extended history of early Mars, drawn from a watershed much larger than the crater, which includes smectites, olivine, and Mg-rich carbonate-bearing terrains, among others (Goudge et al., 2015). Beyond likely having been habitable itself, Jezero Lake and the associated watershed should provide access to samples originating in a diverse set of other potentially habitable environments, and the lacustrine setting implies access to a well-ordered stratigraphic record of the Noachian period on Mars.
Because the fluvial activity associated with lacustrine sedimentary deposits at Jezero Crater likely ended by 3.8 Ga, evidence within those deposits of increasing oxidation over time would be consistent with the ancestral relationship scenario. Alternatively, a relatively reducing surface environment up to 3.8 Ga would not rule out life as we know it on Mars but would weaken the case for any ancestral relationship, at least one linked to the Jezero Crater source region. For comparison, sedimentary rocks within Gale Crater, which formed between 3.6 and 3.8 Ga, record a mix of oxidation states even within the base of the stratigraphic section (Grotzinger et al., 2014), as well as evidence of redox stratification in younger deposits (Hurowitz et al., 2017), consistent with an oxidative surface environment by the Noachian-Hesperian boundary or early Hesperian.
If putative ancient life on Mars was exposed to an oxidizing surface environment before the earliest evidence of life on Earth, a Mars-Earth transfer is not ruled out. If no part of Mars became oxidizing until significantly after the earliest signs of life on Earth (<3.7 Ga), or certainly after the earliest unambiguous evidence of fossil life on Earth (3.5 Ga) (Schopf, 1993; Schopf et al., 2018), then the window of opportunity to complete fixation of the genetic code on an oxidized Mars prior to a hypothesized Mars-Earth transit would close and lead to rejection of the hypothesis that oxidizing conditions on Mars could explain fixation of the genetic code. However, plausible cycling between reducing and oxidizing conditions on early Mars (Wordsworth et al., 2021) would have provided ancient life such an adaptive opportunity.
Silica has high preservation potential, and hydrated silica deposits have been identified within Jezero Crater in association with delta deposits (Tarnas et al., 2019). Organic materials can be concentrated locally and stabilized by adsorbing to silica and the aforementioned clays (Fe/Mg-smectite). Lacustrine carbonates in Jezero Crater also offer high preservation potential (Horgan et al., 2020). A study of amino acid preservation in simulated martian conditions found the highest preservation in smectites and sulfates (dos Santos et al., 2016).
Extrapolation from modeling of hydrolysis and ancient DNA sequencing projects (Allentoft et al., 2012; Millar and Lambert, 2013) suggests that 100 bp DNA at -60°C could have a half-life >2 billion years. However, other factors such as naturally occurring radioisotope decay may limit the decay half-life to perhaps 10 million years (Kminek et al., 2003). RNA generally has higher hydrolysis rates, although the pH dependence is different (Mojarro et al., 2020). Thus, direct detection of remnant nucleic acid polymers in Noachian-aged terrain is extremely improbable. However, nucleobases, the informational component of nucleic acids, have been identified in meteorites (Callahan et al., 2011); thus, at least under cold conditions, nucleobases can survive >4 billion years.
Intact nucleic acids and nucleobases can adsorb to silica, and in the relative absence of later aqueous and thermal alterations, nucleobases can be retained. Silica is, in fact, used to isolate cross-linked RNA and proteins to study their interactions (Asencio et al., 2018). Cross-linking would be expected as part of the diagenesis process, as would be deamination of cytosine to yield uracil (Dabney et al., 2013). Mars 2020's instruments are not designed to specifically interrogate uracil, but it could be targeted in returned samples along with other nucleobases.
Lipids are also one of the most geologically stable biomolecules and are used as a biosignature in studies of Earth life in rocks older than 2 Ga (French et al., 2015). Studies of Mars-relevant diagenesis reveal caveats. Tan and Sephton (2020) found that hydrous pyrolysis from 200°C to 280°C for 72 h significantly degraded lipids, especially with low carbon content, high water-to-rock ratios, and iron-rich minerals (Tan and Sephton, 2020). Carrizo et al. (2020) demonstrated the potential to detect lipid biomarkers using a tunable Raman laser spectrometer and found that of silica-rich versus iron-rich samples tested, the silica-rich material enhanced preservation. Thus, silica-rich deposits in Jezero Crater could be targeted in the search for nucleobases and lipids.
Many of the mineral deposits mentioned are expected to be accessible to the Mars 2020 rover, for example, within the stratigraphy of the western delta deposits. Because Mars lacks a substantial atmosphere, organics within deposits exposed at the surface would be damaged over time by space radiation. To avoid this damage, low exposure ages are desirable. Mudstone on the floor of Gale Crater was found to have a exposure age of 78 ± 30 million years (Farley et al., 2014), highlighting the potential to utilize wind-driven geomorphological change to obtain samples with potential for complex organic molecules as undegraded by cosmic rays as possible. Thus, it may be feasible to use a similar approach in Jezero Crater, leveraging prior wind analysis (Day and Dorn, 2019) and in situ measurements (Mars Environmental Dynamics Analyzer [MEDA]) to assess access to low exposure age stratigraphy. Because accessible exposure ages may still be large, drilling, including, in future missions, below the 1–2 m penetration depth of space radiation, remains critical for accessing undegraded samples. Notably, the ExoMars Rosalind Franklin rover includes a drill capable of reaching 2 m; its planned landing site, Oxia Planum, offers another opportunity to seek evidence of cyanosulfidic chemistry, with rocks dating from 3.6–4.0 Ga and evidence of past aqueous activity (Gary-Bicas and Rogers, 2021; Quantin-Nataf et al., 2021).
While several Mars 2020 rover instruments (SuperCam [Wiens et al., 2017], SHERLOC [Beegle et al., 2013], PIXL [Allwood et al., 2015]) will be used to detect or infer the presence of organics and specific chemical moieties, they may lack the ability to uniquely identify specific molecules. The Mars Organic Molecule Analyser (MOMA) instrument on the Rosalind Franklin rover may enable detection, if present, of organics with chain or ring-based structures, and will characterize molecular weight distribution patterns and chirality (Goesmann et al., 2017). Instruments in development could one day target nucleic acids (Carr et al., 2016; Mojarro et al., 2019) and polypeptides (Ohshiro et al., 2014; Carr et al., 2020).
6. Conclusions
Early Mars likely offered more opportunity for cyanosulfidic prebiotic chemistry to cross the threshold to life than early Earth, including cold temperatures to stabilize accumulation of organics and orders-of-magnitude more land area. Mars' earlier transition to oxidizing conditions could have played a role in fixation of the genetic code on a timeline consistent with genetic evidence. Furthermore, all steps in the meteoritic transfer of life from Mars to Earth have been theoretically or experimentally verified, and such a transfer is consistent with genetic evidence. Mars 2020 and the planned Rosalind Franklin rover are well positioned to test the plausibility of a cyanosulfidic origin of life on Mars, search for ancient evidence of life, and constrain the evolution of Mars' oxidation state over time. Unlike Earth, where ancient rocks are rare, on Mars we can access rocks across nearly the full 4.5 billion year history of that planet (Bouvier et al., 2018). Despite the modern thin (1% that of Earth) atmosphere and cold average surface temperature (-60°C), the subsurface of Mars likely remains habitable (Jones et al., 2011). Future missions, with access to special regions, including deep drilling to seek habitable environments in the subsurface, may be required to target any extant life and unambiguously determine whether life as we know it exists on Mars today and, if so, whether it is related to us.
Footnotes
Acknowledgments
I thank Maria T. Zuber and Gary Ruvkun for extensive discussions and collaborative research on this topic over the last 15 years, and the reviewers for improving the manuscript.
Author Contributions
C.E.C. conceived and wrote the paper.
Funding
C.E.C. was supported by NASA award NNX15AF85G to M.T.Z. and 80NSSC22K0188 to C.E.C.
Author Disclosure Statement
I declare no competing interests.
Associate Editor: Norman Sleep