
Abstract
Habitability has been generally defined as the capability of an environment to support life. Ecologists have been using Habitat Suitability Models (HSMs) for more than four decades to study the habitability of Earth from local to global scales. Astrobiologists have been proposing different habitability models for some time, with little integration and consistency among them, being different in function to those used by ecologists. Habitability models are not only used to determine whether environments are habitable, but they also are used to characterize what key factors are responsible for the gradual transition from low to high habitability states. Here we review and compare some of the different models used by ecologists and astrobiologists and suggest how they could be integrated into new habitability standards. Such standards will help improve the comparison and characterization of potentially habitable environments, prioritize target selections, and study correlations between habitability and biosignatures. Habitability models are the foundation of planetary habitability science, and the synergy between ecologists and astrobiologists is necessary to expand our understanding of the habitability of Earth, the Solar System, and extrasolar planets.
1. Introduction
Life on Earth is not equally distributed. There is a measurable gradient in the abundance and diversity of life, both spatially (e.g., from deserts to rain forests) and temporally (e.g., from seasonal to geological timescales). Our planet has also experienced global environmental changes from the Archean to the Anthropocene, which further conditioned life to a broad range of conditions. In general, a habitable environment is a spatial region that might support some form of life (Farmer, 2018), albeit not necessarily one with life (Cockell et al., 2012). One of the biggest problems in astrobiology is how to define and measure the habitability not only of terrestrial environments but also of planetary environments, from the Solar System to extrasolar planets (also known as exoplanets). The word habitability literally means the quality of habitat (the suffix -ity denotes a quality, state, or condition). Astrobiologists have been constructing different general definitions of habitability, not necessarily consistent with one another (e.g., Hoehler, 2007; Shock and Holland, 2007; Cárdenas et al., 2014, 2019; Cockell et al., 2016; Heller, 2020). Other more specific habitability definitions, such as the canonical Habitable Zone (i.e., the orbital region in which liquid water could exist on the surface of a planet), are used in exoplanet science (Kasting et al., 1993). Ecologists developed a standardized system for defining and measuring habitability in the early 1980s; however, this is seldom utilized in the astrobiology community (USFWS, 1980).
The popular term habitability is formally known as habitat suitability in biology. Ecologists before the 1980s were using different and conflicting measures of habitability, a situation not much different from today for astrobiologists. The U.S. Fish and Wildlife Service (USFWS) decided to solve this problem with the development of the Habitat Evaluation Procedures standards in 1974 for use in impact assessment and project planning (USFWS, 1980). These procedures include the development and application of Habitat Suitability Models (HSMs) (Hirzel and Lay, 2008). Other names for these models are Ecological Niche Models, Species Distribution Models, Habitat Distribution Models, Climate Envelope Models, Resource Selection Functions, and many other minor variants (Guisan et al., 2017). These multivariate statistical models are widely used by ecologists today to quantify species-environment relationships with data obtained from both ground and satellite observations. HSMs integrate concepts as needed from ecophysiology, niche theory, population dynamics, macroecology, biogeography, and the metabolic theory of ecology.
Astrobiologists have largely not utilized HSMs for at least three reasons. First is the naming: habitability is a common word in the earth and planetary sciences, but it is not generally used by biologists. Thus, a quick review of the scientific literature shows no definition of this concept in biological terms. The second reason is the specialization: HSMs are a specialized topic of theoretical ecology, which is not highly represented in the astrobiology community. The third is applicability: HSMs are mostly used to study the distribution of specific wild animals and plants, not microbial communities or ecosystems in general (generally the focus of astrobiological studies), so it may not seem readily applicable to the field of astrobiology, but this is changing (e.g., Treseder et al., 2012). For example, microorganisms (bacteria, fungi, and other unicellular life) exhibit endosymbiotic relationships with animals and plants and also play a key role in their survival. Thus, anything that can be said about habitability at the macroscopic level is tightly coupled to habitability at the microscopic level. Indeed, potential methods for incorporating microbial ecology into ecosystem models are discussed in the work of Treseder et al. (2012). In one way, the mathematical framework behind HSMs is easier to apply to microbial communities than animals because the spatial interactions of animals (e.g., predation) tend to be much more complex. However, microbial life is not easy to quantify in free-living populations, and it is thus harder to validate the HSMs with them, although molecular methods are changing this (Douglas, 2018).
The definition and core framework of HSMs can be extended from Earth to other planetary environments. However, the astrobiology field does not have the luxury of validating HSMs with the presence of life unless when applied to environments on Earth (e.g., extreme environments). Thus, known ecophysiology models are used instead to predict the occurrence, distribution, and abundance of putative life in any planetary environment. Attempting to measure the habitability of a system without knowing all the environmental factors controlling it may seem like an impossible task. However, even on Earth this problem can be approached by selecting a minimum set of relevant factors to simplify the characterization of the systems. While the objective can be to establish whether a system is habitable, it can also be simply to explore how the selected environmental variables contribute to the habitability of the system. Usually, a library of habitability metrics is created for each environment or life-form under consideration, with each metric dependent on the species, scales, or environmental factors under consideration. In a fundamental sense, the only way to really know whether a place is habitable is to find (or put) life on it (Zuluaga et al., 2014; Chopra and Lineweaver, 2016). It is nearly impossible, nor is it desirable, to include all factors affecting habitability in a model, even for environments on Earth. Thus, the objective of habitability models is to understand the contributions of a finite set of variables toward the potential to support a specific species or community (e.g., primary producers, organisms that use abiotic sources of energy) (Guisan et al., 2017). So even if we do not know or do not include all the relevant factors, we can consider the effects of those we do know.
Here, we recommend adapting and expanding the ecologists' nearly four decades of experience in modeling habitability on Earth to astrobiological studies. These models can be used to characterize the spatial and temporal distribution of habitable environments, identify regions of interest in the search for life, and eventually explore correlations between habitability and biosignatures. For example, such models would help test the hypothesis that biosignatures (or biomarkers) are positively correlated with proxy indicators of geologically habitable environments (or geomarkers); that is, there is life whenever there are habitable environments on Earth (Martinez-Frias et al., 2007). We also note that the concept of biosignatures encompasses any detectable signature of life or its by-products on a planet's atmosphere, which includes possible signatures of planetary-scale technology, known as technosignatures (Wright and Gelino, 2018). Measurements by past and future astrobiology-related observations (e.g., from ground, telescopes, or planetary missions) can be combined into a standard library of habitability models. Results from different observations can then be compared, even when using different measurements, since, through the use of HSMs, their results can be mapped to the same standard scale (e.g., zero for worst and one for best regions). A Habitability Readiness Analysis could be developed for any observation campaign to determine how its current instruments could be used, or what new instruments should be added, for habitability measurements in the spatial and temporal habitability scales of interest. Furthermore, it might also be possible to develop new instruments for direct habitability measurements.
This review addresses many of the misconceptions about habitability and stresses the need for better integration between the habitability models used by ecologists and astrobiologists. This is not a review of those factors affecting habitability, which are discussed elsewhere (e.g., National Research Council, 2007; Des Marais et al., 2008; McKay, 2014; Hendrix et al., 2018), but about the multivariate models that integrate these factors. Section 2 presents an overview of current ecology models with an emphasis on the HSM. Section 3 discusses some examples of how habitability is currently implemented in the astrobiology field. Section 4 describes our recommendations on how to adapt and expand the ecology models to the astrobiology field. Section 5 presents important science questions that could be answered from habitability models. Finally, Section 6 presents our concluding remarks.
2. Habitability in Ecology
Habitat Suitability Models (HSMs) are widely used in ecology to study the habitability of environments, many times under different definitions: Species Distribution Models or Ecological Niche Models (Kuhn et al., 2016; Guisan et al., 2017). An important step in the construction of HSMs is the selection of spatially explicit environmental variables at the right resolution to determine a species' preferred environment (i.e., its niche) as close to its ecophysiological requirements as possible. Environmental variables (such as edaphic, from the Greek noun “edaphos” meaning ground factors—defined as any chemical, physical and biological properties of the soil) can exert complex direct or indirect effects on species (e.g., Oren, 1999, 2001; Fierer et al., 2007; Allison and Martiny, 2008; Lauber et al., 2008; Rajakaruna and Boyd, 2008; Fierer et al., 2012). These variables are ideally chosen to reflect the three main types of influence on a species: (1) regulators or limiting factors, defined as factors controlling a species' metabolism (e.g., physical-chemical conditions such as temperature and salinity); (2) disturbances, defined as any perturbations affecting environmental systems; and (3) resources, defined as all compounds that can be consumed by organisms (e.g., nutrients). There are many other variables that exert an indirect, rather than a direct, effect on species distribution. The construction of HSMs follows five general steps: (1) conceptualization; (2) data preparation; (3) model calibration; (4) model evaluation; and (5) spatial predictions (Guisan et al., 2017).
One of the main HSM tools is the Habitat Suitability Index (HSI), which provides one way of quantifying the capacity of a given habitat to support a selected species. An index is the ratio of a value of interest divided by a standard of comparison. The value of interest is an estimate or measure of the quality of habitat conditions for a species in the studied environment, and the standard of comparison is the corresponding value for the optimum habitat conditions for the same evaluated species. An HSI of zero (minimum value) represents a totally unsuitable habitat, and a maximum value of one represents an optimum habitat. In developing an HSI, we should obtain a direct and linear relationship between the HSI value and the carrying capacity of the environment for the species under consideration (USFWS, 1980). The functions describing the species distribution or abundance along each environmental variable in an HSM are called species response curves (Austin and Gaywood, 1994). These curves, when plotted, can vary from simple boxlike envelopes resulting in binary indices to more gradual and complex responses resulting in continuous indices (Fig. 1).
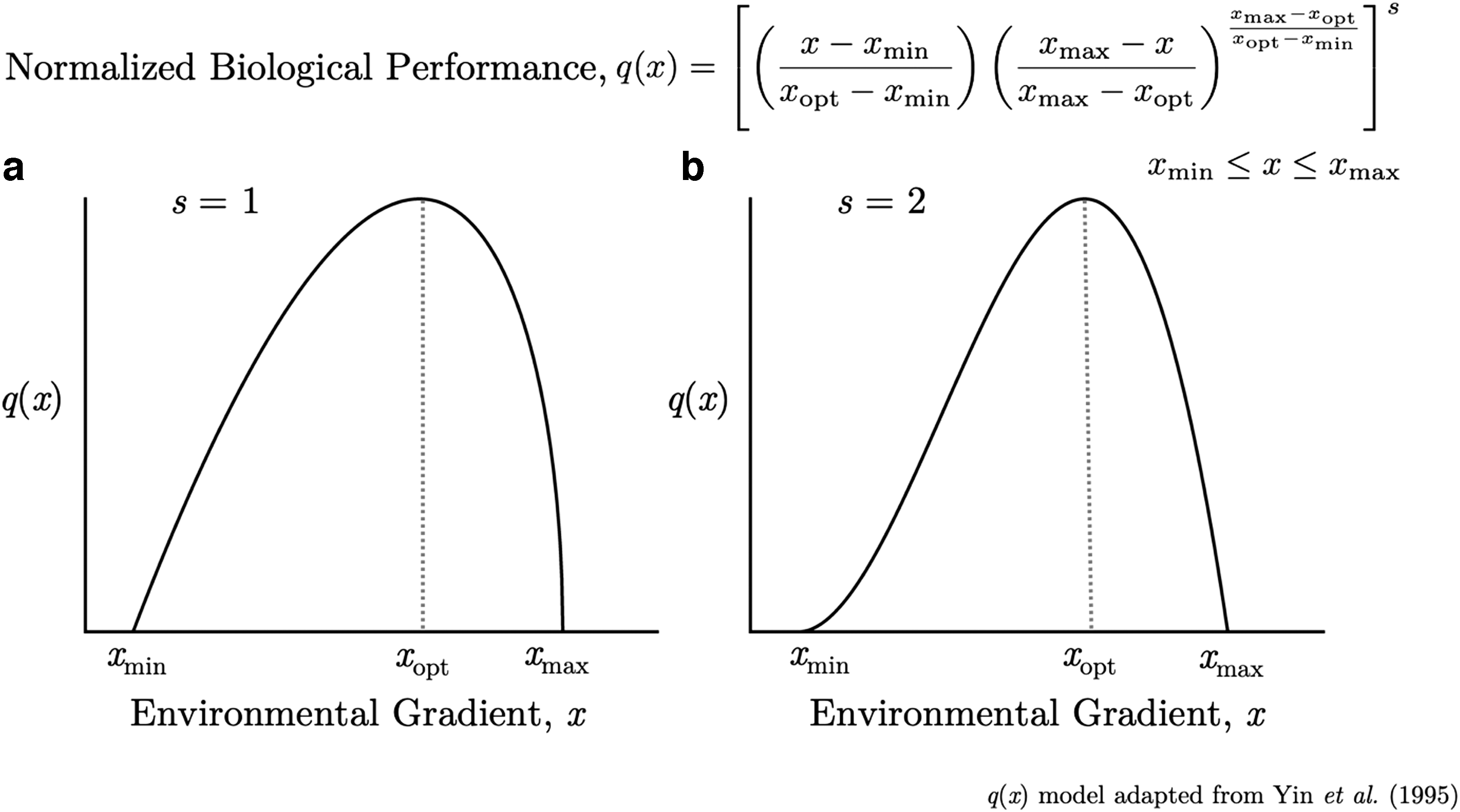
Typical shapes of ecophysiological or species response curves along an environmental gradient (e.g., temperature) for the biological performance (e.g., relative growth rate) of plants (
Carrying capacity is generally defined as the maximum supported population density in equilibrium. More precisely, carrying capacity is the user-specified quality biomass of a particular species for which a particular area will supply all energetic and physiological requirements over a long, but specified, period (Giles, 1978). Since habitability could be taken as proportional to carrying capacity, as defined by the HSI, it is then related to the fraction of mass (e.g., nutrients) and energy (e.g., light) available or usable by a particular species or community from the environment. A common and difficult problem for HSIs is how to combine the effect of many environmental variables into a single index. In theoretical ecology, the solutions are known as aggregation methods. These methods can combine the variables by using arithmetic, geometric, or harmonic means, among others. The general rule is to keep the index proportional to carrying capacity and correlated with the presence and absence of the species of interest in the environment. Occurrences or presence probabilities are generally simpler to combine as products. Ecophysiological response curves often involve the fitting of standard statistical models to ecological data by using simple (multiple) regression, Generalized Linear Models, Generalized Least Squares, or Generalized Additive Models, among others.
The usual approach is to create a library of HSI models for all species (or communities) and environments under consideration, each with its own particular limitations (Brooks, 1997; Roloff and Kernohan, 1999). These models are easy to compare and combine since they use the same uniform scale (e.g., a value between zero and one, proportional to the carrying capacity). Thus, each HSI is only applicable to a specific type of life and habitat as a function of a finite set of environmental variables within selected spatial and temporal scales. There are many other tools of the HSM that can be used to characterize species or their environment. For example, similarity indices are usually simpler to construct than an HSI and can be used for quick comparisons between a set of biological or physical properties (e.g., diversity) (Boyle et al., 1990). Similarity indices are also used in many other applications such as pattern recognition and machine learning (e.g., Cheng et al., 2011).
3. Habitability in Astrobiology
Astrobiologists have proposed many habitability models or indices for Earth, the Solar System, and extrasolar bodies in the last decade (e.g., Stoker et al., 2010; Schulze-Makuch et al., 2011; Armstrong et al., 2014; Irwin et al., 2014; Barnes et al., 2015; Silva et al., 2017; Kashyap Jagadeesh et al., 2017; Rodríguez-López et al., 2019, Seales and Lenardic, 2020). There are some specific universal biological quantities that can be used as proxies for habitability such as carrying capacity, growth rate, metabolic rate, productivity, or the presence of some requirements of life, or even genetic diversity (Heller, 2020). There is also an ongoing debate as to whether any concept of habitability needs to be binary (yes/no) in nature (Cockell et al., 2019), continuous (Heller, 2020), or probabilistic (Catling et al., 2018). While a binary interpretation of habitability only allows a given planet to be habitable (for a given species) or not, a continuous model also allows for the possibility of a world (planet or moon) to be even more habitable than Earth, that is, to be superhabitable (Heller and Armstrong, 2014; Schulze-Makuch et al., 2020). Constructing a direct measure of habitability requires knowing how the environment affects one of the biological quantities for some species or community. We do not need to specifically estimate these quantities, but only to know how the environment proportionally affects them. For example, we know how temperature affects the productivity of primary producers such as plants and phytoplankton. Most require temperatures between 0°C and 50°C, but such producers do better (i.e., have the highest productivity) near 25°C (Silva et al., 2017). Their “thermal habitability function” looks like a bell-shaped curve centered at their optimum productivity temperature. Direct measures of habitability are also better represented as a fraction from zero to one.
Biological productivity (the dry or carbon biomass produced over space and time) is one of the best habitability proxies for astrobiology since it is easy to estimate for many ecosystems, via ground or satellite observations. The Miami Model was the first global-scale empirical model to give fair estimates of terrestrial net primary productivity (NPP, the rate of fixed photosynthetic carbon minus the carbon used by autotrophic respiration) (Zaks et al., 2007). This simple model only uses two measurements, annual mean surface temperature and precipitation, to successfully infer the global distribution of vegetation (Adams et al., 2004). One important limitation of this type of model is that climate variables such as precipitation not only affect but also are affected by vegetation. There is increasing evidence, for example, that tropical forests have strong impacts on cloud base heights (Van Beusekom et al., 2017) and precipitation patterns on Earth (Molina et al., 2019). Today, many complex biogeochemical models and satellite observations (e.g., NASA's TERRA, AQUA, and Soumi NPP models) are combined to estimate local to global NPPs (Cramer et al., 1999; Ito, 2011). These satellite products are being used to create habitability indices to monitor terrestrial biodiversity now and through climate change (e.g., Pan et al., 2010; Radeloff et al., 2019). Therefore, the NPP is also a measure of global terrestrial health or habitability since primary producers are the basis of the food chain.
Most habitability models are limited to indirect measures of habitability due to a lack of information. This is especially true for exoplanets. For example, the occurrence of Earth-sized planets in the Habitable Zone of stars (termed the Eta-Earth value) can be considered a continuous indirect measure of stellar habitability (i.e., the suitability of stars for habitable planets). The Habitable Zone, the region around a star where an Earth-like planet could maintain surface liquid water, is generally considered to be a binary indirect measure of planetary habitability (Kasting et al., 1993), although others have argued that it should be regarded as a probability density function (Zsom, 2015; Catling et al., 2018). While the location of the Habitable Zone depends on stellar type, its extension also greatly depends on the assumed atmospheric composition (e.g., Heng, 2016). Furthermore, atmospheric dynamics effectively work to homogenize differential heating of the surface, creating a short-term response on the planet's global temperature. This differential heating is a result of the planet's obliquity, which governs the latitudinal distribution of incoming stellar radiation (Spiegel et al., 2009; Nowajewski et al., 2018). The Habitable Zone boundaries themselves also evolve over time. This has major implications for water delivery, water retention, and oxygen buildup on potentially habitable planets (e.g., Ramirez and Kaltenegger, 2014; Luger and Barnes, 2015).
The Habitable Zone can be defined in terms of either the planet's distance from the star, its incoming stellar flux, or its global equilibrium temperature. When using the equilibrium temperature definition, the extension of the Habitable Zone depends on the planet's orbital forcings, particularly eccentricity and obliquity. For example, when orbital eccentricity increases, the average equilibrium temperature decreases, thus extending the size of the Habitable Zone (Méndez and Rivera-Valentín, 2017). Similarly, higher fixed obliquity and/or rapid changes in obliquity values result in higher average equilibrium temperatures, which also result in extending the outer edge of the Habitable Zone (Armstrong et al., 2014). Furthermore, when using the equilibrium temperature definition, the extension of the Habitable Zone depends ultimately on the planet's energy balance. On Earth, the global energy balance is a result of the complex interaction between physical and biological processes. Biota affect the global energy balance in manifold ways including direct effects on surface albedo and latent heat fluxes (e.g., transpiration) (Jasechko et al., 2013; Duveiller et al., 2018). Tidal heating from the newly formed and nearby Moon might have played a role early in Earth's history (Heller et al., 2020) but is irrelevant today. On Earth-sized planets in the Habitable Zones around M dwarf stars, tidal heating can have a strong effect on the planetary energy budget, potentially making some parts of the Habitable Zone uninhabitable (Barnes et al., 2009).
The Earth Similarity Index, inspired by the diversity similarity indices used in ecology to compare populations (Boyle et al., 1990), is a measure of Earth-likeness for a selected set of planetary parameters (Schulze-Makuch et al., 2011). Future observational constraints of Earth-similar atmospheric constituents (i.e., N2, CO2, H2O) could improve our handle on this and similar metrics. For instance, 3D global climate models indicate that spectral features of water vapor on close-in terrestrial exoplanetary atmospheres may be detectable by the James Webb Space Telescope (Kopparapu et al., 2017; Chen et al., 2019), depending on the presence of clouds (Komacek et al., 2020). Even though the presence of water vapor in the atmospheres of terrestrial exoplanets can indicate habitability, it is necessary to perform exhaustive work to determine which species could survive under conditions of extreme humidity. For example, mammals are not capable of surviving hyperthermia produced under high air temperatures and high humidity conditions, so planets with extreme differential heating between latitudes may be uninhabitable for them, despite having liquid water on their surface (Nowajewski et al., 2018). Furthermore, many animals (including all mammals) may not survive in atmospheres with CO2 (N2) pressures exceeding ∼0.1 bar (Ramirez, 2020).
The current Habitable Zone paradigm is misunderstood by many people—the public, the press, as well as other scientists—but as with all habitability models, it has a specific application and is neither incorrect nor useless for neglecting the subsurface oceans in the outer Solar System, venusian clouds, or other environments far from Earth-like conditions. The Habitable Zone does not tell us whether planets are habitable (or even if there are planets there), but it shows the impact of a few important variables on planetary habitability. The concept of a Habitable Zone was developed to identify terrestrial exoplanet targets that could potentially host life. It was first proposed by Edward Maunder in 1913 (Maunder, 1913; Lorenz, 2020) in his book Life on Other Planets, with refining definitions later on (Huang, 1959; Hart, 1978; Kasting et al., 1993; Underwood et al., 2003; Selsis et al., 2007; Kaltenegger and Sasselov, 2011; Kopparapu et al., 2013, 2014; Ramirez and Kaltenegger 2017, 2018; Ramirez, 2018). The Habitable Zone can be defined as the circumstellar region where standing bodies of liquid water could be stable on the surface of a terrestrial planet. Here, the insistence on the presence of liquid surface water is based on the fact that all known examples of life on Earth require liquid water to exist. However, this definition is suitable only for remote observations of planets and does not consider any life which might exist within the subsurface. There is a reason for this: the search for life on exoplanets will rely on remote observations of atmospheres for the foreseeable future, lacking the luxury of in situ measurements used in solar system planetary science. Therefore, identifying water in the atmosphere of planets (in addition to other biosignature-relevant gases) is the only way to narrow down potential life-hosting targets, since subsurface life deep in the interior may not be able to modify the atmospheres of planets enough to be detectable remotely.
The abundance of liquid water in a planetary environment may be inherently unstable (Gorshkov et al., 2004), which leads to questions about the role of life in the definition of habitability itself (Zuluaga et al., 2014). A disequilibrium may be one the most conspicuous signatures of a habitable and inhabited planet (Lovelock, 1965, 1975; Kleidon, 2012; Krissansen-Totton et al., 2016); consider, for example, the composition of Earth's atmosphere (Lenton, 1998). One common problem with some, if not all, biological models is that they assume that the full set of physical characteristics of the environment, including climate, is a boundary condition for life, that is, that biological systems depend on climate but not the other way around. This premise is challenged by the fact that the observed state of the Earth system is the result of a complex and dynamic interaction between biological (e.g., ecosystems) and physical (e.g., climate) systems (Budyko, 1974; Gorshkov et al., 2000; Kleidon, 2012; Zuluaga et al., 2014). A critical question is how such a thermodynamically unstable state can be maintained during eons (e.g., the span of Earth's life) despite variable (e.g., solar luminosity) and sudden large (e.g., asteroid impact) external forcings . The answer depends on the interactions between biological and physical systems on Earth. A planet might enter a habitable state (i.e., allow for the presence of liquid water at its surface) at any given point in time simply through chance occurrence—a random change in planetary energy balance, for example. However, long-term persistence of a habitable state (e.g., the persistent habitable state of Earth over the last 4 billion years, approximately) indicates the existence of natural regulation mechanisms (Walker et al., 1981; Lenton, 1998; Gorshkov et al., 2000; Kleidon and Lorenz, 2004, Salazar and Poveda, 2009).
4. Recommendations for Astrobiology
The astrobiology field is playing a critical role in our understanding of planetary habitability. The habitability of Earth, the Solar System, and exoplanets can be studied thanks to measurements taken with multiple ground, orbital, or remote sensors. At the same time, astrobiology-related missions can synergistically take advantage of the predictions of habitability models in their selection of potential exploration strategies, mission priorities, instrument design, and observations and experiments. Here we list four recommendations for the astrobiology community:
5. Science Questions
Each astrobiology-related project, mission, or instrument should anticipate and answer a series of basic scientific questions about the environment or environments to be studied as a core part of the planning process. The answers to these questions should be updated based on the results. To do so, it is important to define an environment of interest, both in space and time (termed a quadrat in ecology), and answer the following science questions as part of the initial analysis:
6. Conclusion
Habitability models are successful analysis tools for characterizing habitable environments on Earth. In this review, we compared some of the different models used by ecologists and astrobiologists and suggested how to integrate them into new habitability standards. These standards are relevant for any astrobiology-related observations, including the study of extreme environments on Earth, planetary missions, or exoplanets. Ecologists have been using these models for more than four decades to understand the distribution of terrestrial life at local to global scales (Section 2). Astrobiologists have been proposing different models for some time, with little integration and consistency between them and different in function to those used by biologists (Section 3). The astrobiology community should create habitability standards for observations and missions with astrobiology objectives, as the USFWS successfully did long ago for ecologists (Section 4). These standards are necessary to make sense of data from multiple observations, develop predictions for environmental niches that can be tested, and understand the extraterrestrial correlations between habitability and biosignatures (Section 5).
There is no need for the astrobiology community to reinvent the methods and tools used by ecologists. It is true that ecology methods are more capable than our limited planetary and astronomical data allow, but they also provide the basic language and framework to connect Earth and astrobiology science for decades to come. For example, there are many theoretical and computational tools used in ecology to quantify environments and their habitability, mostly known as Habitat Suitability Models. See the work of Guisan et al. (2017) for an extensive review of these models and the work of Lortie et al. (2020) for a current review of the computational tools. Most of these tools are available as packages in the R Computing Language 3 in the Comprehensive R Archive Network (CRAN) 4 and GitHub (e.g., Environmetrics 5 , HSDM 6 ). New, higher-resolution remote sensing instruments and exploration technologies will create better habitability maps from rover, lander, and orbiter data. Habitability models will eventually lead us to a better understanding of the potential for life in the Solar System and beyond, and perhaps even the factors that influence the development of life itself. Habitability models are the foundation of planetary habitability science. After all our scientific and technological advances, we still need a stronger integration between biology, planetary sciences, and astronomy (Cockell, 2020).
Footnotes
Acknowledgments
This work was supported by a NASA Astrobiology Institute (NAI) workshop grant, the Planetary Habitability Laboratory (PHL), and the University of Puerto Rico at Arecibo (UPR Arecibo). Thanks to NASA Puerto Rico Space Grant Consortium and the Puerto Rico Louis Stokes Alliance For Minority Participation (PR-LSAMP) for supporting some of our students. Thanks to Ravi Kumar Kopparapu from the NASA Goddard Space Flight Center and James Kasting from Penn State for valuable comments. RH is supported by the German space agency (Deutsches Zentrum für Luft- und Raumfahrt) under PLATO Data Center grant 50OO1501. RMR is supported by the Earth-Life Science Institute and the National Institutes of Natural Sciences: Astrobiology Center (grant number JY310064). AA and MFS are funded by the Science and Technology Development Fund, Macau SAR. JHM gratefully acknowledges support from the NASA Exobiology program under grant 80NSSC20K0622. GG was supported by the NSF Luquillo Critical Zone Observatory (EAR-1331841) and the LTER program (DEB 1831952). JF acknowledges partial support from NASA PSTAR grant 80NSSC18K1686. The US Department of Agriculture (USDA) Forest Service's International Institute of Tropical Forestry (IITF) and UPR Río Piedras provided additional support. A portion of the research by MM was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration (80NM0018D0004). This is LPI contribution number 2596. LPI is operated by USRA under a cooperative agreement with the Science Mission Directorate of the National Aeronautics and Space Administration.
Associate Editor: Lewis Dartnell