
Abstract
This paper presents a reformulation of the submarine alkaline hydrothermal theory for the emergence of life in response to recent experimental findings. The theory views life, like other self-organizing systems in the Universe, as an inevitable outcome of particular disequilibria. In this case, the disequilibria were two: (1) in redox potential, between hydrogen plus methane with the circuit-completing electron acceptors such as nitrite, nitrate, ferric iron, and carbon dioxide, and (2) in pH gradient between an acidulous external ocean and an alkaline hydrothermal fluid. Both CO2 and CH4 were equally the ultimate sources of organic carbon, and the metal sulfides and oxyhydroxides acted as protoenzymatic catalysts. The realization, now 50 years old, that membrane-spanning gradients, rather than organic intermediates, play a vital role in life's operations calls into question the idea of “prebiotic chemistry.” It informs our own suggestion that experimentation should look to the kind of nanoengines that must have been the precursors to molecular motors—such as pyrophosphate synthetase and the like driven by these gradients—that make life work. It is these putative free energy or disequilibria converters, presumably constructed from minerals comprising the earliest inorganic membranes, that, as obstacles to vectorial ionic flows, present themselves as the candidates for future experiments. Key Words: Methanotrophy—Origin of life. Astrobiology 14, 308–343.
1. Introduction
T acted as a reservoir for Fe2+… The oceanic crust was ultramafic or komatiitic … The pyrrhotite (FeS) content was generally high and, … at a distance from the oceanic ridges, seawater, occupying cracks in this crust, would have been involved in relatively low-temperature convection cells driven by crustal heat … This convecting seawater at <200°C would have serpentinized the crust, becoming alkaline by this process of hydrolysis … Also, hydrogen would have been produced during serpentinization … and methane formed from a portion of this hydrogen and CO2 by the Fischer-Tropsch reaction with FeO in silicates. Phosphate … and reduced sulphur … would have been released from mafic rocks to these relatively low-temperature solutions. (Russell et al., 1989)
The first experiments grounded in this hypothesis showed that “fine chimneys were apparently composed of greigite (Fe3S4)” and that precipitation of the semi-permeable colloidal membrane takes place at the interface between the two fluids where the pH change is abrupt … Such a continuous membrane would have an extensive reactive surface per unit volume, … fine iron sulphide framboid sacks and chimneys could act as … culture chambers and flow reactors … for life to originate by phosphorylation and growth of organic molecules on the iron sulphide surfaces … Waste from such reactions would be carried away by the venting solutions. (Russell et al., 1989)
Since then, and particularly since the discovery of the Lost City alkaline hydrothermal vent field in 2000, which was broadly as anticipated (Russell et al., 1989; Shock, 1992; Kelley et al., 2001; Martin et al., 2008), further consideration has been given to the following:
(1) a somewhat more acidic early ocean with NO augmenting atmospheric CO2 and SO2 dissolved in the then hydrosphere, generating a steep redox gradient against hydrothermal H2 (Russell et al., 1993; Russell and Hall, 1997; Martin et al., 2007);
(2) the process of serpentinization, which is exothermic and strain-inducing and provides positive feedbacks to the hydrothermal convective drive (Kelley et al., 2001; Lowell and Rona, 2002; Russell et al., 2003, 2013; Vance et al., 2007);
(3) the fact that life's major preoccupations, both at its very start and as the basic necessities for all life today, are C1 metabolism and phosphate anhydride bond formation very far from equilibrium with its constituent phosphates (Baltscheffsky, 1971; Fuchs, 1989, 2011; Branscomb and Russell, 2013);
(4) nitrate, nitrite, and photolytic ferric iron and manganite having acted as high potential electron acceptors that enabled concomitant redox reactions (Nealson and Saffarini, 1994; Russell and Hall, 1997; Ducluzeau et al., 2009; Kopf et al., 2013);
(5) molybdenum in sulfide and/or green rust, bifurcating electrons, one exergonically and the other endergonically to drive carbon fixation (Boucherit et al., 1991; Nitschke and Russell, 2009, 2011, 2013; cf. Itaya et al., 1987; Helz et al., 2013);
(6) the ambient pH gradient of ∼5 units having the theoretical potential, as a proton motive force, to drive the pyrophosphate:orthophosphate ratio to extreme heights to fuel proto-biochemistry (Russell and Hall, 1997, 2006; Russell et al., 2013), altogether focusing the thermodynamic disequilibria at, and across, the membrane sufficient to meet the criticisms of Pascal et al. (2013) while ensuring protection of organic products from the oxidizing potential of nitrate and nitrite as they are spent on the outer surface of the membrane (cf. Sousa et al., 2013);
(7) the concentration and combinatorial interactions of products at low entropy in the confined space of the mineral compartments and more particularly in the interlayers of green rust comprising the membranes (Russell and Hall, 1997; Russell et al., 2003, 2013; Mielke et al., 2011);
(8) the complex precipitate membranes, consisting partly of iron sulfides dosed with Ni, Co, and Mo, as well as ferrous/ferric oxyhydroxides, also dosed with Ni and Mo, with potentially functional affinities to the active centers of the metalloenzymes—the green rust possibly acting as the first peristaltic free energy–converting structures and transporters prior to the emergence of transmembrane protein bundles of α-helices (Russell et al., 1994, 2013; Nitschke and Russell, 2009; McGlynn et al., 2012; Schoepp-Cothenet et al., 2012; Branscomb and Russell, 2013; Nitschke et al., 2013); and
(9) acetate, water, and nitric oxide being the first waste products (Russell and Martin, 2004; Nitschke and Russell, 2013).
It is informative to compare this bottom-up or geological approach to life's emergence—one that places steep geochemically produced proton gradients and redox disequilibria at the forefront—with Peter Mitchell's first and only foray into considerations of how life originated, and this from a top-down perspective. In 1957, during the time Mitchell was struggling toward formulating his chemiosmotic theory for adenosine triphosphate (ATP) synthesis, that is, how the energy currency of all cells is minted through the proton motive force rather than a high-energy organic intermediate in the glycolytic pathway (Mitchell, 1961; Prebble, 2002), he attended the First International Symposium on the Origin of Life on the Earth, and he presented a paper that illustrated his mind-set at the time. He began, I shall attempt to consider the mechanism whereby the contact between the organism and its environment is regulated, particularly in relation to the functions of the membranes that form the boundary between the organism and its environment. It will be appreciated that I cannot therefore consider the organism without its environment, and that from a formal point of view the two may be regarded as equivalent phases between which dynamic contact is maintained by the membranes that separate and link them. This circumstance serves at the outset to emphasize the fact that living organisms are distinguished, not by their momentary appearance, but by their behaviour and by their relationship to their environment. (Mitchell, 1959)
Referring to Mitchell's chemiosmotic theory—a theory judged by Orgel (1999) to compare with those of Heisenberg, Schrödinger, and Einstein (Lane, 2010)—Nick Lane reiterates the fact that life transcends mere “mass action” chemistry. Although facing the same thermodynamic landscape, life transcends simple chemistry (Lane, 2010) through the deployment of molecular, disequilibria- or free energy–converting nanoengines, many of them situated in membranes (Branscomb and Russell, 2013)—a point of view not accepted by Mitchell, though finally clinched by Boyer's demonstration of rotational catalysis in ATP synthase that undergoes conformational changes driven by the proton motive force (Boyer, 1975, 1997). All such engines work by mechanistically fusing an endergonic, “driven,” free energy–creating process to a (necessarily stronger) exergonic, “driving,” free energy–consuming one. For one example, Complex 1 exploits exergonic vectorial electron flow to drive protons from cytoplasm to periplasm endergonically in a system likened to the functioning of a steam engine (Efremov et al., 2010; Lane, 2010; Efremov and Sazanov, 2011a, 2011b; Baradaran et al., 2013)—electrons conduct to an acceptor, so driving protons to the periplasm. It might be speculated that ancestors of Complex 1 are traceable back to the [NiFe] site of hydrogenases (Darrouzet et al., 1998; Hedderich, 2004). The rotary ATP synthase, equated to the Wankel combustion engine, then uses the proton gradient resulting from the efforts of Complex 1 and the like as the exergonic driver to endergonically condense adenosine diphosphate (ADP) and inorganic orthophosphate (Pi) to ATP (Yoshida et al., 2001). ATP is the energy currency of the cell, but the process of making it forces it into a state in “astronomical disequilibrium” with respect to its hydrolysis products (Boyer, 1997; Branscomb and Russell, 2013). It is spent rapidly through the phosphorylation of organic molecules, thereby driving endergonic biosynthetic reactions through the elimination of the components of water.
The organic monomers that made up the relatively small pool acted on by ATP could not have been gleaned from the high entropy mix comprising the myriad of ill-assorted organic molecules comprising carbonaceous chondrites, many of them aromatic or highly branched, but were the product of C1 metabolism (Cody and Alexander, 2005; Fuchs, 2011). Yet we know that the abiotic reduction of carbon dioxide to formaldehyde or at least to a formyl group, for example, is highly endergonic, a reduction that challenges the theorist of autogenesis as it thwarts the experimentalist (Vladimirov et al., 2004). Of course, were there no steep thermodynamic barriers to be scaled in the fixation of carbon dioxide, there would hardly be a call for life to realize approaches toward redox equilibria on the outer surfaces of a wet rocky planet—catalyzed geochemical reactions would be adequate for the job. Nevertheless, there is an advantage to be gained from examining the transition from geochemistry to biochemistry from the bottom up, that is, to “look under the hood” at life's first free energy–converting nanoengines or “mechanocatalysts” (Russell, 2007; Nitschke and Russell, 2010; Branscomb and Russell, 2013; cf. He et al., 2012). Such an approach encourages us to see life as one of the last in a vast hierarchical cascade of emergent, disequilibria-converting entropy-generating engines in the Universe (Russell et al., 2013). In doing so, we keep our sights on the “astro” in astrobiology.
In systems driven far from equilibrium, self-organized dynamic structures, acting as engines (i.e., “free energy converters”), arise spontaneously (Cottrell, 1979). Their effect is invariably to accelerate the rate at which the driving disequilibrium generates entropy and is thereby dissipated (Prigogine, 1978). Tellingly, the Universe itself, at the moment of its Big Bang birth, was by a vast measure the most extreme example, known or conceivable, of a far-from-equilibrium system, born as it was in a condition of disequilibrium so great as to be virtually inestimable (Penrose, 2005). From this pinnacle of improbability, it could only, as the second law of thermodynamics demands, go endlessly “down” to ever increasing total entropy. Indeed, the history of the Universe has been “nothing but” the playing out of the dynamics of accelerated entropy production via emergent, self-organizing engines. All the dynamic structures and processes of the Universe, both great and small, from galactic superclusters to burning and dying stars, black holes, the writhing pirouettes of quasar jets, planetary systems, convective currents in myriad guises, to the poppies on the cool green hills of Earth—all are engines, all contributing members of this great self-organizing cascade of accelerated entropy production.
But in this medley of engines, black holes stand apart and have a special role to play. They are not only an end point in the production of entropy through gravitational collapse, at which the entropy per unit mass is a maximum; but almost all the Universe's entropy inventory is currently, and will increasingly be, in the form of supermassive black holes (Ruffini and Wheeler, 1971; Penrose, 2005; Scharf, 2012). Finally, they are themselves the most powerful and consequential engines in the Universe. When they accrete matter from other stars or the interstellar medium, they become extraordinarily powerful internal combustion engines, with fuel, carburetion, a combustion chamber, and multiple exhaust systems. They can be throttled from a quiet idling to an explosive roar of transgalactic impact. These engines are suspected of regulating the formation of stars and galaxies and driving their evolution, creating most of the magnetic flux in the universe, and ionizing the Universe itself shortly after the Big Bang (Meier, 2012).
We are thus justly, and most ironically, to be seen as the spawn of black holes, themselves the spawn of a Universe born in the greatest possible discomfort of disequilibrium. Without going into detail, we can follow the history of the Universe's engines from galactic and stellar processes through planetary accretion and geophysical convection to the emergence and evolution of life itself (Russell, 2007; Nitschke and Russell, 2010; Branscomb and Russell, 2013; Vattuone et al., 2013). And as the mist begins to lift from the divide that separates geochemistry from biochemistry, we can glimpse the biological bank that lies directly opposite. What is revealed is that, whereas life, like all other dynamic phenomena in the Universe, is forced to operate through the invention and deployment of engines to convert disequilibria—dissipating one to create another—the engines of living cells comprise a vast and uniquely complex heterarchical networked system of linked conversions. This great system, driven by externally supplied disequilibria, produces a myriad of internal “intermediate” and enabling disequilibria in the form of both structures and processes. These it uses, among much else, to drive the otherwise intractable reactions at the very beginnings of metabolic pathways up such steep thermodynamic gradients. The challenge before us, then, is to work out how it could all have gotten started.
In this contribution we consider the following:
(i) how the birth and earliest states of our own planet set the stage for life's onset,
(ii) the transition from geochemistry to biochemistry in a Hadean alkaline hydrothermal mound.
(iii) how well the top-down and bottom-up evidence converges on a model for life's origin,
(iv) exploration of those proto-biotic reactions that would have been at least mildly exergonic in the milieu offered by conditions within an alkaline submarine hydrothermal mound,
(v) identification of the endergonic challenge to the autotrophic pathways and the necessary requirement for carbon-fixation and proto-pyrophosphatase engines,
(vi) the nature of the guidance system that lifts life across the Darwinian threshold, and
(vii) briefly how other wet and rocky worlds elsewhere in the Solar System and the Universe might be evaluated for the presence of extant or former life.
2. The Birthing and Earliest Processes Operating on Our Own Planet
To set the scene for life's emergence, it is instructive to consider the birth of our solar system and the earliest few million years of our planet. Following the main stage of accretion from solar dust, chondrites, planetesimals, and Theia's impact, molten Earth cooled rapidly, possibly through the operations of whole mantle convection (Halliday, 2004; Zahnle et al., 2007; Gargaud et al., 2012; Sleep et al., 2014). Among the myriad of elements bequeathed to our planet by the hypernovae responsible for engendering this solar system were the radiogenic fuels 235U, 40K, and 26Al that would, in concert with gravitational energy, drive the thermal convective surfacing and resurfacing of Earth through mantle overturn, plumes, and/or impact melting (Turcotte and Schubert, 2002; Cockell, 2006; Harrison, 2009; Huss et al., 2009; Korenaga, 2013; Griffin et al., 2014; Harris and Bédard, 2014). In the absence of fully fledged plate tectonics in the Hadean (Johnson et al., 2013; Griffin et al., 2014), mantle plumes would have done the same job, though less efficiently (Fig. 1) (Bédard, 2006). The latter process would bring fresh mantle-derived material or mantle itself to the surface of this and other hot, wet, rocky worlds. Magmatic plumes are effectively engines with some positive (catalytic) feedback provided by the exothermic basalt-to-eclogite transition first recognized as an additional driving force by Holmes (1931) (Bédard, 2006; Sleep et al., 2014).
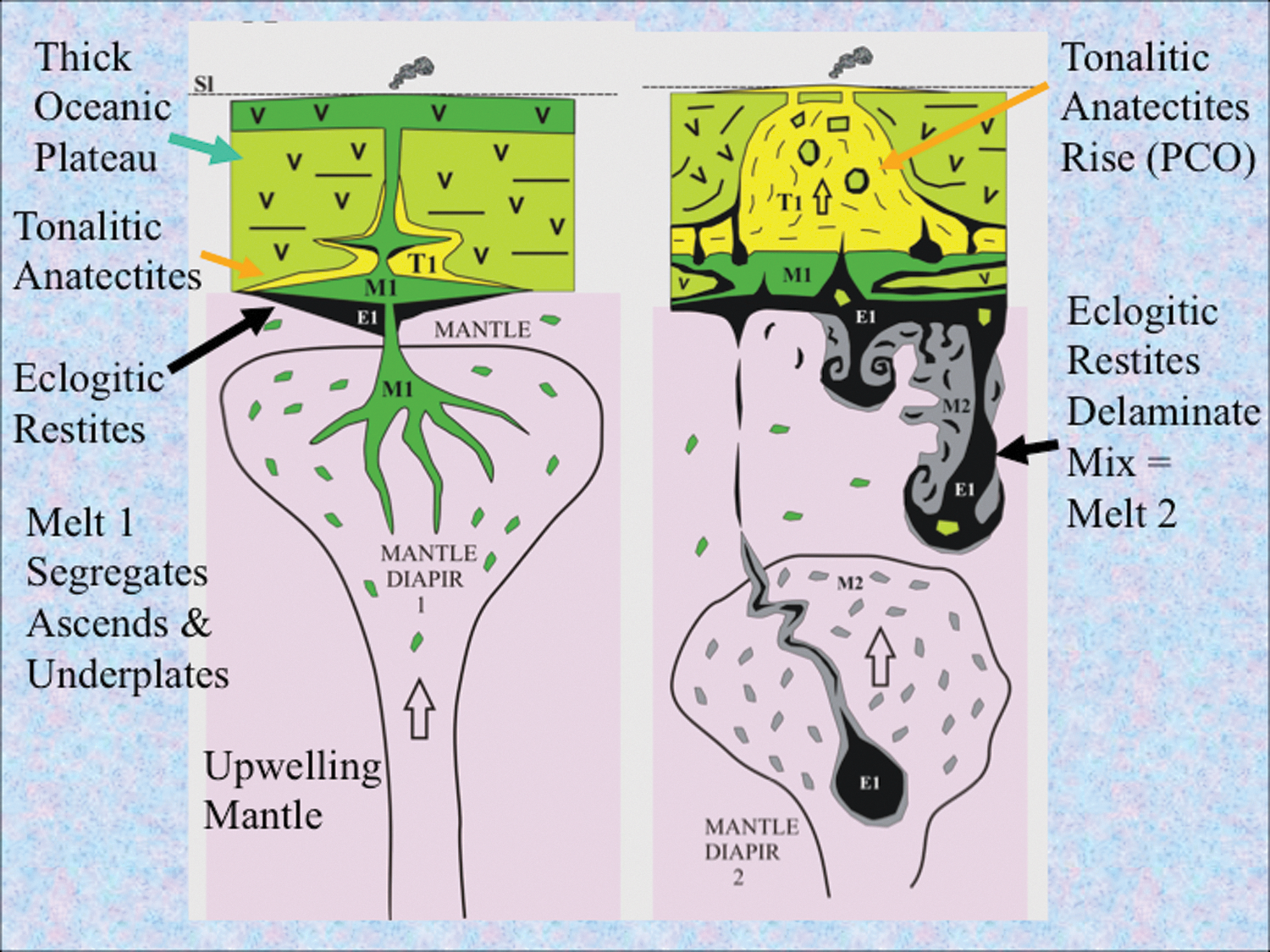
Diagrams showing the remelting of a thick volcanic plateau by a reinvigorated mantle plume, illustrating a delamination tectonomagmatic engine producing fresh ancient ocean crust on Earth continuously for 200–300 million years (Bédard, 2006). Left-hand cartoon shows molten magma (M1) generating a thick volcanic crust. This same batch of magma also ponds at the base of the crust, where it differentially melts the more siliceous and less dense component of the preceding volcanic host to produce quartz-rich tonalitic melt (T1). The more magnesium-rich residue partially metamorphoses to dense eclogite which begins to sink (E1). The right-hand cartoon shows how the buoyant siliceous melt invades the plateau to produce acidic (silica- and nickel-rich) volcanism and accompanying hot springs that have the potential to dispense carbon dioxide to the atmosphere and metal-rich solutions to the ocean, respectively (and see Sobolev et al., 2005). At the same time the dense eclogite (E1) delaminates and falls into the mantle. The larger bodies bore channels that may be exploited by further rising hot mantle diapirs. Smaller delaminated bodies mix into the shallow upper mantle and catalyze the formation of a second mantle melt (M2). Extraterrestrial wet and icy worlds with interiors hot enough to produce mantle plumes and thereby new ocean crust as well as oxidized volatiles could also set the stage for the onset of metabolism and life (and see Gaidos, 2000; Pasek and Greenberg, 2012). Reproduced from Bédard (2006) with permission.
While water, perhaps amounting to many present-day Earth oceans, was likely introduced from hydroxylated olivines in the solar disc (Alexander et al., 2012; Vattuone et al., 2013), much of the carbon was probably gained later as CO2 through delivery of icy comets from beyond the frost line, augmented by delivery of complex organic molecules in carbonaceous chondrites (Ehrenfreund and Charnley, 2000; Cody et al., 2001; Cody and Alexander, 2005; Yabuta et al., 2007; Orthous-Daunay et al., 2010; Kebukawa et al., 2011; Marty, 2012). Once absorbed into magma, any such organic molecules would pyrolyze to produce further carbon oxides and carbonate, only to be released at high temperatures in volcanic and hydrothermal exhalations, mainly as CO2 with smaller amounts of CO (Shock, 1992; Wood et al., 1990, 2006; Charlou et al., 2002; Trail et al., 2011, 2012). That portion of CO2 still retained in the crust would either disproportionate to graphite and methane during high-temperature (∼600°C to 500°C) serpentinization or be reduced to methane by inwardly diffusing hydrothermal hydrogen at, or above, 400°C (Welhan 1988; Kelley, 1996; Etiope and Sherwood Lollar, 2013; Etiope et al., 2013; Liu et al., 2013; McMahon et al., 2013; Ni and Keppler, 2013).
The first all-enveloping ocean rained out to a depth of perhaps 5 km or more from a proto-atmosphere that consisted of many bar of carbon dioxide and ∼2 bar of nitrogen and significant sulfur dioxide around 4.4 billion years ago (Bounama et al., 2001; Halevy et al., 2007; Martin et al., 2007; Zahnle et al., 2007; Elkins-Tanton, 2008; Goldblatt et al., 2009; Kotwicki, 2009; Trail et al., 2011; Maruyama et al., 2013). Any short-lived tidal pools on the margins of rare and rapidly weathered volcanic edifices puncturing the deep ocean surface were repeatedly swamped by high-amplitude tides generated by the more proximal Moon orbiting the more rapidly rotating Earth (Denis et al., 2011). Notwithstanding the suggestion that the early atmosphere was hydrogen-rich (Wordsworth and Pierrehumbert, 2013), H2 is likely to have gravitated rapidly to space, so beginning the oxidation of the planet's surface (Catling, 2006). Also, the solar wind blew up to 50% of the original inventory of atmospheric nitrogen to space (Ozima et al., 2005). The melting of bolides on impact may have produced occasional short-lived excursions to high CH4 partial pressures, though radiative reoxidation was likely rapid (Zahnle et al., 2010). And post the Moon-forming event, even large bolides could only have vaporized the near surface of the ocean at most (Abramov and Mojzsis, 2009). The Sun's radiation then only had around 70% of its present power, but the CO2, which predominated in the atmosphere, would have kept the ocean generally unfrozen except when a large portion of atmospheric CO2 was lost to hydrothermal convective interaction with the mafic crust and its sinking, or even subduction, to depth (Newman and Rood, 1977; Shibuya et al., 2010; Sleep et al., 2014). Even so, the CO2 would have been rapidly returned as the carbonated crust foundered and was subjected to thermal metamorphism (Dasgupta, 2013). Cosmic and solar clouds would also have masked the Sun on occasion and caused a drop in temperature. Since there was no solid core, a geomagnetic dynamo was not well developed, leaving the planet to the mercy of galactic and solar cosmic rays, which were especially strong during further occasional hypernova explosions in the then-crowded stellar environment (Baade and Zwicky, 1934; Montmerle et al., 2006; Labrosse et al., 2007; Melott and Thomas, 2011; Moeller and Hansen, 2013). Movement of the solar system in the Galaxy would have led to order-of-magnitude changes in cosmic ray intensity (Atri and Melott, 2012; Cohen et al., 2012). At the same time and for the same reason, protons and other charged particles from local gamma ray bursts both produced and entrained cascades of electrons, which thereby induced leader lightning strikes in thunderstorms (Gurevich et al., 1999; Gurevich and Karashtin, 2013). Cosmic and solar electron and proton rays are likely to have produced nitrogen oxides (NO x ) from CO2 and N2 on early Earth, the first directly and the second through the effects of lightning (Yung and McElroy, 1979; Reeves et al., 1992; Bieber et al., 1999; Martin et al., 2007; McLaughlin et al., 2008; Grenfell et al., 2012). Chemically similar atmospheres and oceans likely occurred on early Venus and Mars, which may have been conducive to the emergence of metabolism and life on those planets also (Russell and Hall, 1999; Mahaffy et al., 2013; Michalski et al., 2013; Vattuone et al., 2013; Webster et al., 2013; Wetzel et al., 2013). The icy moons of Jupiter and Saturn, as well as the myriad of extrasolar Earth-like planets of the kind revealed by Kepler, are likely to have broadly comparable chemical polarities (Schulze-Makuch and Irwin, 2002; Rugheimer et al., 2013).
Whatever the mechanisms that produced new ocean floor on Earth, the crust was probably thicker than it is today (Korenaga, 2008, 2013; Griffin et al., 2014; Sleep et al., 2014; and see Van Mierlo et al., 2013). In the case of plate tectonic mechanisms, slow spreading leads to the exhumation of mantle peridotites through gravitational slippage (e.g., the Indian ocean floor southwest of the ridge; Sauter et al., 2013). Otherwise, lavas involved in overplating were likely composed of high magnesium basalts and komatiites. Thus, the Hadean ocean floor would have been mafic to ultramafic with scatterings of trondhjemites, phosphate-rich volcanic glass, and xenoliths of schreibersite-bearing meteorites (Cody et al., 2001; Bédard, 2006; Pasek et al. 2013; Griffin et al., 2014).
The all-enveloping acidulous ocean [a feature that may have been common to oceans in icy worlds such as Europa (e.g., Carlson et al., 1999; Kargel et al., 2000; Schulze-Makuch and Irwin, 2002; Hand et al., 2007, 2010; Zolotov and Kargel, 2009; Pasek and Greenberg, 2012), as well as the original hydrospheres of Mars and Venus (e.g., Vattuone et al., 2013)] was the recipient of four main types of exhalation: (i) acidic volcanic gases (∼1500°C); (ii) hot (∼400°C), carbonated and highly alkaline submarine springs driven by mafic intrusives; (iii) hot (∼400°C) acidic tonalite–trondhjemite–granodiorite–associated submarine springs; and (iv) warm (∼100°C) alkaline springs and seepages resulting from serpentinization (Russell et al., 1989; Shock, 1990, 1992; Bounama et al., 2001; Douville et al., 2002; Botcharnikov et al., 2003; Hopkinson et al., 2004; Kelley et al., 2005; Martin et al., 2006; Proskurowski et al., 2006; Dauphas et al., 2007; Frost and Beard, 2007; Martin et al., 2007; Shibuya et al., 2007, 2010, 2012; Zahnle et al., 2007; Elkins-Tanton, 2008; Koschinsky et al., 2008; Kemp et al., 2010; Shanks and Thurston, 2010; Heck et al., 2011; Reeves et al., 2011; Arndt and Nisbet, 2012; Mloszewska et al., 2012; Travis et al., 2012; Klein et al., 2013; Schrenk et al., 2013). The combined effects of these exhalations were to create strong redox disequilibria between the reduced mantle and the relatively oxidized volatisphere (Fig. 2). The chemical gradients were thus set for the onset of life. Volcanoes supplied oxidized entities such as carbon dioxide, the main substrate for life, along with polyphosphates, sulfur dioxide, and nitric oxide (the latter supplemented by UV, cosmic rays, lightning, and meteorite impacts acting on CO2+N2). The hot (∼400°C) submarine acidic exhalations provided metals Fe>Mn≫ Zn>Co>Ni>W for catalysis and for the early enzymes. The hot (∼400°C) alkaline springs supplied calcium and silica, and the warm (∼100°C) alkaline springs and seepages produced the electron-rich fuels for metabolism, hydrogen, and methane along with materials such as ammonia and in some cases hydrogen sulfide (as HS−) as well as nanomolar thiomolybdate (Yung and McElroy, 1979; Kasting, 1990; Yamagata et al., 1991; Russell and Hall, 1997, 2006; Erickson and Helz, 2000; Douville et al., 2002; Krasnopolsky, 2006; Schumann and Huntrieser, 2007; Martin et al., 2008; Smirnov et al., 2008; Nitschke and Russell, 2009; Erlykin and Wolfendale, 2010; Grenfell et al., 2012; Schoepp-Cothenet et al., 2012; Gordon et al., 2013; Helz et al., 2013).
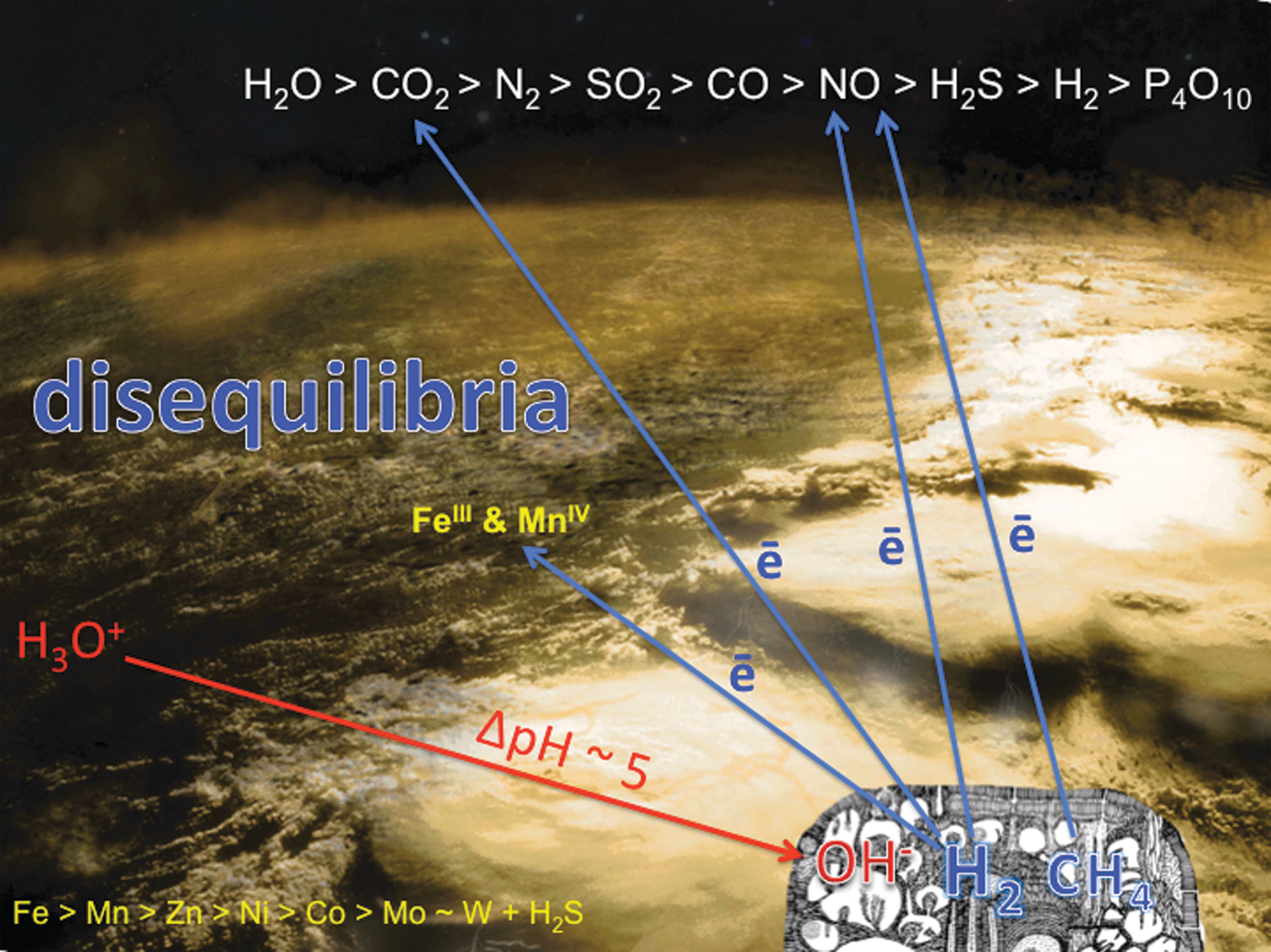
The candidate pH and redox disequilibria driving the onset of biochemistry on wet rocky worlds. Oxidized and acidic volatiles are supplied to the atmosphere and hydrosphere by volcanoes (top); acidic ∼400°C springs supply the metals and H2S to the ocean (lower left); iron and manganese from these springs are photooxidized (center); reduced entities exploited by emergent metabolism are mainly supplied in alkaline hydrothermal solution to a submarine precipitate mound (lower right) (Nealson and Saffarini, 1994; Douville et al., 2002; Proskurowski et al., 2006; Martin et al., 2007; Zahnle et al., 2007). These are the initial far-from-equilibrium conditions considered to have driven the onset of life.
3. The Hadean Hydrothermal Hatchery of Life—The First Transition
The nature of the porous mounds produced at the warm alkaline submarine spring that we consider to have been the hatchery of life may be surmised from extant springs today, as well as from laboratory simulation experiments. At Lost City, chimney structures comprise brucite (Mg(OH)2) enclosing friable filaments of aragonite (Kelley et al., 2005; Ludwig et al., 2006). The brucite appears to have precipitated around the aragonite (CaCO3) as a result of the hydrothermal solutions mixing with ocean water (Kelley et al., 2005). Very high Fe-brucite ([Fe,Mg][OH]2) and ferrous/ferric oxyhydroxides indicative of the mixing of similar hydrothermal solutions with ocean water have been intersected in peridotitic ocean floor on the Iberia Abyssal Plain (Hopkinson et al., 2004). A vestige of what may have been an extinct mound has also been recorded on the ultramafic floor of the western Indian Ocean (Bonatti et al., 1983). Those remains consist of the expanding clay, sepiolite (Mg4Si6O15(OH)2·6H2O), and poorly crystallized magnesium and iron (layered double?) hydroxides. Two analyses of the sepiolite revealed concentrations of Ni at 150 and 365 ppm, Co at 15 and 67 ppm, V at 23 and 78 ppm, and Zn at 120 and 225 ppm. Layered double hydroxides such as green rust (now termed fougèrite) comprise cationic brucite-like layers alternating with hydroxyls, water molecules, and/or a variety of anions (Trolard et al., 1997; Braterman et al., 2004; Schoonen et al., 2004; Génin et al., 2005, 2006; Ruby et al., 2010).
How would such mounds compare to those generated on early Earth? While the alkaline solutions derived from serpentinization would have been much the same, except for the occasional bisulfide contribution, the anoxic acidulous Hadean Ocean (pH ∼5.5), with its load of iron and other transition metals, silica, phosphate and polyphosphate, phosphite, sulfide (as H2S), as well as nitrate and nitrite, was very different (Macleod et al., 1994; Pasek et al., 2013). Without catalytic surfaces or biota to promote reactions, these entities would have remained supersaturated, only precipitating on meeting the hydrothermal alkaline effluents. Depending on depth, a portion of the iron may have precipitated as siderite (Macleod et al., 1994). If so, this iron carbonate could have been replaced by greigite (Fe3S4) as any bisulfide-bearing hydrothermal solutions percolated through the mound (Sagnotti et al., 2005). Alternatively, the iron could have been precipitated directly as mackinawite (FeS) (Mielke et al., 2011). And in place of brucite and iron-rich brucite, the hydroxides would have been much more iron-rich. Indeed, because of oxidants in the Hadean Ocean, we imagine a major constituent of the first precipitate membranes would have been the mixed-valence brucite-like ferrous/ferric oxyhydroxide—fougèrite (green rust) (Trolard and Bourrié, 2012). In our attempts to simulate such springs in Hadean conditions, we have generated chimneys and micro- and nanocompartments comprising mackinawite (FeS), greigite (Fe3S4), silica, as well as the ferrous-ferric oxyhydroxides (Russell et al., 1989, 1994; Mielke et al., 2010, 2011). Mielke and coworkers (2011) argued that these micro- and nanocompartments would restrict diffusion of hydrothermal fluids and thereby focus reactions with the invasive carbonic ocean at, or within, their boundaries “and act as ‘low-entropy’ traps whereby compatible products would be forced to interact through their proximity” (cf. Westheimer, 1962; Page and Jencks, 1971).
We argue here that metabolism was forced to emerge in such fine compartments to resolve the disequilibrium between the hydrogen and methane delivered in an off-ridge alkaline hydrothermal convection cell, with volcanic carbon dioxide and other oxidized entities such as nitric oxide dissolved in the acidulous Hadean Ocean (Nitschke and Russell, 2013) (Fig. 2). When particular critical disequilibria are reached on wet rocky worlds, convection and metabolism are two autocatalytic symmetry-breaking robust processes that take over from mere conduction and aqueous geochemistry, respectively (Russell, 2007). So although, in our opinion, early biochemistry emerged from, and was coupled to, the off-ridge or off-mantle plume convection cell, it does not follow that biochemistry is merely better geochemistry (Martin and Russell, 2007). However, knowing the environment from which life sprang and the commonalities between the geochemistry and metabolism can surely help us understand life's first steep ascents and faltering steps, as we explore below.
4. Top-Down and Bottom-Up Evidence Converges on an Autogenic Model for Life's Origin
At life's emergence, our planet was basic and relatively reduced on the inside yet somewhat acidic and oxidized on the outside (Russell and Hall, 1997). As such it compared to the polarity of a fuel cell or a simple-celled prokaryote (Russell et al., 2003; Nitschke and Russell, 2009; Lane et al., 2010; Barge et al., 2014). As we have seen, it exhales oxidized volatiles such as H2O, CO2, SO2, and NO at high temperature—molecules that are reduced geochemically to H2, CH4, H2S, and NH3 as they are entrained in hydrothermal convection cells traversing the oceanic crust (Fig. 2). Although tempting, we must (have learned to!) resist the urge to assume that prokaryotes first emerged to “quicken” these geochemical processes (Nitschke and Russell, 2013; cf. Russell et al., 1998). It makes more sense to see the earliest cells as exploiting the fuels offered by this gigantic inanimate body, that is, as being dependent on the environment of early Earth (Nitschke and Russell, 2013; cf. Mitchell, 1959). This view refocuses our interest on a site where the fuels were presented in the most acceptable forms—an alkaline submarine spring where the same general polarity holds while the H2, CH4, HS−, and NH3 are concentrated, palatable, and steadily and continuously supplied, and there is all-important opportunity for the disposal of waste, that is, the release of entropy (Fig. 3) (Russell et al., 1989, 2010; Schoonen and Xu, 2001; Smirnov et al., 2008; Mielke et al., 2010; Gordon et al., 2013).
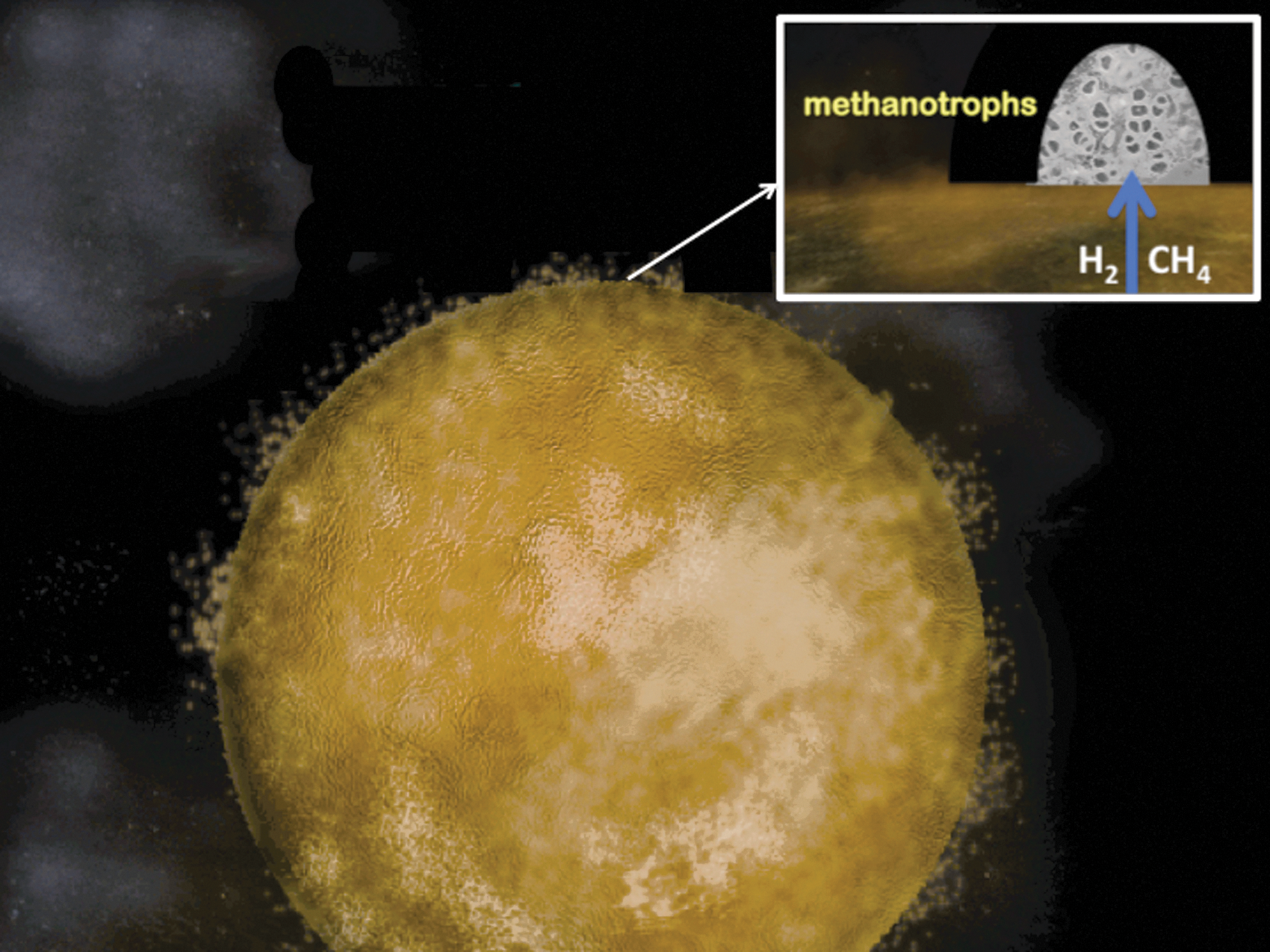
Picture of the Hadean water world. The first ocean was ≥5 km deep, entirely submerging any proto-continents (Bounama et al., 2001; Elkins-Tanton, 2008). Inset sketch shows how a proto-biotic methanotrophic community could have been nurtured by the methane and hydrogen emanating at a submarine alkaline vent on the Hadean Earth's ocean floor. The polarity of the water world is similar to that of a prokaryote, generally reduced on the inside and oxidized on the outside so that electrons are transported by various mechanisms toward the exteriors of both (Russell and Hall, 1997). Protons (or acidic volatiles) are also continuously pumped to the exterior of both, the one through volcanoes and certain very hot springs, the other by various enzymes such as Complexes I, III, and IV. (Photographs courtesy of Minik Rosing and Billy Brazelton.)
From this point of view, it can be argued that the electro-geochemical state of the abiotic planet drove the emergence of an autotrophic metabolism, a metabolism given to the reduction of carbon dioxide and the concomitant oxidation of methane and hydrogen. In Table 1, we itemize features and requirements of life and compare these with what is on offer from the abiotic world, both with respect to various disequilibria and to the materials that could be put to use by emergent metabolism. The geological and geochemical studies of what is known of early Earth as they pertain to such an emergence of life appear to be converging with enzymological and phylogenetic retrodictions from what is known of the ancient metabolic pathways employed by organisms that occupy the lowest branches of the evolutionary tree, at least as they appear to us today (Table 1) (Say and Fuchs, 2010; Ducluzeau et al., 2014; Nitschke et al., 2013; Schoepp-Cothenet et al., 2013).
Life emerging from this world faced a two-front “endergonic” battle to attain the fixing of carbon and the generation of an extreme pyrophosphate/orthophosphate disequilibrium. We consider carbon fixation first and argue that hydrogen—split into two electrons and two protons at Fe-Ni sulfide clusters comprising the membrane—reduced the carbon dioxide that permeated the inorganic walls of the compartments where water activity was low to carbon monoxide, a mildly anti-entropic reaction involving redox bifurcation mediated probably by molybdenum or tungsten (McGlynn et al., 2009; Nitschke and Russell, 2013). At the same time, we surmise that the hydrothermal methane was oxidized endergonically with nitrate or nitrite to methanol at FeIII sites in fougèrite (green rust) in a reaction also involving molybdenum or tungsten sites with electrons released from hydrogen at another (higher potential) Ni-Fe sulfide site (Antony et al., 2008; Nitschke and Russell, 2013) (Fig. 4). The methanol was further oxidized with nitrite or the ephemeral NO to formaldehyde at a Mo/W site before being re-reduced and thiolated to a methyl group on a Ni-Fe sulfide cluster. Here, it condensed with the CO before being released on reaction with a second methane thiol as methyl thioacetate, a likely entry point into biosynthesis through further hydrogenations, condensations, phosphorylations, carboxylations, and aminations (Huber and Wächtershäuser, 1997, 2003; Russell and Martin, 2004; Nitschke and Russell, 2013). The polar arrangement, acting at the hydrothermal mound, has held steady over the 4 billion years, even to our own physiology, ideally poised around pH 7.4, with hydrogen for biosynthesis contributed now by NAD(P)H (Booth, 1985; Kellum, 2005; Russell, 2007; Williams and Ramsden, 2007; Spitzer and Poole, 2009) and the electron acceptor oxygen—in place of CO2, oxidized nitrogen entities, and FeIII—produced through oxygenic photosynthesis. Then as now, trace elements, particularly some d-block metals, have to be delivered in optimal concentrations to play their part in hydrogenation, dehydrogenation, carboxylation, and electron transfer (Nitschke et al., 2013). It is likely that, in the absence of genetic guidance, some of the nitrate and nitrite may have been reduced all the way to ammonia on oxidation of a portion of the fougèrite to magnetite and of mackinawite to iron oxides and oxyhydroxides, so adding a peripheral source for assimilation of this key molecule (Table 2) (Hansen et al., 1996; Hansen and Koch, 1998; Gordon et al., 2013).
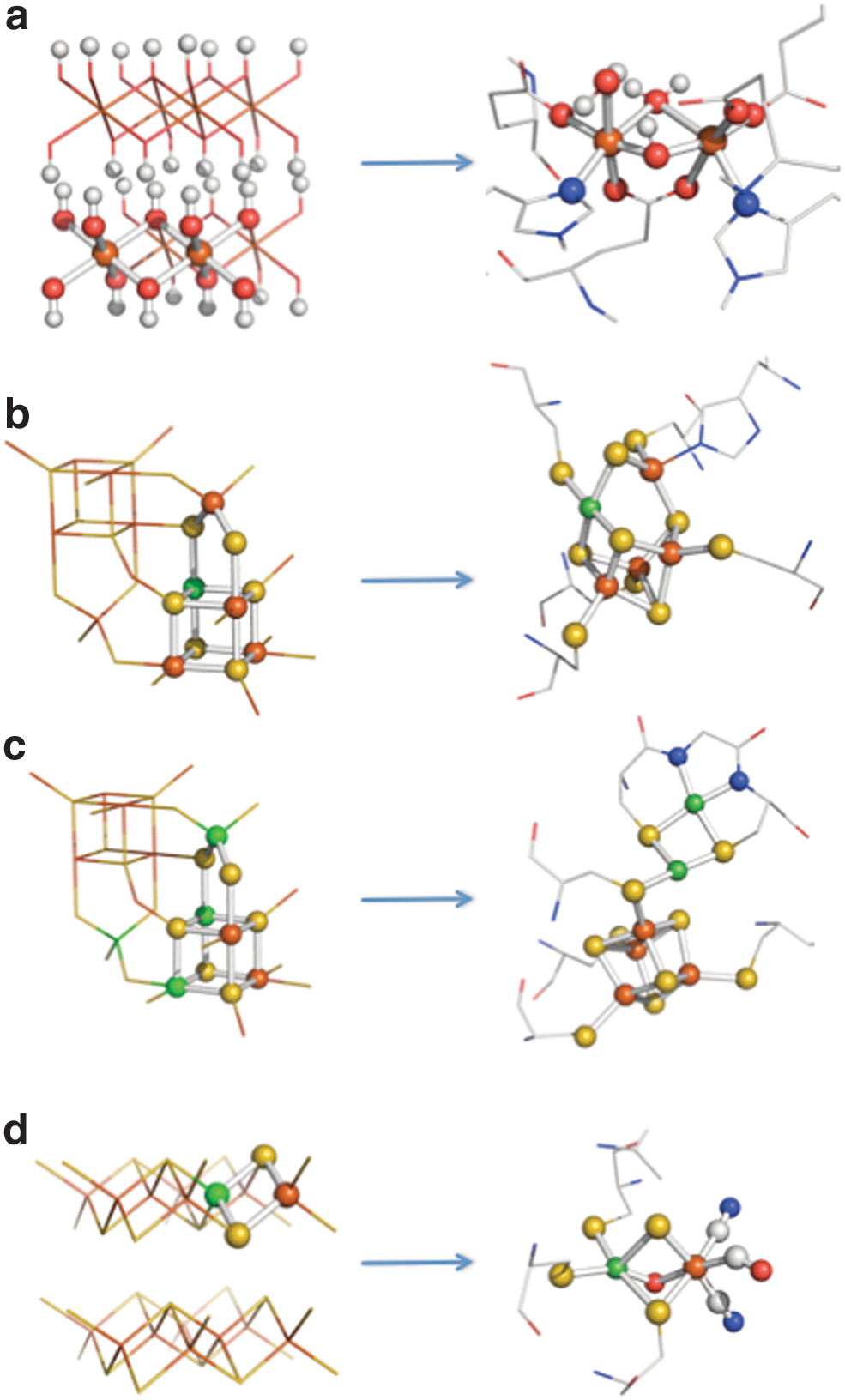
Structural comparisons between transition element sulfides and oxides of the kind found in the precipitate membranes with the active sites of metalloenzymes present in the LUCA: (
Figures 1 –3 sketch the overall geological and geochemical context of the onset of this putative denitrifying methanotrophic acetogenic metabolism from the bottom up, while Fig. 5 demonstrates what we consider to be the two major dynamic phase transitions, that is, (i) from geochemistry to a metabolism whereby information is directly propagated through mineral and peptide recognitions, growth, and other thermodynamic physicochemical imperatives (Cairns-Smith, 2008; Milner-White and Russell, 2011; Greenwald and Riek, 2012; cf. Hoffmann, 2012; Kang et al., 2013) and (ii) from metabolism to a fully functioning recognizable biochemistry through coded guidance toward attractor states (Corliss, 1986; Koonin and Martin, 2005; Yarus, 2011; Goodwin et al., 2012). A particular major commonality between modern cells and the hydrothermal mound lies in the structures of the metal sulfide and oxide enzymes and their similarity with mineral sulfides and oxides likely to be precipitated in the hydrothermal mound—themselves subject to proton, redox, and temperature gradients (Russell and Hall, 1997, 2006; Baymann et al., 2003; Volbeda and Fontecilla-Camps, 2006; Rickard and Luther, 2007; McGlynn et al., 2009; Rivas et al., 2009; Helz et al., 2013; Nitschke et al., 2013) (Fig. 4).
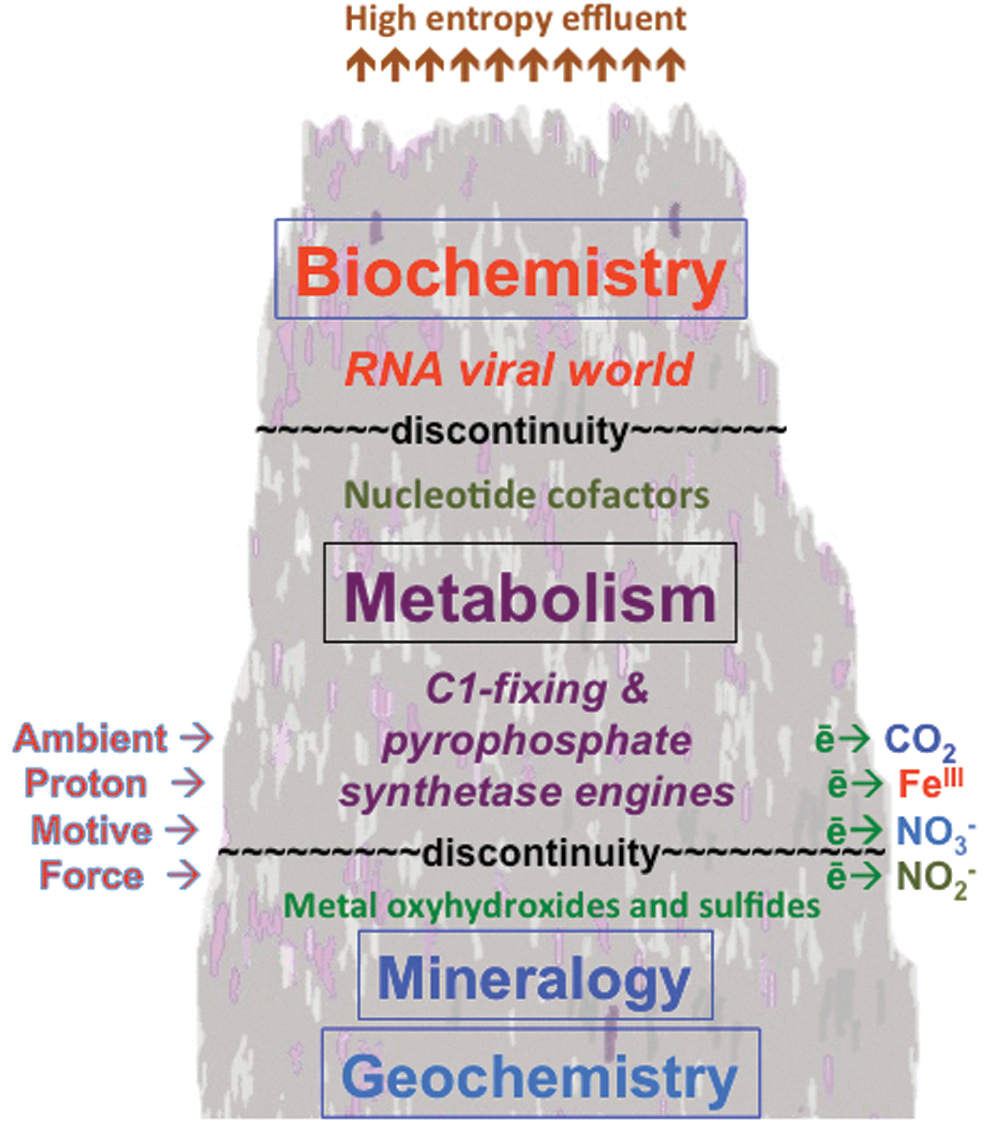
This electro-geochemical hydrothermal system occupying a growing compartmentalized mound evolves through mineral recognition and propagation processes leading, through an amyloidal takeover, to a cofactor world which transforms through the stringing of bases along a peptide-like backbone into the RNA world capable of Darwinian evolution and the emergence of fully fledged autotrophic life (Russell et al., 1994; Milner-White and Russell, 2008; Yarus, 2011; Goodwin et al., 2012).
Our current hypothesis differs from our earlier formulations (Russell and Martin, 2004) in that we now invoke the use of methane as well as hydrogen as both fuel and substrate, and call on nitrate and nitrite to oxidize the CH4, while hydrogen, assisted by the ambient steep proton gradient, reduces CO2 to CO (Nitschke and Russell, 2013). We now consider that CH4 and CO2 are equal sources of carbon for proto-biosynthesis (Fig. 6a). This contrasts with the Huber and Wächtershäuser (1997) view that the primordial initiation reaction for a chemoautotrophic origin of life was the acetyl coenzyme A pathway pulled solely in the reductive direction from CO2 by pyrite formation.
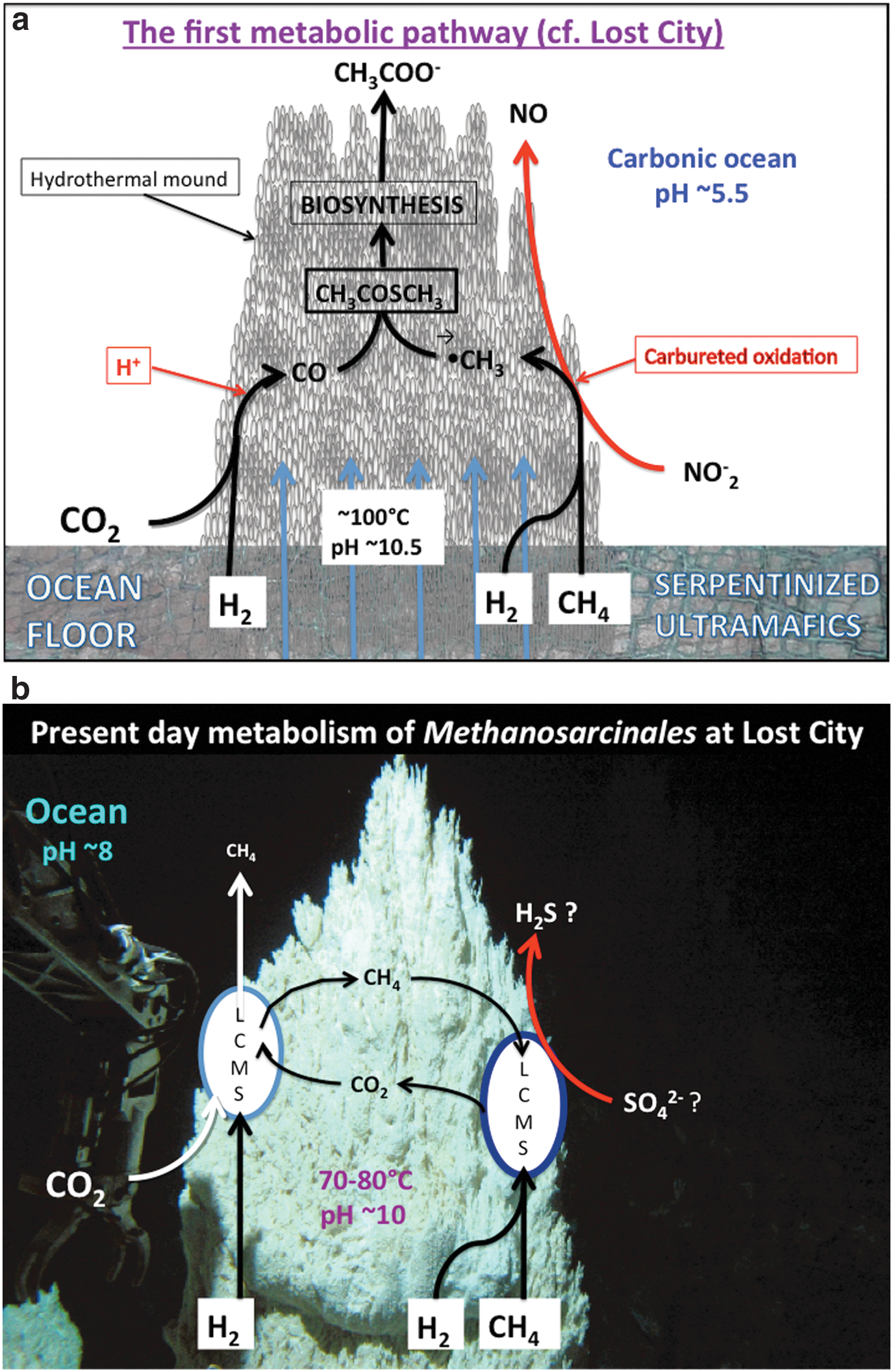
(
The discovery of the Lost City vent field in 2000 inspired new interest in the alkaline hydrothermal hypothesis (Kelley et al., 2001; Martin et al., 2008). And the ongoing operations of the particular biota elucidated there are equally encouraging, since one of the dominant microbiological processes as exploited by one type of Methanosarcinales reprises what we take to be the very first biochemical process—one that exploits both hydrogen and methane as fuel and sulfate as an electron acceptor (Fig. 6b) (Brazelton et al., 2011). (In our view the chief electron acceptor and provider of the oxygen to methanol in the first metabolist would have been nitrite rather than sulfate.) The aspect of the theory that calls upon methane as one carbon source is given further credence in principle through the identification of an archaeon, “Candidatus Methanoperedens nitroreducens,” that oxidizes methane using nitrate as an electron acceptor through a complete reversal of the methanogenesis pathway (Haroon et al., 2013). Of course, we are not suggesting that these methanotrophs are in any way the direct descendants of those microbes that first emerged at the alkaline submarine mound, but merely that they facilitate a similar metabolism. Once these first, and steepest, of steps along pathways that lead to activated acetate were scaled, that is, CO2 reduction and CH4 oxidation, a gently undulating thermodynamic plateau was reached where subsequent reactions proved to be either exergonic in mildly alkaline solution or required a limited infusion of free energy (Table 2). Such reactions are considered next.
5. Exergonic Reactions Favored by the Alkaline to Acid Hydrothermal Environment
Hot springs in mafic and ultramafic environments all produce reduced molecules, and those alkaline springs exhaling at Lost City are no exception (Kelley et al., 2001; Douville et al., 2002). The Lost City alkaline solutions carry millimolar concentrations of hydrogen and methane, and micromolar amounts of formate and nanomolar concentrations of C2–C4 hydrocarbons (Proskurowski et al., 2006, 2008; Konn et al., 2009; Lang et al., 2010). Laboratory studies on organic molecules dissolved in alkaline solution were first pursued by Butlerov (1861). In experiments with some affinity to biochemical cycles, he demonstrated the synthesis of a variety of sugars from formaldehyde in solutions dosed with calcium hydroxide. Once trioses are formed in such experiments, more specific oligomerizations to tetrose and fructose ensue (Table 2). Indeed, Mellersh and Smith (2010) pointed out that “life uses a reaction very similar to that of Butlerov but controls it using phosphorylated substrates.” In this regard, it is notable that glyceraldehyde, an early product, is easily phosphorylated to glyceraldehyde-3-phosphate (Kolb and Orgel, 1996). Huber and Wächtershäuser (1997) demonstrated the benefit of alkaline solutions in their production of activated acetate—the substrate for biosynthesis in the acetyl coenzyme A pathway—from methane thiol and CO with nickel as catalyst (and see Fig. 6a). A key reaction in the pathway is the non-enzymatic condensation of formaldehyde to methylene tetrahydrofolate (Friedkin, 1963), a vital step in the oxidative tributary to the proto-acetyl-CoA pathway as newly formulated by Nitschke and Russell (2013). Coenzyme A itself is a thiol, and life has taken full advantage of the fact that sulfur is equally at home ligated to the transition metals as it is bound to carbon and with itself (Beinert, 2000a, 2000b). Coenzyme B and coenzyme M, which are involved in methanotrophy and methanogenesis, are also thiols. The ability of sulfur to bind with itself is significant not only to catalysis and energetics but also to the early development of homeostasis; organic sulfides could have been directly converted to insoluble dithiols through the action of protons derived from the acidulous ocean through the membrane (Russell et al., 1994). Bracher et al. (2011) showed how thiol-thioester exchange of organic moieties is more rapid than hydrolysis at relatively high pH (and see Schouten et al., 1993; Schneckenburger et al., 1998), which suggests that sulfur-mediated exchange processes are more important than oxygen-mediated processes.
There are many other experimental examples of the synthesis of organic entities from the simplest of molecules in alkaline hydrothermal solutions of moderate temperature as would be expected from the alkaline hydrothermal theory. This is because alkaline conditions produce unprotonated organic anions, thiols, and phosphates that engender cascades of interactions and condensations. Table 2 lists a variety of reactions central to biosynthesis and proto-enzyme formation. Further examples are the synthesis of ammonia, and thence amino acids and peptides (Hansen et al., 1996; Huber and Wächtershäuser, 2003; Huber et al., 2003), the latter possibly generated on sphalerite or cleiophane (pristine ZnS) (Ohara and Cody, 2010; cf. Boyce et al., 1983). At comparable pH values, two iron sulfides, mackinawite and its partial oxidation (or sulfidation) product greigite, are formed with structures similar to those in iron-sulfur proteins (Table 3, Fig. 4) (Russell and Hall, 1997, 2009; Rickard and Luther, 2007; Lin et al., 2009; Csákberényi-Malasics et al., 2012; Nitschke et al., 2013). Indeed, the iron-sulfur cubane component of greigite can be thiolated to give a structure comparable to the cysteine-ligated ferredoxins (Bonomi et al., 1985). Milner-White and Russell (2005, 2011) suggested that the backbones to short (uncoded) peptides could act as the “nests” for both the HPO4 2- and the [Fe4S4(SC6H5)4]2- “eggs.” The theoretical calculations of Hong Enriquez and Do (2012) indicate that peptides 8–10 residues long would even be capable of sequestering bare Fe4S4 clusters, especially in the cooler reaches of the hydrothermal mound. Moreover, a soluble peptide involving a lysine, with a basic terminal amine residue, has been shown to sequester phosphate in a manner highly reminiscent of the P-loop motif found in all prokaryotes (Bianchi et al., 2012). Short peptides can also sequester the catalytic metal atoms nickel and cobalt (Martin et al., 1960; Ma et al., 1967). We can imagine a selection pressure that favors the sequestering of the smaller anions or cations by the longer peptides and thereby a positive feedback that improved rates of catalysis and proto-metabolism.
We note here that there are exceptions to the alkaline tendency to promote reactions. For example, glycylglycine (generated at high pH) can be condensed through the activation of the carboxyl groups to tetraglycine with trimetaphosphate at pH 6.5 (Yamanaka et al., 1988); and oligoglycines (gly4≫ gly6) are generated, also with trimetaphosphate, optimally at pH 5 and 38°C with 0.5 M MgCl2 as catalyst (Yamagata and Inomata, 1997) (Table 4). Pyrophosphate itself has also been produced from acetyl phosphate and orthophosphate at pH 6.5 (de Zwart et al., 2004; Barge et al., 2014). These findings could be interpreted to favor the view that dipeptides were polymerized to tetrapeptides and beyond on the outer margins of the inorganic membrane. If so, one could imagine a peptide takeover of the inorganic membrane/wall in such a way that would not only improve its outer structure but also retain the inorganic centers vital to catalysis and electron transfer, thus improving their reactivities (Violante et al., 1995; Mielke et al. 2011; Milner-White and Russell, 2011; McGlynn et al. 2012). These conditions also favor reactions that play into stages or offshoots of the reverse (incomplete) tricarboxylic acid cycle (Fuchs and Stupperich, 1978; Wang et al., 2011). The iron sulfide catalysts of the type used by Wang and his coworkers should also be precipitated in a slightly more bulky form in the more acidic conditions that obtain at the outer periphery of the inorganic membranes (Rickard et al., 2001; Wolthers et al., 2003) (Table 3).
If these reactions were all there were to early life, we might ask what the problem is. It is those very first steps of carbon fixation and the ongoing requirement for condensations driven against water activity that are the challenge. We turn to these “uphill” entropy-reducing reactions next, bearing in mind that some of these could be coupled mechanistically to the exergonic reactions described above.
6. Endergonic Reactions and the Need for Engines
Once a metabolic system is established, many of the myriad of chemical reactions that take part in proto-biochemical metabolic pathways, cycles, and networks are mildly exergonic, even in the absence of oxygen, as we have just detailed, and feed back positively to the organization (Amend and McCollom, 2009, Tables I, IV, and V; Amend et al., 2013). It is the entry points to the autotrophic pathways exacting a high free energy price that must first be assailed (Maden, 2000).
So what were the thermodynamic barriers that had to be climbed before life could get going from scratch? The first and most obvious is the fixation of carbon. It has been argued that, as the acetyl coenzyme A pathway is the simplest and probably the most ancient and cheapest energetically, and is used by certain autotrophs, all that has to be done is to find a way of copying it in an abiotic world in the lab (Wächtershäuser, 1990; Russell and Martin, 2004). It does look straightforward, one CO2 molecule is reduced to CO while another is reduced to a methyl group, and the two are then condensed to an activated acetate. Yet only the last acetate-forming step has been convincingly demonstrated (Huber and Wächtershäuser, 1997); the reduction of carbon dioxide at moderate temperatures beyond formate at significant yield has so far not been achieved in a laboratory setting. The abiotic reduction of CO2 to formaldehyde in aqueous solution is stymied by thermodynamic barriers so steep that even electrochemical efforts have failed the ascent (Maden, 2000; Vladimirov et al., 2004; Narayanan et al., 2011; cf. Schoonen et al., 1999). The second barrier is to condensation reactions in an aqueous environment, particularly the condensation of monophosphates and amino acids. Thus, because abiotic processes are not energetically conducive to carbon fixation and polymerizations, approaches that couple exergonic processes and steep gradients to these endergonic reactions by using a nanoengine are now considered.
7. Searching for a Carbon-Fixation Nanoengine
The oxidation of methane may offer a back door to the synthesis of activated acetate. Starokon and coworkers (2011) provided an indication of how this may have been facilitated. These authors oxidized methane to methanol even at room temperature through the α-oxygen bonded to a cationic Fe2+ site standing proud within the cage of a manufactured synthetic iron-bearing pentasil zeolite, which acts too as an entropy trap (and see Panov et al., 2002; Kachurovskaya et al., 2004). However, it must be emphasized that the α-oxygen-rich zeolite driving the reaction is prepared through endothermic oxidation of the synthetic zeolite by nitrous oxide at 200–250°C (Starokon et al., 2011, 2013; and see Schwarz, 2011; Sazama et al., 2013). Nevertheless, a further indication is afforded by known redox changes in ferrous hydroxides and fougèrite—clusters similar in structure to methane monooxygenase—that also facilitate aminations of carboxylic acids (Huber and Wächtershäuser, 2003; Nitschke et al., 2013) (Fig. 4). Microbiological support for a methanotrophic tributary to the acetyl-CoA pathway may be gleaned from the work of Ettwig and coworkers (2010). These authors showed that the oxidation of methane is achieved anaerobically by Methylomirabolis oxyfera through the concomitant reduction of nitrite. And Haroon and coworkers (2013) also demonstrated the biotic oxidation of methane with nitrate by the archaeon Methanoperedens nitroreducens. However, the process appears to be merely the reversal of methanogenesis, so it can hardly be considered primordial. We surmise that nitrite might drive a similar, though less efficient, partial oxidation of fougèrite, and that, given its structural similarity to the methane monooxygenase enzyme used by Methylomirabolis oxyfera, might have enabled the same reaction—an idea open to experimentation (Yoshizawa et al., 2000; Ettwig et al., 2010; Nitschke and Russell, 2013; Nitschke et al., 2013).
A pterin-molybdenum cofactor is required as part of the enzyme complex that facilitates oxidation of methane to a methyl group, a process which involves redox bifurcation with oxygen derived from NO (Ettwig et al., 2010). Martin and Russell (2007) pointed out that pterins and the like “might be more critical in getting an ancient but very complex biochemistry going than has traditionally been presumed,” and this requirement appears ever more acute and remains unresolved (cf. Rebelo et al., 2003). Because of the easy passage between the oxidation states of molybdenum, the metal's involvement in redox bifurcation, its facility to bind sulfide (especially as MoIV) and oxygen (as MoVI), its proven wide use as an industrial catalyst, and its ubiquity throughout all life's domains, we assume that it acted in some form as a precursor to the pterins. Hence, our interest in the search for an abiotic locale for an active molybdenum atom prior to pterins becoming available, which would act as the redox bifurcation nanoengine (Branscomb and Russell, 2013). Inorganic cluster possibilities to be investigated in this regard include Mo atoms contained in sites in a mackinawite (FeS) cluster, linear tetranuclear clusters [(MoO2S2)Fe2S2(MoO2S2)]4-, [(MoS4)Fe2S2(MoS4)]4-, and [(MoS4)FeO(OH)(MoS4)]3-, or as Mo(CN)8 4-/3- in hydrotalcite, a layered double hydroxide comparable to fougèrite (Anglin et al., 1987; Itaya et al., 1987; Nitschke and Russell, 2011; Geim and Grigorieva, 2013; Helz et al., 2013; Nicolosi et al., 2013) (Table 3). The linear sulfide clusters in particular may have been ligated non-covalently to a peptide (Burgmayer and Stiefel, 1985; Rivas et al., 2009; Nitschke and Russell, 2011; Schoepp-Cothenet et al., 2012). To be borne in mind when considering a possible precursor is the need for exposed sites for the molybdenum atom(s) and for protonation sites, perhaps afforded by the distal sulfurs (Nitschke and Russell, 2009; Jacques et al., 2014). Of course, once short peptides involving serine and cysteine became available, a scaffold for a peptide-coordinated molybdenum center might have emerged to act as an interim redox-bifurcating catalyst (Dutta et al., 2012; Jacques et al., 2014).
The other intermediate in the denitrifying methanotrophic acetogenic pathway is carbon monoxide, derived by the reduction of carbon dioxide in a reaction that is also thermodynamically uphill and therefore also requires a proton gradient and redox bifurcation (Maden, 2000; Nitschke and Russell 2009, 2013). In our hypothesis, the catalyst for this reaction is a nickel-bearing greigite cluster (SNiS[Fe4S4]SFeS), the assumed precursor of carbon monoxide dehydrogenase (cf. Vaughan and Craig, 1978; Dobbek et al., 2001) (Fig. 4). A single electron from a contiguous molybdenum site is presumed to reduce the greigite as the paired electron reduces fougèrite, which itself was previously oxidized by nitrite or nitrate (cf. Ragsdale, 1997; Hansen and Koch, 1998). (In our view, both electrons are derived through the oxidation of hydrothermal hydrogen to protons on a nickel-bearing mackinawite and are, we imagine, attracted separately to the respective FeIII sites—one “uphill” to the greigite and one “downhill” to the fougèrite.) Once the CO is sited as a nickel-carbonyl entity on the greigite, ligand-accelerated autocatalysis promotes reaction with methane thiol to produce the activated acetate (Huber and Wächtershäuser, 1997; Wächtershäuser, 2007; Haydon et al., 2011).
In this formulation of the acetyl coenzyme A pathway, the two proto-tributaries—one reducing CO2 to CO, the other oxidizing and sulfidizing CH4 to CH3SH—are driven from either end of redox space to converge as the methyl thiol reacts with a CO group coordinated to a nickel atom, itself a constituent of a greigite cluster that was one occupant of the precipitate membrane and acted as a precursor to acetyl coenzyme A synthase (Fig. 4) (cf. Huber and Wächtershäuser, 1997; Crabtree, 1997; Russell and Martin, 2004). The resulting acetyl methyl sulfide (CH3COSCH3)—the supposed precursor of acetyl-CoA—is the key intermediate to further hydrogenations, carboxylations, phosphorylations, and aminations deriving from stages in the incomplete reverse tricarboxylic acid cycle terminating at succinate and glyoxylate (de Duve, 1991; Morowitz et al., 2000; Huber and Wächtershäuser, 2003; Huber et al., 2003; Smith and Morowitz, 2004; Martin and Russell, 2007; Srinivasan and Morowitz, 2009; Braakman and Smith, 2013).
With these further steps required, it is clear that carbon fixation is only half the battle at the very origin of metabolism. We also need a mechanism of free energy conversion that can produce a chemically appropriate and diffusible source of free energy, plausibly a strong disequilibrium between a polyphosphate and orthophosphate. This would enable further endergonic reactions, particularly dehydration condensations, to be driven within the precellular environment, which we take to be the nanometric laminar pore spaces within the inorganic membranes (Mielke et al., 2011; Nitschke and Russell, 2013; Russell et al., 2013). Before taking up this challenge, we need to remind ourselves of how present-day life deals with the problem.
8. How Life Now Makes and Dissipates Proton Gradients
Referring back in particular to Complex 1 and ATPase—the interdependent examples of just two of the biological nanoengines operating in our own bodies—we note that the primary fuels are electrons and protons, or sodium and even potassium ions in some other organisms (Mitchell, 1976; Martin, 1992; Mulkidjanian et al., 2008; Buckel and Thauer, 2013). It is the electrons attracted exergonically in Complexes I, III, and IV toward the cell's high potential exterior that force protons to follow suit. Once the protons are strongly out of equilibrium with the cell's interior, they stream back down an extremely steep gradient through ATP synthetase, in turn driving a massive disequilibrium between a high concentration of ATP as against the pool of orthophosphate and ADP. Complex 1 and ATP synthetase are, of course, extraordinarily complicated and hence highly evolved piston-like α-helices and eccentric nanorotary engines, respectively (Abrahams et al., 1994; Yoshida et al., 2001; Efremov and Sazanov, 2011a). Do these free energy converters have ancestors that might help us identify possible missing entities occupying the first membrane that exploited exergonic geochemical gradients and reactions to drive endergonic reactions? If so, what were they built with, and how were they assembled and constructed (Branscomb and Russell, 2013)? While approximately two electrons are required to translocate four protons through Complex 1, the endergonic condensation of one molecule of ATP through ATP synthetase costs three to four protons when driven by a typical proton motive force of ∼180 mV (Mitchell and Moyle, 1965; Davies et al., 2011; Efremov and Sazanov, 2011b). Hedderich (2004) speculated that the ion-pumping nickel-iron hydrogenases from archaea and extremophilic bacteria acted as the ancestors of archaeal Complex 1 through the “addition of alternative electron-input domains, replacement of the [NiFe] center by a quinone-binding site and the addition of further membrane subunits.” Whatever its ultimate ancestry, this engine/enzyme would not have been required across the margins of the mound at the emergence of life as external protons were provided at a concentration of ∼10−5 M in the ocean as against a H+ concentration of ∼10−11 M on the inner surface—one of the telling advantages of the submarine alkaline hydrothermal theory.
While the ambient pH gradient across the walls of the hydrothermal mound releases us from a search for precursors to Complexes I, III, and IV, we do need to find precursors to what is the extraordinarily sophisticated rotary mechanism of ATP synthetase. Baltscheffsky (1971) suggested that something like a pyrophosphatase might have played a part in energizing proto-biosynthesis, an idea given experimental support by Moyle et al. (1972) (and see Baltscheffsky et al., 1999). A proton pyrophosphatase has now been characterized by Lin et al. (2012) and a sodium pyrophosphatase by Kellosalo et al. (2012). They are rather similar in structure, each consisting of 16 transmembrane helices, six of these comprising an inner flexible passageway surrounded by the remaining 10 that constitute the outer wall. It is likely that at least some pyrophosphatases can use either H+ or Na+ indiscriminately (Luoto et al., 2013). In most organisms, these pyrophosphatases are located in the membrane or in intracellular vacuoles. The entrance or exit funnels to these enzymes are around 1.5 Å (0.15 nm) across. While described as a proton and sodium transporter, respectively, these enzymes are probably reversible (Façanha and de Meis, 1998; Maeshima, 2000). In other words, with ion flow running down-gradient, they could act as pyrophosphate synthetases. If so, we expect two orthophosphates (2×HPO4 2-) to be driven into the vacuole by the proton (or sodium) disequilibria where they would condense to the diphosphate (HP2O7 3-). During the condensation, the protein undergoes an allosteric transition so that the cavity is now open to the cytoplasm, causing release of the diphosphate where it is in strong disequilibrium with respect to the two orthophosphates. Because the proton gradient would have stretched to 5 orders of magnitude while the sodium gradient was zero, we argue—pace Mulkidjanian et al. (2008)—that the ancestral ion gradient was based on protons and not sodium ions (Russell and Hall, 1997; Martin et al., 2008; Ducluzeau et al., 2014; and see Kelley et al., 2001, Table 1). Moreover, Baltscheffsky and Persson (2014) showed from genomic studies of the archaeon “Candidatus Korarchaeum cryptofilum” that the pre-LUCA evolutionary form of the membrane-integral inorganic pyrophosphatase was a proton pump (and see Elkins et al., 2008). Baltscheffsky and Persson (2014) tentatively suggested that sodium pumping was post-LUCA. The challenge before us then is to imagine a primitive functional equivalent of H+-pyrophosphatase. Thus, we consider next whether the kind of mineral structures implicated in coupling primitive carbon fixation to the ambient redox potentials could also serve as the engine for coupling the mound's proton gradient to the maintenance of an internal pyrophosphate disequilibrium.
9. Proto-Pyrophosphate Synthetase Nanoengines
As we noted above, the second great “disequilibrium conversion” challenge is to find a primitive functional equivalent of H+ pyrophosphatase. There, we made a case for metal hydroxides having contributed to the earliest inorganic membranes developed at submarine alkaline springs. The main contributors to such membranes would have been layered double hydroxides such as the hydrotalcite pyroaurite [∼Mg6Fe2(CO3)(OH)16·4(H2O)] and the mixed-valence green rust or fougèrite ∼[(FeII,Mg)2FeIII(OH-)5·CO3 2-] (Cairns-Smith, 1982; Trolard et al., 1997; Feder et al., 2005; Trolard and Bourrié, 2012). Other likely precipitates are montmorillonites, smectites, and other expandable cationic clays with comparable properties to fougèrite. Ever since Bernal's suggestion that clays would have adsorbed organic molecules on early Earth (1951, 1960, 1967), many origin-of-life researchers have considered such layered minerals and mineral clusters as being central to life's emergence (Cairns-Smith, 1982, 2008; Ponnamperuma et al., 1982; Cairns-Smith et al., 1992; Ertem and Ferris, 1998, 2000; Hill et al., 1998; Ferris, 2002, 2005; Aldersley et al., 2011; Adraa et al., 2012; Feuillie et al., 2013). In particular, Arrhenius (1986, 2003) has long championed a role for the mixed-valence iron hydroxides and other layered double hydroxides in the synthesis of prebiotic molecules, albeit from cyanide and formaldehyde. In our opinion, molecules such as cyanide are inimical to the redox state of the early atmosphere. Instead, cyanide along with carbon monoxide could have been produced in the inorganic membrane in a way similar to cleavage of tyrosine catalyzed by Fe-S cluster-mediated radical chemistry, and glycine decomposed to cyanide (along with CO) also by an iron sulfur cluster–containing enzyme (Driesener et al., 2010; McGlynn et al., 2010; cf. Peters et al., 2006; Milner-White and Russell, 2011; Kuchenreuther et al., 2013). Arrhenius and his colleagues also experimented with the potential role of anion-selective minerals as concentrators and catalysts in the chemistry of the organic phosphates (Kuma et al., 1989; Arrhenius et al., 1993, 1997; Pitsch et al., 1995; Krishnamurthy et al., 1999; and see Baliarsingh et al., 2013) (Table 2). For example, hydrotalcite catalyzes the selective condensation of aldol phosphate, a reaction along a path to the nucleic acids (Pitsch et al., 1995). Organic phosphates are also strongly adsorbed and concentrated on layered double hydroxides (Kolb et al., 1997).
Greenwell and Coveney added substantially to the knowledge base with their simulations of processes involving the intercalation of organic anions in the layered double hydroxides (Greenwell and Coveney, 2006; Greenwell, 2010; Coveney et al., 2012; Ma et al., 2012). In particular, they showed how the double layers are likely to flex and even engage in peristaltic behavior to drive diffusion of organic intercalates (Thyveetil et al., 2008; Coveney et al., 2012). Following these leads, we have suggested that fougèrite could have operated as a pyrophosphatase engine (Russell et al., 2013). Comprising the precipitates at the outer margin of an alkaline mound, ferrous hydroxide may have been oxidized to fougèrite by the nitrate and nitrite in the ocean. Opposing ferric ions in fougèrite proximal to the acidulous ocean would repel each other, opening the interlayer up to invasion by anions such as nitrate (Arrhenius, 2003). Having invaded the interlayer, nitrate would “pillar” a microgallery ∼4 Å (∼0.4 nm) high (Choy et al. 2000), leading to polaron formation, perhaps giving an opportunity to orthophosphate (diameter=4.8 Å) dissolved in the Hadean Ocean and adsorbed onto the outer faces of green rust, to be driven into the layered double hydroxide by the steep proton gradient (Ookubo et al., 1993; Arrhenius et al., 1997; Hagan et al., 2007; Yan et al., 2010; Zhou et al., 2011; Barthélémy et al., 2012; Russell et al., 2013; cf. Baikousi et al., 2013). In our model, a positive redox polaron wave propagates along the layered double hydroxide from the oxidized acidic ocean exterior toward the reduced alkaline interior, locally distorting the brucite-like layers and inducing, through charge conservation, the migration of doubly charged orthophosphates (HPO4 2-) (cf. Wander et al., 2007; Katz et al., 2012). The peristaltic drive is vectorially augmented by the direction of the proton gradient (cf. Horváth et al., 2011). The protons would be driven down-gradient by the Grotthuss mechanism (covalent transfer of a proton from one molecule to another) in a process that could extend within the membrane through interlayers in mackinawite (Agmon, 1995; Muñoz-Santiburcio et al., 2013; cf. Kellosalo et al., 2012). In these confined low-entropy two-dimensional spaces, where water activity is low, Mg2+, also from the ocean, acts to offset the charge repulsion between two monophosphates (perhaps as KPO4 2- or HPO4 2-), which are forced to condense to HP2O7 3-. The pyrophosphate is then driven peristaltically into the interior. Electrons derived from hydrothermal hydrogen at a nickel–iron sulfide center within the membrane re-reduce the fougèrite (Fig. 7). But whether phosphates can be driven by the steep proton gradient to displace interlayer carbonate requires experimental validation in light of the research by Bocher et al. (2004). So the question remains: can transmembrane redox and pH gradients totaling around 1 V abiotically drive the formation of a strong pyrophosphate versus orthophosphate disequilibrium via the endergonic condensation of mono- to pyrophosphate (cf. Paulo and Tavares, 2011)?
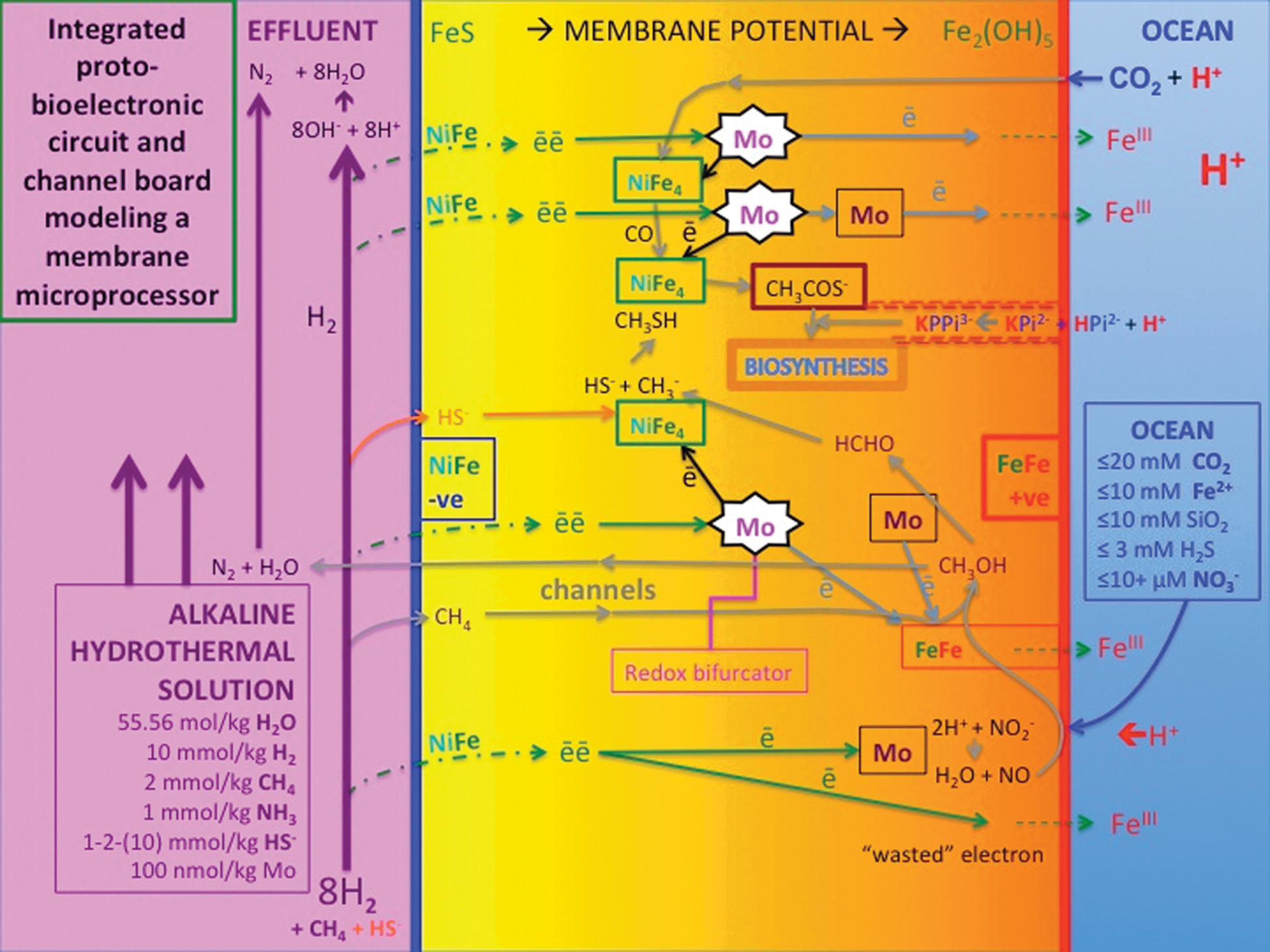
The engines that put metabolism on the road. Highly idealized deconstruction of Fig. 6a to reveal the hypothetical proto-bioelectronic and proto-bioprotonic circuitry and channels in the inorganic membrane that feed the redox and proton disequilibria to the putative carbon-fixing and pyrophosphatase nanoengines (cf. Szent-Györgyi, 1968a, 1968b; Poinsignon, 1997; Nitschke and Russell, 2013). Nickel sites in the FeS comprising the innermost zone of the inorganic membrane oxidize the hydrothermal H2, sending waves of electrons through the semiconducting iron minerals toward the oxidants bathing the membrane exterior. The resulting protons react with the hydroxyls in the hydrothermal fluid to produce more water. Some of the electrons reduce nitrate and nitrite to highly active NO and N2O at a molybdenum site stabilized by protons [e.g., [Fe2S2(MoS4)2]4- and/or [FeO(OH)(MoS4)2]3- (Itaya et al., 1987; Helz et al., 2013)], which in turn are hypothesized to oxidize methane to methanol on green rust [fougèrite, Fe2(OH)5, cf. Martinez-Espinosa et al., 2007; Starokon et al., 2011]. Two more electrons reduce MoVI atoms to MoIV, which then bifurcate; one electron is drawn along a large entropy-increasing path to FeIII in fougèrite comprising the membrane exterior, while the other takes the entropy-decreasing path against membrane potential and reduces the low potential nickel iron sulfide (e.g., Fe5NiS8). These reduced NiFe4 clusters then feed electrons to CO2 and formaldehyde in preparation for the assembly of CO and a methyl group to activated acetate, the entry point to putative carbon-fixing denitrifying methanotrophic acetogenesis. Fougèrite may also act as a proton pyrophosphatase. Note that while the characteristics of the alkaline ∼10 mM hydrothermal feed of electron donors remain stable for ≥1017 μs (i.e., ≥32,000 years, Ludwig et al., 2011), the concentrations of the oxidants are likely to vary widely in time and be the limiting entities in completing the circuit. The ocean may also have supplied many mM of SiO2 and a few mM of H2S to the alkaline hydrothermal chimneys (Mielke et al., 2010; Shibuya et al., 2010). Based on Nitschke and Russell (2013) and Russell, Nitschke and Branscomb (2013).
10. The Putative Hybrid Nanoengine
A model isolating and integrating the suggested mechanisms—the carbon-fixing and pyrophosphate synthetase nanoengines—imagined and invoked to resolve the disequilibria acting across the first inorganic membrane is presented in Fig. 7. Here, in the spirit of Szent-Györgyi (1968a, 1968b), we seek to combine electron conduction and redox bifurcation/confurcation with vectorial chemical transport in fluid channels such that the putative mechanocatalytic nanoengines at their confluence drive the “hard” reactions of carbon fixation and subsequent organic condensations (cf. Mitchell, 1975; Huang et al., 2012). The aim of this proto-bioelectronic and proto-bioprotonic circuit diagram is to provide a basis for experimental investigations. The system combines the mechanisms of a fuel cell directly with the two hypothesized engines of proto-metabolism, all situated within the membrane (Russell, 2007; Branscomb and Russell, 2013; Russell et al., 2013; Barge et al., 2014).
11. The Organic Takeover
Assuming we are on the right track, it must be admitted that a fougèrite H+-pyrophosphatase does not resemble the bundle of transmembrane helices comprising H+/Na+-pyrophosphatases as elucidated by Lin et al. (2012) and Kellosalo et al. (2012). So what could have been the natural intermediate step between the mineral motor and the protein? Milner-White and Russell (2005, 2011) suggested that the organic takeover involved just those molecules that are most exergonic to produce—the amino acids and uncoded peptides (Fig. 8) (Amend and Shock, 1998; Amend and McCollom, 2009). In this light, it is not so surprising that Ferris and his collaborators (1996) have generated long peptides on the layered mineral illite. They imagine mineral surfaces covered by polymers 50 residues long or more on early Earth. Spontaneously precipitated layered minerals, such as mackinawite (∼FeS), fougèrite (∼Fe2(OH)5), and perhaps tochilinite (6FeS0.9·5(Mg,Fe)(OH)2), would seemingly offer attractive surfaces and interlayers for ligation of such multi-dentate ligands that comprise the backbones and side chains of peptides (Zolensky, 1984; Beard, 2000; Mielke et al., 2011; McGlynn et al., 2012; Kang et al., 2013; cf. Shanker et al. 2013). Indeed, McGlynn et al. (2012) demonstrated that the addition of peptides or RNA to the alkaline solution modifies the elemental stoichiometry of the chimney precipitates to the effect of increasing the oxygen-to-iron ratio and likely that of the hydroxide proportions of the membrane. Membrane stability is also increased in proportion to polymer length (Bormashenko et al., 2006). It is also likely that spontaneously precipitated mackinawite (FeS) and fougèrite (∼Fe2(OH)5), the assumed “objets trouvés” for engine components, may have had some templating control over subsequent precipitation as the inorganic compartments were forced to grow during hydrothermal inflation (Russell et al., 1994; Baymann et al., 2003; Cairns-Smith, 2008; Greenwald and Riek, 2012; Branscomb and Russell, 2013; Kang et al., 2013; Nitschke et al., 2013).
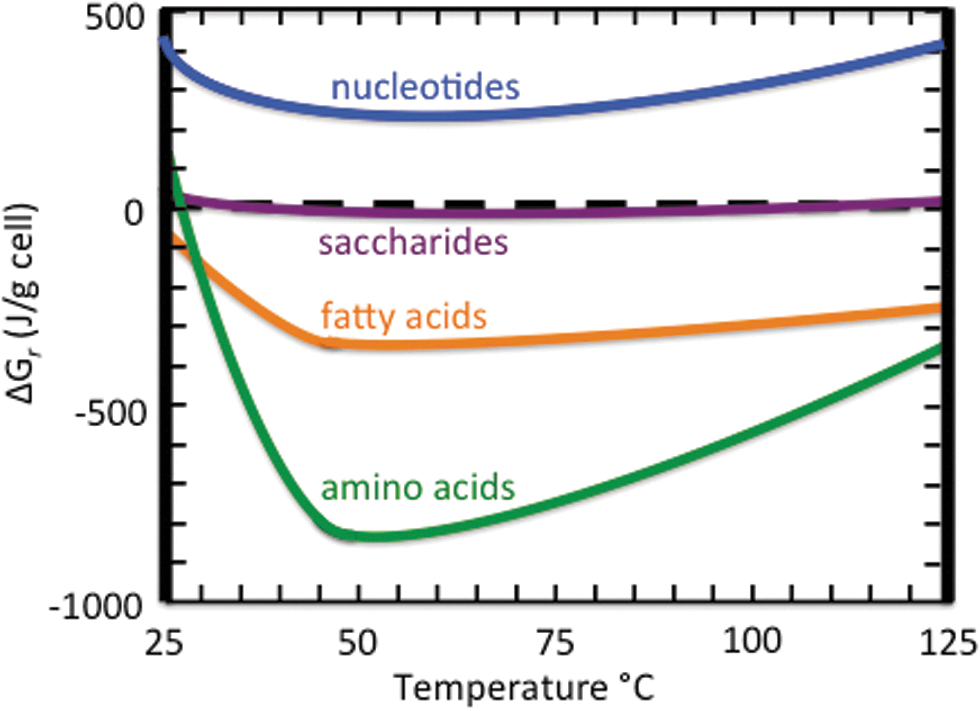
Calculated Gibbs energies of reaction for total cellular constituents assumed to be generated on mixing of Hadean seawater with a Lost City–like hydrothermal fluid as a function of temperature (Amend and McCollom, 2009). Fatty acid synthesis is exergonic at all temperatures, whereas amino acid synthesis is exergonic above ∼27°C, and strongly so between 30°C and ∼90°C. Amines and saccharides are potential products close to equilibrium. The synthesis of nucleotides, in strong contrast, is endergonic at all temperatures for all mixtures (Amend and McCollom, 2009, Tables I, IV, and V). This is why life does not live on RNA/DNA alone, could not have begun relying on these molecules in these conditions, and why these polymers could never clog up the cell. Redrawn from Fig. 2 in Amend and McCollom (2009), with permission.
With increase in proportion of peptide to layered double hydroxide and sulfide, we imagine an organic takeover of the membrane by amyloid in an evolving process of what might be called “minerobiolization” (Russell, 1991; Milner-White and Russell, 2005). Minerobiolization with amyloidal peptide templated upon these mineral clusters would markedly improve the membrane structure, durability, and discrimination (Zhang et al., 1993; Chernoff, 2004; Carny and Gazit, 2005; Russell and Hall, 2006; Milner-White and Russell, 2008; McGlynn et al., 2012). Indeed, such uncoded peptides are known to be self-recognizing and thereby to arrange themselves as parallel α-sheets (Armen et al., 2004; Milner-White et al., 2006; Hayward and Milner-White, 2008). The α-sheet can spontaneously convert into the somewhat more stable and nontoxic β-sheet of insoluble proteins known as amyloid by the process of peptide plane flipping (Milner-White et al., 2006; Hayward and Milner-White, 2008). As either α- or β-sheets, they would have been rapidly self-propagating and temperature-resistant and could have comprised the first organic membranes and cell walls (Zhang et al., 1993; Milner-White and Russell, 2005; Childers et al., 2009, 2010; Maury 2009; Greenwald and Riek, 2010; Goodwin et al., 2012). As remarked by Greenwald and Riek (2012), amyloidal peptide may therefore be said to propagate information in the form of conformation (and see Maury, 2009; Li et al., 2010; Goodwin et al., 2012), and amyloid-like fibrils have been shown to replicate (Takahashi and Mihara, 2004). Moreover, peptides are much more versatile with respect to the requirements of basic metabolism than any other of the biotic molecules, comprising, as they do, the main structures of the enzymes, proteins, and even the cofactors as well as much of the membrane. They are much stronger than lipids as evidenced by their role in cell walls—in weblike structures strong enough to contain turgor pressures (Kandler and König, 1998; Desvaux et al., 2006). Thus, they have the potential to act at the emergence of life in such roles as cell membrane/cell walls, and their backbones can sequester metals and phosphate without reliance on side-chain order; that is, they operate as nests for the inorganic active centers of enzymes comparable to the P-loop without the requirement for coding (Table 2) (Milner-White and Russell, 2005, 2011; Childers et al., 2009; Bianchi et al., 2012; Hong Enriquez and Do, 2012). However, various short peptides with particular sequences useful as binding nests may have afforded reproducible protein structures, some of them oligomeric, through self-assembly (Lupas et al., 2001).
Given a reservoir of amino acids, amyloid grows exceedingly rapidly (Collins et al., 2004; Goodwin et al., 2012). Many of the amino acid residues comprising amyloid proteins are “nitrogen-rich,” for example, asparagine, glutamine, and arginine, although serine is also common. The sequences in which they occur are largely immaterial to amyloid growth (Ross and Poirier, 2005). Indeed, the amyloid structure can maintain dynamic contact even as the sequences of these particular amino acids are shuffled, thus giving opportunities for selection and survival based on function. Such analog behavior may be thought of as Lamarckian molecular evolution guided by attractor states (Kuhn, 1972; Corliss, 1986; Hengeveld, 2007; Halfmann and Lindquist, 2010; Goodwin et al., 2012). At this stage, it is the main-chain CONH atoms, rather than the side chains, that dominate function. Were the main chains at the edges of blocks to be juxtaposed to blocks of similar charge, the membrane would remain semipermeable, and channels composed of α-sheet peptides could encourage the transport of protons across the membrane (Milner-White and Russell, 2008). Whether such transport could be gated and coupled to pyrophosphate synthesis is ripe for research (e.g., Hansen and Poulsen, 1999; Dong, et al., 2006; Childers et al., 2009).
12. The Second Transition across the Darwinian Threshold—the Guidance System
As explained by Goodwin et al. (2012), the kind of thermodynamically driven evolutionary process described in the last section falls short of digital Darwinism. So how might the Darwinian threshold be crossed (Woese, 2002)? What are the steps leading toward and across this threshold toward a nucleotide guidance system? First, there is the problem of synthesis (Fig. 5). While cyanide has often been appealed to as explanation for the facile production of certain nucleic acid bases (Sanchez et al., 1967), abiotic HCN has not been recorded in moderate-temperature alkaline submarine springs (e.g., Lang et al., 2010). And while it is also true that nucleotides have been generated in the laboratory (Powner et al., 2009), the various conditions imposed are particular, discontinuous, and result in complex mixtures, and eventually face the “asphalt problem” (Benner et al., 2012). Instead, given that the substrates for the biosyntheses of the nucleobases are aspartate, glutamine, and glycine, with contributions too of carbon dioxide, ammonia, formyl phosphate, and glyceraldehyde-3-phosphate (Berg et al., 2006)—all molecules that are likely to have been present or generated in the hydrothermal mound—it seems that this is an occasion for “top-down” inference to be entertained (Martin and Russell, 2007). Add to this observation that effective phosphorylation mechanisms and rapid mineral-assisted condensation of aldehyde phosphates have been demonstrated to form tetrose and pentose phosphates in high yield, the emergence of the simple nucleic acid enzymes may be more facile than generally assumed (Pitsch et al., 1995; Krishnamurthy et al., 1999; Martin and Russell, 2007).
Supposing this to be the case, we follow White's (1976) suggestion that the first nucleic acid enzymes involved in intermediary metabolism had limited substrate specificity and would catalyze classes of reactions rather than specific metabolic reactions. Yarus (2011) suggested that such early cofactors could take part in their own replication. In particular, nicotinamide adenine dinucleotide phosphate (NADP) and flavin adenine dinucleotide (FAD) might replicate through base pairings with adenine and uracil. Yarus portrays a collective of these molecules as the “initial Darwinian ancestor.” Strung together with other nucleic acid bases, the RNA polymers could behave like viruses living off metabolic energies, though partially paying back the system with some catalytic activity (Koonin and Martin, 2005). However, the synthesis of a negatively charged phosphoribose backbone on which to hang a variety of nucleobases is beyond the limits of spontaneous unguided self-organization. Considering the work of Goodwin et al. (2012), one possibility is that the first nucleobase backbones were produced through the condensation of amine and aldehyde to produce cationic alkyl iminium backbones. This is a process more easily imagined in the amyloidal milieu. On this tack, Braun et al. (2011) suggested that the earliest replicating entities were amyloid–nucleic acid fibers produced through electrostatic interactions between nucleic acids and peptides. The fibers would grow rapidly through the addition of monomers, generally alternating between hydrophobic and hydrophilic residues as influenced by replicating nucleic acids (Collins et al., 2004; Braun et al., 2011).
Perhaps the Yarus mechanism evolved from these first putative amyloidal peptides and membranes (Zhang et al., 1993). ATP drives all these condensations today (Goodwin and Lynn, 1992; Benner, 2004), but at the emergence of life, pyrophosphate—presumed to have been produced in proto-H+ pyrophosphatases—may have fulfilled the same role (Yamanaka et al. 1988; Russell and Hall, 1997; Yamagata and Inomata, 1997; Baltscheffsky et al., 1999; Baltscheffsky and Persson, 2014). Moreover, there was likely a surplus of just the right stuff in the vicinity of amyloidal membrane growth. For example, it may be that ammonia could have been generated on site through the reduction of nitrate and nitrite by fougèrite to add to the CO2 from the ocean and formyl phosphate synthesized within the membrane (Hansen and Koch, 1998; Martin and Russell, 2007).
Once “invented,” the major achievement of the RNA world was the development of a pocketlike primitive ribosome—another entropy trap—with the peptidyl transfer center at its heart (Sievers et al., 2004; Hsiao et al., 2009; Fox et al., 2012). This ribosome could generate short uncoded peptides more effectively than those probably generated on the mineral surfaces and interlayers comprising the compartment walls of the mound. And once generated, these peptides could further stabilize and protect the primitive ribosome from which they emerged—the kind of positive feedback behavior required of emergent life. That peptides preceded a nucleic acid–based world is suggested too by the presence of an 18-mer unstructured glycine-rich peptide in the modern peptidyl transfer site where it carries out the same function: the stabilization of ribosomal RNA (Nissen et al., 2000; Hsiao and Williams, 2009; Hsiao et al., 2009; Belousoff et al., 2010). Indeed, Hsiao et al. (2009) suggested that the peptide is a molecular fossil and an ancestor of the ribosomal proteins.
The plausibility of this sketch of how the evolutionary transition was effected might seem to face the same problem as the organic soup hypothesis, that is, concentrations of polymers way below a required critical mass for interaction and recombination. However, research by the Braun group illustrates how nucleic acids might be concentrated in such a complex system (Mast et al., 2013). Though in their experiments they used DNA rather than the less stable RNA, these authors demonstrated how a thermal gradient across inorganic compartments of a dimension expected in a hydrothermal mound could drive a convective polymerase chain reaction to produce longer polymers, and how once formed these are driven to, and concentrated in, the coolest region of the compartment through thermal diffusion (Baaske et al., 2007; Mast and Braun, 2010; Mast et al., 2012).
13. Discussion
In the present discussion, we have concentrated on how metabolism might have begun from scratch and outlined our reformulated transdisciplinary theory for an autotrophic emergence of life in a submarine alkaline mound. This theory abides by the rules of thermodynamics, pays due consideration to initial conditions on wet rocky worlds, and looks to the Last Universal Common Ancestor (LUCA) for correspondence. It explains the polarity of the cell, alkaline and reducing inside, acidulous and oxidizing outside; the vector of the proton motive force and the universality of its use; the similarity of mineral and active protein center structure; and the commonalities of certain “proto-biochemical” reactions and metabolic pathways and syntheses (Nitschke and Russell, 2013).
Under this view, life emerged quickly in the confines of the founding mound within the duration of the vitalizing low-entropy hydrothermal feed. The first steps were the steepest: reduction of carbon dioxide, oxidation of methane, and the production of pyrophosphate very far from equilibrium with its orthophosphate source. These steps necessitated close physicochemical couplings to exergonic reactions (Branscomb and Russell, 2013). This is where we find ourselves converging with another well-established origin-of-life research field, one that championed mechanistic roles for layered double hydroxides and smectites (expandable clays) at the emergence of life, but with the difference of placing them in, and as components of, the inorganic membrane (Arrhenius, 2003; Meunier et al., 2010). After all, the affinities are now well established between the structures of certain mineral clusters comprising the precipitate membranes—mackinawite, greigite, violarite, and fougèrite—with the most ancient metalloenzymes involved in autotrophy (Fig. 4) (Nitschke et al., 2013). What is not so clear is how the flexible motions of the transmembrane bundles of α-helices involved in so many of life's nanoengines that couple exergonic with endergonic reactions functioned in a mineral world before the emergence of proteins (Childers et al., 2012; Branscomb and Russell, 2013). Here, appeal is made to the layered double hydroxide fougèrite (green rust)—a likely precipitate on the outer margins of an alkaline hydrothermal Hadean mound—to act in this capacity, albeit inefficiently (Mielke et al., 2011; McGlynn et al., 2012; Russell et al., 2013). Of course, it is true that their haphazard architecture would have made them unreliable at best. Nevertheless, presuming that these materials engineered the first steep steps to metabolism, their inherent autocatalytic behavior would have seen to their improvement through a rapid organic takeover. Once the initial steep carbon fixation steps were overcome, the synthesis of carboxylic, amino, and fatty acids as reduced hydrothermal solutions met Hadean Ocean in a precipitate mound, was generally exergonic, so much so as to suggest basins of attraction to particular metabolic pathways and, eventually, cycles where evolution was largely “analog” in nature (e.g., Corliss, 1986; Amend and McCollom, 2009; Goodwin et al., 2012). Even so, catalysts and enzymes, initially inorganic but eventually protein-based, were required to expedite and control the rates of reactions.
The phase transition from aqueous geochemistry to metabolism through prebiotic physicochemical reactions, like all symmetry-breaking and irreversible events, would have been so rapid as to call into question the idea—rooted in the “organic soup” concept—that prebiotic organic molecules be considered as targets for “pre- or proto-life” detection (Lane et al., 2010; Russell and Kanik, 2010). Indeed, as we have argued here and previously, the main prebiotic molecules were CO2, CH4, H2, HPO4 2-, P4O10, NH3, NO, FeS, Fe3S4, Ni, Co, Mg, Mo, W, and the like (Nitschke and Russell, 2009). Evidence for this is provided by the fact that the biosphere is overall autotrophic and appears to have been so from life's onset (Fuchs 1989, 2011; Berg et al., 2010; Schoepp-Cothenet et al., 2013). Efficiencies of inorganic precipitates as catalysts and nanoengines were likely improved significantly when replaced by amyloidal peptide.
The next step on the way from metabolism to life is the sorting out of a muddle of possible reaction manifolds (Cabello et al., 2004)—the taming, constraining, and guiding of reactions involved in various possible natural attractor states, for example, those otherwise controlled by physicochemical thermodynamics such as the reduction of nitrate to nitrite, nitric, and nitrous oxides through to ammonia. This is when the RNA world begins to make a contribution. Heterocyclic nitrogen compounds formed in an amyloidal milieu are an imperative even for an uncoded early metabolism (Nissenbaum et al., 1975; Martin and Russell, 2007). Strung together on a peptide-like backbone, the metabolism involved in the cell's free energy economy could be guided and evolve. Once an RNA world and eventually a ribosomal world kicked in, favorable reactions would be sorted out in metabolic networks more responsive to the environment (Fig. 3) (Berthoumieux et al., 2013; de Lorenzo, 2014). An intriguing aspect of the onset of coding is the demonstration by Amend and McCollom (2009) of the relatively high thermodynamic costs of nucleotide synthesis in submarine hydrothermal conditions (Fig. 8). Not only is it now evident from energetic considerations that metabolism had to have preceded coding, it is also clear that the first protocells must have been more than sacks of RNA—that once introduced, RNA and DNA could never increase to the point that they would clog up the internal workings of protocells. Indeed, polynucleotides had to pay their way through reciprocal arrangements and collaboration with amino acids to produce the first coded proteins, and in so doing, cross the Darwinian threshold to cellular life (Woese, 2002). Of course, this “thermodynamic cost” refers to unavoidably endergonic reactions. Indeed, it seems clear that no replicative and/or “coding” system could operate based only on exergonic reactions and quasi-equilibrium chemistry. Even such a strong advocate of “replication first” as Addy Pross has argued “the replication reaction must be effectively irreversible. That condition, in turn, means the system must be maintained in a far-from-equilibrium state” (Pross and Pascal, 2013). Here, the key point is that by the second law endergonic reactions can only take place—and non-equilibrium states thereby be maintained—if they are mechanistically coupled to stronger exergonic reactions so that the coupled pair, acting thermodynamically as a single, unitary process, is itself exergonic. This conclusion raises the question: what were the necessary sources of driving free energy, what would it take to produce and maintain them, and very nontrivially, how could they be thermodynamically coupled to the needed endergonic reactions? Our position is that some primitive prebiotic metabolism, roughly of the type here proposed, must have been in place to meet these fundamental and inescapable thermodynamic requirements. It is in this rather limited sense, and for this reason, that we prefer a “metabolism first” model of the emergence of life.
Although the disequilibria that drove life into being are easily imagined, the nature and operations of the first engines assumed to have negotiated the steep endergonic barriers to autotrophic metabolism are not. Moreover, it is to be expected that any such engines, and their housings, would have been inefficient and unreliable at best. Had it not been for the large numbers of alkaline hydrothermal systems likely operating on early Earth, each with billions of inorganic membranous compartments, we might be tempted to entertain Monod's pessimistic surmise that life is an accident and we are alone in the Universe (Monod, 1972). But with this redundancy of opportunity—that there were so many chemical systems in so many dispositions in so many hydrothermal mounds over so much time—life's emergence appears inevitable. We imagine that the proto-metabolic free energy converters approximating the ones described here would easily ratchet up to the next level of complexity, that is, to a coded world guided by RNA. Once RNA-guided evolution began to make its contribution, such entities would be capable of taking the many steps, through the LUCA, to the complex life that constituted our biosphere as it was 3.8 billion years ago (Rosing, 1999).
Evolutionary retrodictions and postdictions addressing life's emergence restrict history-based inquiry to surviving prokaryotes, their constituents, their particular phylogenies, and the “calculated” geophysical and geochemical conditions that occurred around 4 billion years ago on our own planet. Much information on the initial conditions that drove life into being has been lost to tectonic process and biochemical weathering, while the progress of earliest life has been lost to extinctions as well as to the absence of evolutionary information before the evolution of the ribosome. However much we surmise over the mechanisms that enabled life to emerge, and how it spread to environments where appropriate disequilibria obtained and obtain here on Earth, astrobiological research has the potential to add dimension to our understanding of how, and under what conditions, life emerged from a low-entropy feed to play its part in increasing the overall entropy of the Universe (Boltzmann, 1886; cf. England, 2013). It is possible that different physicochemical disequilibria, different high potential electron acceptors, and environmental challenges on other worlds might have called for somewhat different metabolic pathways and strategies to be entertained by emergent life elsewhere. For example, in the absence of high potential electron acceptors, life may have eventually emerged via the reductive acetyl coenzyme A pathway after all (Russell and Martin, 2004). Whatever the possibilities, the various interdependent physical and chemical conditions to be revealed by the exploration of, for example, Mars, and eventually of Europa and Enceladus and the exoplanets, can only help our general understanding and perhaps force a rethinking of how life might have emerged here on Earth. Supposing life did begin in an alkaline submarine mound, we may surmise that on our own planet, needing surfaces as substrate, it would have expanded into, or been carried along and just below, the ocean bottom, eventually to inaugurate the deep biosphere (Roussel et al., 2008; Suzuki et al., 2013). Starved of electron acceptors, microbes may then have been forced to adopt methanogenesis as a means of losing electrons. Geochemical conditions in the mafic and ultramafic crust hosting the present deep biosphere, even when obducted, can have hardly changed in the last 4 billion years and thus might provide indications of what the oldest inhabitants of the planet were like (Thauer et al., 1977; Morrill et al., 2013).
However, the immediate and mutually dependent experimental challenges are (i) to recreate the putative denitrifying methanotrophic acetogenic pathway and its extension toward the reductive, though incomplete, version of a citric acid cycle, presumably necessitating electron bifurcation involving molybdenum, and (ii) to demonstrate that a steep proton gradient across a fougèrite-bearing membrane will drive a strong pyrophosphate disequilibrium (Smith and Morowitz, 2004; Martin and Russell, 2007; Branscomb and Russell, 2013; Nitschke and Russell, 2013; Russell et al., 2013). Both these challenges require experimentation in sterile conditions under the kind of high pressures that would be met at depth in the Hadean Ocean.
14. Unresolved Issues
The recognition (i) that all life absolutely and fundamentally depends on driving strongly endergonic, thermodynamically “uphill” reactions; (ii) that, by the second law, this in turn requires that each such reaction be “coupled,” as the driven partner, to a (stronger) exergonic reaction, acting as the driving partner; and (iii) that such reaction coupling may require the mediation of a molecular “mechanocatalyst engine” specific to the reactions being coupled has forced the realization that some simpler mechanocatalysts might have been on hand to engineer life's origin. We have alighted on green rust (fougèrite) as one of the most plausible mineral precipitates at the membrane that, within its interlayers, is potentially capable of the kind of physical articulation allied to redox and ionic gradients likely to result in particular products and their transport (Arrhenius, 2003). The demonstration by Hansen and Koch (1998) of the reduction of nitrite to ammonia through the partial oxidation of fougèrite prompts the question whether that same mineral, suitably doped, could concomitantly oxidize methane to methanol and formaldehyde. To date, such abiotic oxidation has only been achieved with surface oxygen on an α-cationic Fe2+ site within the cage of a synthetic zeolite (see Kachurovskaya et al., 2004, Fig. 4). Experiments at high pressure interacting fougèrite with methane, hydrogen, and nitrite across an inorganic membrane are called for. The aim in this case would be to test whether fougèrite could act as the inorganic analogue of the methane monooxygenase enzyme and, at the same time, act as a proton-pyrophosphate synthetase (Huang et al., 2013; Russell et al., 2013).
We are also lacking a good model for the first redox bifurcation engine that we hold to be vital for entry to C1 metabolism. A difficulty is that it is not entirely clear how two-electron bifurcating engines work in metabolism today. Nitschke and Russell (2011) invoked strongly crossed-over individual redox potentials dependent on local pH gradients involving a molybdenum atom (or tungsten at high temperature). However, the detailed molecular properties of the mechanism by which these shifts would be produced mechanistically to ensure that the exergonic (i.e., driving) one-electron reduction only takes place if, “at the same time,” the endergonic (i.e., driven) one-electron reduction takes place remain to be worked out. Mitchell (1975) involved the concept of electron bifurcation (Wikström and Berden, 1972; Wikström and Saari, 1977) in his chemiosmotic theory, and we are alert to the interdependency of redox bifurcations and protonations (and see Buckel and Thauer, 2013). Jacques et al. (2014) offered models of how protons are involved in the activation of nitrate reductase, a di-pterin with a molybdenum cofactor. The molybdenum is invariably situated in an exposed position at the axis of a symmetrical di-pterin and is reduced as the pterin proximal to an [4Fe-4S] center is protonated. The onus is on us to suggest an inorganic cluster that might be protonated as molybdenum is reduced. In an inorganic membrane, molybdenum could be present in sulfide or oxide clusters. By reference to operations of ferredoxin (FdL), we tentatively speculate that of all known entities, linear clusters such as [Fe2S2(MoS4)2]2-/4- or [FeO(OH)(MoS4)2]1-/3- might be reduced and protonated as hydrogen is oxidized on a nickel-bearing mackinawite (Helz et al., 2013). Molybdenum can also occur as iron-MoIV-oxide domains within green rust (Boucherit et al., 1991). However, such an association has not been observed in modern metalloenzymes. In theory, the entropy gained as one electron from a molybdenum cluster reduces nitrate to nitrite more than offsets the reduction of entropy entailed in the second electron's loss to the recalcitrant CO2. This potentially might be made possible by a reorientation of a putative molybdenum cluster as the two protons are drawn one by one to the alkaline interior. In any case, the challenge is to realize an abiotic system in which the inherently exergonic one-electron substrate oxidation serves to gate the one-electron endergonic one and which at the same time prevents both electrons from taking the easy exergonic ride downhill. Whether this can be achieved through sterical (mechanocatalytic) or purely electrochemical means is controversial even among the authors of this article.
15. Conclusions
(1) Immediately prior to the onset of life, icy wet rocky worlds would have the same electrochemical polarity as do cells, that is, a reduced and basic interior (cf. cytoplasm) and a relatively oxidized and slightly acidic exterior (cf. periplasm). Both are electrochemical cells with outputs or inputs of up to ∼1 V (Russell and Hall, 1997; Nitschke and Russell, 2013). The likely factor limiting the first metabolizing system is the proximal availability of high potential electron acceptors such as nitrite, nitrate, and ferric iron.
(2) Hydrothermal mounds at submarine alkaline springs produced through serpentinization would have focused and miniaturized Earth's polarity and configuration, presenting hydrothermal hydrogen, methane, ammonia, and hydrogen bisulfide to nitrate/nitrite, carbon dioxide, and ferric iron in the metal-bearing acidulous carbonic ocean for their controlled irreversible interactions across the spontaneously precipitated inorganic membranes (Russell and Hall, 1997; Nitschke and Russell, 2009). This would have set the table with strong redox disequilibria to drive C fixation and a strong and correctly oriented proton gradient disequilibrium to drive the generation of an ATP-like diffusible source of free energy in the form of pyrophosphate versus orthophosphate.
(3) The first steps to carbon fixation, one reductive and one oxidative, are the very steepest—fixing C1 carbon from CO2 pumped out of volcanoes and dissolved in the ocean, and CH4 carried by the alkaline hydrothermal solution. Both processes involve free energy converting catalysts to perform redox bifurcations and exploit the proton motive force (Nitschke and Russell, 2013; Russell et al., 2013). The “organic soup” concept fails to account for the presence of these mechanisms in the LUCA (Lane et al., 2010).
(4) The first free energy–converting nanoengines or mechanocatalysts that enable and drive C1 metabolism up these steep steps are argued to involve (i) iron nickel sulfides as hydrogenases and dehydrogenases, (ii) layered double ferrous/ferric oxyhydroxides (fougèrite) as methane monooxygenase and as a proto-pyrophosphatase, (iii) redox bifurcation agents involving individual atoms of molybdenum complexed in the sulfides and/or fougèrite constituting the first membranes (Boucherit et al., 1991; Helz et al., 2013). With the involvement of these inherent catalysts, emergent life transcends mere chemistry and geochemistry as well as mere catalysis (Nitschke et al., 2013). Activated acetate is the first product (Huber and Wächtershäuser, 1997), and the revised pathway is denoted by the descriptor denitrifying methanotrophic acetogenesis (Nitschke and Russell, 2013). The pathway is assumed to eventually extend to succinate and glyoxalate through hydrogenations, carboxylations, and condensations driven by ATP. Aminations and further condensations produce amino acids, peptides, and ultimately nucleic acids and polymers (Martin and Russell, 2007).
(5) The organic takeover from the mineral precursors is facilitated by the surface structure of amyloidal peptides between 6 and 10 amino acids long that behave as nests for inorganic clusters. Together these do the work of proto-enzymes (Milner-White and Russell, 2008, 2011; Bianchi et al., 2012; Hong Enriquez and Do, 2012). Propagating rapidly, amyloid can substitute for the inorganic membrane (Zhang et al., 1993). The amino acids might interact with ammonia, formyl phosphate, and carbon dioxide to produce the nucleobases that initially function as coenzymes (Berg et al., 2006; Martin and Russell, 2007). Strung together on a peptide-like backbone, these RNAs could act as the first transportable genes (Koonin and Martin, 2005; Goodwin et al., 2012).
(6) We argue that, once the initial hard steps of free energy conversion were scaled by the available free energy–converting engines, further biosynthetic pathways involving thiolations, condensations, phosphorylations, and aminations are all exergonic and relatively straightforward. The same goes for the pathways and cycles themselves, each becoming more cost-effective as enzyme efficiencies and peptide and protein motors are incorporated into the individuated systems or cells and gifted, through horizontal gene transfer, to their neighbors.
(7) The only opportunity of egress from the mound was to the ocean bottom and crust where, although concentrations of nutrients and fluxes of electron donors' and acceptors' energy were lower, they were otherwise similar (Takai et al., 2004; Russell and Hall, 2006; Preston et al., 2011; Brazelton et al., 2012; Ménez et al., 2012; Takami et al., 2012; Orcutt et al., 2013). And while in these reduced circumstances carbon turnover was likely orders of magnitude slower than at the vent, the unsurpassed economy of the acetyl coenzyme A pathway would have been adequate to support acetogenic bacteria (Russell and Martin, 2004; Martin and Russell, 2007; cf. Lever, 2012; Hoehler and Jørgensen, 2013). The loss of electron acceptors at the base of this deep biosphere may have forced the emergence of methanogenesis as the mechanism of electron loss.
(8) The evolutionary path from the hydrothermal hatchery to the complex biochemical world of today transcended geochemical and analog operations such that it attained the heterarchical metabolic and digital processing that is life as we know it (Goodwin et al., 2012; Nitschke and Russell, 2013). The effects were to (i) access hitherto untapped but metabolically commensurate and appropriate free energies, including solar free energies, which allowed leaps to higher levels of complexity (Baltscheffsky, 1997; Mulkidjanian and Junge, 1997; Ferry and House, 2006; Russell, 2007; Russell et al., 2008; de Lorenzo, 2014); (ii) find ways of increasing the efficiencies of free energy conversion to the ultimate consequence of increasing the overall rate of entropy production, for example, by endosymbiosis of mitochondria and chloroplasts (Allen, 2003; Lane and Martin, 2010).
(9) Evolution subsequent to the LUCA operated as a search engine for free energies and materials fundamentally related to those that got life going in the first place, though at lower flux until photosynthesis emerged: life forms that, notwithstanding their diversity, maintain deep and common characteristics as their shared inheritance from how—and in what context—metabolism began. In fundamental terms, how it was done in the beginning is how it has ever thereafter been done over the succeeding ∼4.4-billion-year journey from that inanimate geochemical world (Russell, 2007).
(10) Hydrogen from water—released geochemically and subsequently photosynthetically through mineral-like Fe±Ni and Mn4Ca active centers, respectively—continued as the dominant source of electrons, while carbon dioxide became the main primary source of carbon and oxygen the dominant electron sink (Russell and Hall, 2006; Russell et al., 2008).
(11) Although the synthesis of most biomolecules is exergonic, the fact that the biosynthesis of the nucleotides is highly endergonic is fortuitous in that there is no tendency for them to clog up the early cytoplasm—RNA and DNA always have to pay their way (Amend and McCollom, 2009).
(12) In considering habitability and the potential for life elsewhere in the Solar System and beyond, the physical and chemical disequilibria that obtain on wet icy rocky worlds, and the various processes that might relieve them, need to be established (Lovelock, 1965; McCollom, 1999; Zolotov and Shock, 2003; Hand et al., 2009; Stüeken et al., 2013). If life's origin is ultimately coupled to geophysical convection in a particular geochemical context, one should be able to make predictions about life's likelihood on a planet or moon of interest from application of coupled chemical and fluid/geodynamical modeling, and from the availability of key feedstocks, thus accounting for other planetary energetic drivers, for example, tidal and radiogenic heating, solar wind interactions, magnetic dynamos—appropriate to the object in question. In this way, it might be possible to predict whether the required gravitational, thermal, and chemical gradients are likely to have been forced, or are being forced still, beyond the critical values needed for the drive to life.
Footnotes
Acknowledgments
We thank John Allen, Ariel Anbar, Jean Bédard, Billy Brazelton, John Callas, Ivria Doloboff, Kevin Hand, Mickey Honchell, Michael Malaska, Randall Mielke, Ken Nealson, Yuk Yung, and the Thermodynamics, Disequilibrium and Evolution (TDE) NAI Focus Group for discussions, and two anonymous reviewers for their suggestions. The research described in this publication was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration, with support by a NASA Exobiology and Evolutionary Biology award (NNH06ZDA001N) and supported by the NASA Astrobiology Institute (Icy Worlds). E.B. is supported by the NASA Astrobiology Institute (Universal Biology).
Abbreviations
ADP, adenosine diphosphate; ATP, adenosine triphosphate; LUCA, Last Universal Common Ancestor.