
Abstract
Significance:
The redox balance of cells provides a stable microenvironment for biological macromolecules to perform their physiological functions. As redox imbalance is closely related to the occurrence and development of a variety of diseases, antioxidant therapies are an attractive option. However, redox-based therapeutic strategies have not yet shown satisfactory results. To find the key reason is of great significance.
Recent Advances:
We emphasize the precise nature of redox regulation and elucidate the importance and necessity of precision redox strategies from three aspects: differences in redox status, differences in redox function, and differences in the effects of redox therapy. We then propose the “5R” principle of precision redox in antioxidant pharmacology: “Right species, Right place, Right time, Right level, and Right target.”
Critical Issues:
Redox status must be considered in the context of species, time, place, level, and target. The function of a biomacromolecule and its cellular signaling role are closely dependent on redox status. Accurate evaluation of redox status and specific interventions are critical for the success of redox treatments. Precision redox is the key for antioxidant pharmacology. The precise application of antioxidants as nutritional supplements is also key to the general health of the population.
Future Directions:
Future studies to develop more accurate methods for detecting redox status and accurately evaluating the redox state of different physiological and pathological processes are needed. Antioxidant pharmacology should consider the “5R” principle rather than continuing to apply global nonspecific antioxidant treatments. Antioxid. Redox Signal. 34, 1069–1082.
Introduction
Redox regulation is closely related to cell homeostasis. The redox balance of cells provides a stable microenvironment for various biological macromolecules to perform their normal functions. Redox-dependent post-translational modification of proteins is an important switch to regulate protein function and cell signal transduction (74). The theory of free radical in aging points out that oxidative stress caused by a large number of free radicals is an important cause of aging (35). Oxidative damage to biomacromolecules or disorderly oxidative modification of proteins is also related to the occurrence and development of various diseases, including cancers, diabetes, and neurodegenerative diseases. In recent decades, therefore, antioxidant supplementation has been a strategy in nutrition, anti-aging, or disease treatment.
Even though emerging research evidence has suggested that antioxidants (such as vitamin C) can reduce oxidative damage and subsequently increase healthy longevity (9, 60, 71, 97), a number of experiments or clinical trials have shown harmful effects (57, 101). For example, it has been shown that high doses of synthetic antioxidants, when applied to proliferating cells, cause DNA damage and induce premature senescence (57). A clinical study on health in older women also finds that dietary vitamin (multivitamins and vitamin B6) and mineral supplements may be associated with increased mortality (72). Similar phenomena have been observed in the treatment of some diseases. Although antioxidants have been found to have several beneficial effects on neurodegeneration (26), clinical evidence is still relatively unsatisfactory (16). Datotop, an antioxidant therapy for Parkinson's disease (PD) supplemented with isoprene and tocopherol, was proposed by the National Institutes of Health in 1987 to slow down early PD. After 10 years of clinical trials, however, this treatment has had no obvious effects on PD (43).
In addition, some clinical trials and biological studies have indicated a controversial role of antioxidants in carcinogenesis, some studies showing that antioxidants can effectively kill liver cancer cells (67, 102), whereas others, such as two independent studies by researchers from Sweden and the United States published back-to-back in Cell in 2019, have reported a mechanism by which antioxidants can promote lung cancer metastasis (5, 64, 106). In nonsmall-cell lung cancer, nuclear factor erythroid 2-related factor 2 (NRF2) (or nuclear factor erythroid-derived 2-like 2 [NFE2L2]), an important transcriptional factor initiating the antioxidant system, is hyperactivated, either through stabilizing mutations in NFE2L2 or by inactivating its negative regulator Kelch-like ECH-associated protein 1 (KEAP1), leading to BTB domain and CNC homolog 1 (BACH1) stabilization to stimulate cancer metastasis. This breakthrough suggests that the original redox state of cells is a key point in determining the effect of antioxidants. Moreover, the type of tumor should be considered when considering antioxidant treatment; for example, supplementation of β-carotene, vitamin C, or vitamin E can prevent colorectal cancer but not gastrointestinal cancer (14).
Why do antioxidants fail to work well in aging or in some diseases closely related to oxidative stress? These problems suggest that we need to recognize redox more accurately. Sies et al. stated in their review of “oxidative stress” that there is no overall cellular redox state (89); global redox measurement and intervention have to be reconsidered. In this review, we propose “precision redox” and discuss and analyze the nature of redox from the following three aspects: differences in status, differences in function, and differences in therapeutic effects.
Redox Status
Redox status varies with different redox indicators
Redox regulation systems in cells include small molecules and macromolecules. Reactive species are diverse in chemical nature and include reactive oxygen species (ROS), reactive nitrogen species (RNS), reactive chlorine/bromine species (RCl/BrS), reactive sulfur species (RSS), reactive carbonyl species (RCS), and reactive selenium species (RSeS) (89). The major ROS/RNS species include hydrogen peroxide (H2O2), hydroxyl radicals (·OH), superoxide radicals (O2 −), nitric oxide (NO), and peroxynitrite (ONOO−). Antioxidant enzymes, including superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase, glutathione S transferase (GST), glutaredoxin, NO synthase, and S-nitrosoglutathione reductase (GSNOR), are the main macromolecular redox regulation systems. These enzymes are important for regulating ROS/RNS levels. Moreover, some redox couples, including glutathione/glutathione disulfide (GSH/GSSG), nicotinamide adenine dinucleotide hydrogen (NADH)/nicotinamide adenine dinucleotide (NAD)+, nicotinamide adenine dinucleotide phosphate hydrogen (NADPH)/nicotinamide adenine dinucleotide phosphate (NADP)+, Cys/Cyss, peroxiredoxin (Prx or Prdx)-sulfiredoxin, and thioredoxin (Trx)/Trx disulfide, work in concert with antioxidant enzymes to regulate cell redox status (20, 36, 48). These redox species or proteins involved in redox regulation can be used to indicate the redox status of cells.
The redox status of cells is classically assessed by measuring the redox potential (Eh), expressed in millivolts (mV). Different redox thiol couples differ in their redox potential; for example, in the cytoplasm, EhGSH/GSSG is approximately −220 to −260 mV, EhNADPH/NADP+ is approximately −241 to −393 mV, and EhCys/CySS is approximately −70 to −160 mV (20, 34, 75). In addition to differences in redox potentials of each parameter at resting state, there may also be differences in the changes in some parameters in response to stimulation. For example, in HeLa cells, H2O2 treatment significantly increases the fluorescence value of Nuc-HyPerRed (nuclear H2O2-sensitive probe) but has no effect on the ratio of NADH/NAD+ in the nucleus (116). At the tissue level, different species also reflect different redox status. For example, in tumors, redox status varies in different parts of the tumor. The level of antioxidant enzymes in nonmalignant tissues adjacent to vulvar squamous cell carcinoma is significantly lower than that in the malignant tissues (95). And, GSH level is higher in nonmalignant tissues near gastric tumors, but there is no significant difference in antioxidant enzyme levels (95). Therefore, the redox status of cells varies with redox species.
Redox status is spatiotemporally regulated
The redox status in different organelles is different. For example, the redox potential (Eh) of the GSH/GSSG couple is about −300 mV in mitochondria, −220 to −260 mV in the cytoplasm, −130 to −153 mV in the cell membrane (20, 32, 34), −150 mV in the endoplasmic reticulum (ER) (22), and −240 mV in lysosome (10). This shows that the mitochondria, cytoplasm, and lysosomes are in a relatively reduced state, whereas the ER and cell membrane are in a relatively oxidized state. Trx redox potential can also be measured and is used to assess the redox status in different organelles. The redox potential (Eh) of the Trx1(−SH)2/SS couple is about −300 mV in the nucleus, −210to −280 mV in the cytoplasm, and that of the Trx2(−SH)2/SS couple is about −360 mV in the mitochondria, suggesting that the nucleus is more reductive than the cytoplasm and more oxidative than mitochondria. Using these indicators, we can evaluate the redox status of the major organelles in the cell (Fig. 1) and can see that cellular redox is dependent on subcellular compartmentalization.
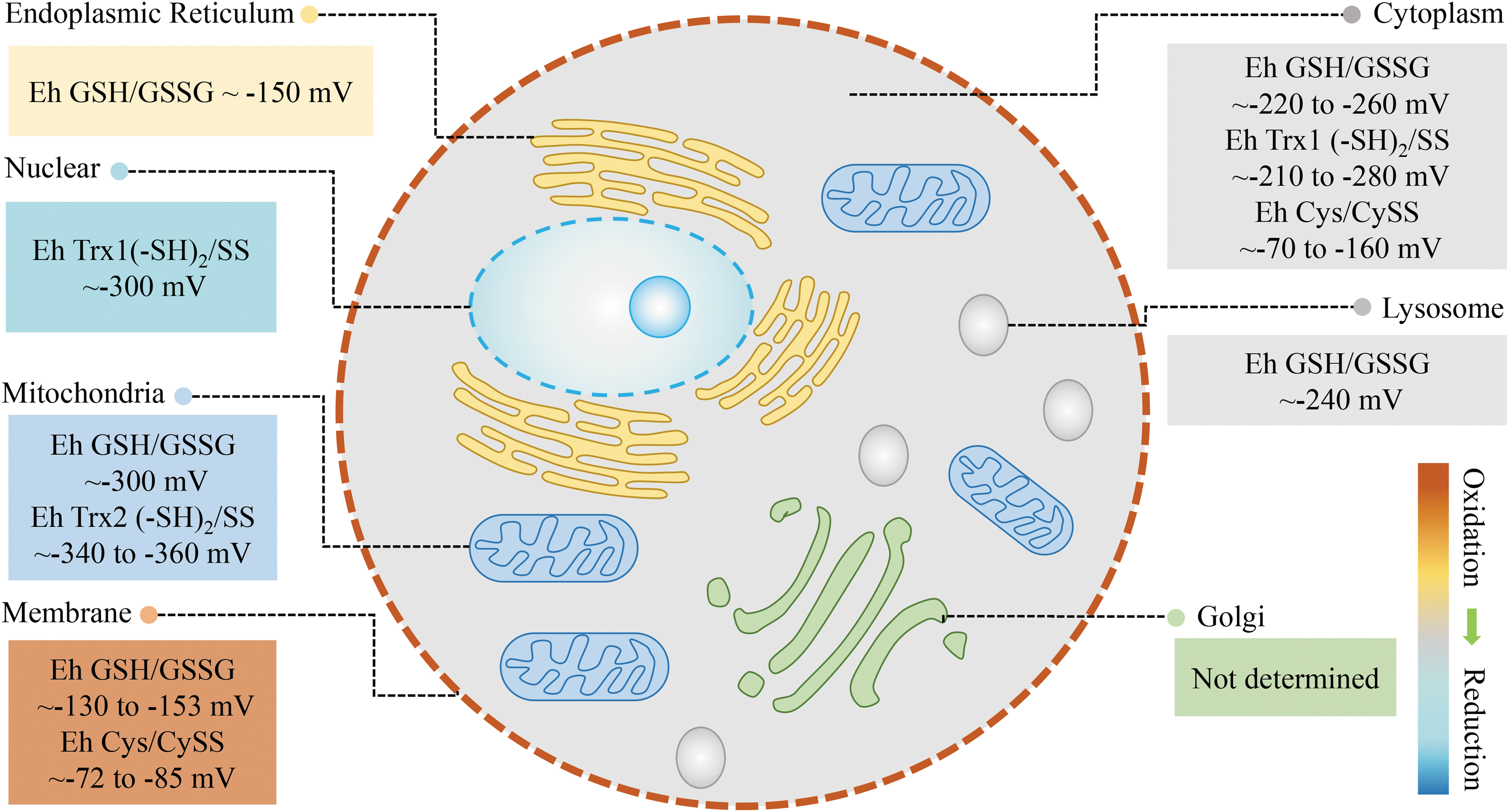
The cell cycle is a very important process in the life and activity of the cell. Some studies have detected changes in redox indicators at various stages of the cell cycle using genetically coded fluorescent probes. The ratio of NADPH/NADP+ increases significantly at cell division and returns to its original level after division. This suggests that redox state is different at different points of the cell cycle, showing periodic changes. However, there is no significant change in other indicators (NADH/NAD+, thiol, H2O2) during the cell cycle, providing further evidence that different redox species reflect different redox status (116). In addition to cell cycle-dependent regulation, redox status is also regulated rhythmically. It is reported that endogenous H2O2 levels oscillate rhythmically in cells and in the mouse liver throughout circadian cycles. This suggests that there is a relatively oxidized state during the active phase of the mouse circadian cycle and a relatively reduced state during the resting phase (77). In Parastacus brasiliensis promatensis, the SOD activity in the gonads of females increases in the summer when compared with the autumn; conversely, CAT and GST levels do not change throughout the year. Tissue-specific grading of lipid peroxidation intensity has also been observed in females during the spring: gonads < muscle < gills < hepatopancreas (79). These reports indicate that the redox status of different tissues is also variable and may change with season.
Changes in redox status at different stages of life are also spatiotemporal. During the aging process in Caenorhabditis elegans, the redox status in the cytosol and mitochondria becomes more oxidative with age; however, the redox status in the ER shifts from oxidative toward reductive with age (56). This not only shows that the redox state is constantly changing throughout the life cycle but also that redox changes are different in different organelles. We have also shown that the expression of GSNOR increases in the aging brain of humans and mice, indicating that the level of NO also changes in these tissues with aging (113). Interestingly, some studies have found an increase in the total oxidant status (TOS) and total antioxidant capacity (TAC) during labor and delivery in humans (65) and livestock (18). At the same time, the oxidative stress index (OSI = oxidation level/antioxidant level) is unchanged despite the increases in TOS and TAC. The OSI decreases and NOx levels increase significantly 1 week postpartum (17, 94), indicating that redox status and oxidative stress during late pregnancy and the postpartum period are different.
Redox status also changes under different external environments or pathological conditions. Redox state is reported to change in different tissues of hibernating animals during different periods of hibernation (105), different tissues varying in their level of oxidative stress. H2O2 content in the heart and brain during late torpor (LT) is elevated compared with levels during the summer active (SA) state, implying that these tissues are more susceptible to the effects of oxidative stress. In the process of hibernation, oxidative stress and antioxidant indices increase or decrease but then return to SA levels after hibernation, indicating that the redox state changes with the process of hibernation, and it has significant controllability. In alloxan-induced hyperglycemic rabbits, the level of nitrotyrosine (NT) is elevated in the pancreas, kidney, and heart, whereas the testicular level of NT is unaffected. Some changes in antioxidant enzyme levels also differ in these tissues, indicating that tissues from hyperglycemic animals differ in their ability to neutralize nitrosative stress (33).
Redox status is thus regulated precisely in a spatiotemporal manner, and different locations (tissues or cellular organelles) and time periods, including different stages of the cell cycle, circadian rhythms, seasons, different stages of life, external environments, and pathological conditions, should be borne in mind to provide an accurate evaluation.
Redox status may vary in different individuals
In addition to differences in the redox status of different cells or tissues, at the systems level, there are also differences in redox status between individuals. First, there are differences in redox levels between species (59, 92, 93). One study has found that the activity of SOD and the concentration of CAT and GSH in the liver, heart, and brain of mice, rats, guinea pigs, rabbits, pigs, and cattle are different. Taking SOD activity as an example, in the liver, the order of species from high to low SOD activity is: pig, guinea pig, cattle, rabbit, rat, and mouse, whereas for the heart, the order is: pig, mouse, cattle, rabbit, rat, and guinea pig. The order for the concentration of GSH in the liver (from high to low), however, is: mouse, rat, guinea pig, rabbit, pig, and cattle (92). Another study also found that the rates of superoxide anion and H2O2 production were different in five species of dipteran flies (93). A typical example of individual differences in redox is the naked mole-rat (NMR, Heterocephalus glaber), the rodent species with the longest known life span (>28.3 years). The level of GSH/GSSG in mole-rats is much lower than that in similar-sized mice. Moreover, the NMR shows 10-fold higher levels of in vivo lipid peroxidation than mice, indicating that maybe it is not only oxidative stress that modulates longevity (6, 62, 84). These results not only indicate the difference in redox levels between species but also provide evidence that different redox indexes reflect different redox levels in different tissues.
In addition to differences between species, even in mice, there is also variation in redox states between different strains in response to external stimulation. Caloric restriction (CR) prevents age-related decrease in GSH:GSSG in C57BL/6 mice, whose life span is extended by CR, but not in DBA/2 mice, in which life span remains unaffected (83). At the individual level, clinical studies show that the level of antioxidant enzymes varies greatly between individuals with the same cancer type (12). The application of antioxidants in disease treatment therefore should also consider these individual differences in determining the best approach.
Different Functions Correspond to Different Redox Statuses
Subcellular redox status is closely related to cell function
As mentioned above, the spatial distribution of redox species and thiol couples in cells is not uniform (20, 32). This difference is closely related to cell function. The mitochondrial electron transport chain is the main origin of ATP synthesis and one of the main sources of ROS production; mitochondrial redox state is thus involved in the regulation of cell metabolism. A higher level of NADH/NAD+ is necessary in the mitochondrial matrix for the reductive force of mitochondrial ATP production (49). The redox thiol couple NADPH/NADP+ also regulates metabolism by modulating tricarboxylic acid (TCA) cycle-associated enzymes. Mitochondria need to maintain a more reduced state to have the highest rates of electron transfer, and oxidative stress will lead to the damage of electron transport chain (34). Nuclear redox state regulates cell proliferation and the cell cycle. A relatively reduced state provides the conditions for optimizing enzymatic activity that is required for transcription factors to bind DNA in the nucleus (61, 111). The redox potential of the cytosolic NADH/NAD+ and NADPH/NADP+ is more oxidized than in mitochondria (42, 49, 107). The physiological concentration of H2O2 in the cytosol is maintained in the submicromolar range (0.01–100 nM) (40). At this concentration, H2O2 mainly functions as a signal molecule through redox relay of highly reactive proteins. Therefore, a slightly more oxidized state in the cytoplasm than in the mitochondria is beneficial for the rapid and accurate transmission of redox signals. A much more oxidized state is necessary in the ER for oxidative protein folding (7).
In addition to redox small molecules, antioxidant enzymes and other redox regulatory macromolecules also differ in their distribution or level in each organelle in line with different functions. Trxs are ubiquitous antioxidant enzymes that play important roles in many health-related cellular processes. Mammalian cells possess two Trx isoforms, Trx1 and Trx2. Trx1 is localized in the cytosol and translocates to the nucleus during oxidative stress. Cytosolic and nuclear Trx1 are regulated independent of each other and exhibit different functions (34, 104). However, Trx2 is present in mitochondria and functions in mitochondrial redox homeostasis (34, 104). Prxs are also ubiquitous, highly expressed antioxidant enzymes forming up to 1% of cellular protein content (19). Prx exists in six isoforms (PrxI to PrxVI) in mammalian cells, Prx I and Prx II being localized in the cytosol and nucleus, Prx III in mitochondria, Prx IV in the ER and Golgi and extracellularly, Prx V in the cytosol, mitochondria, and peroxisomes, and Prx VI in the cytosol (29). These different Prx subtypes are not redundant, and differences in their function are gradually being reported in more and more studies (1, 29, 55). In summary, the various redox state parameters in each subcellular compartment can regulate specific target proteins that contribute to different cell functions. In other words, from the perspective of cell function, precision redox is very important.
The levels of ROS/RNS determine if they play an advantageous or detrimental role
In cells, ROS are involved in redox signaling or oxidative stress, depending on their concentrations (Fig. 2). Low levels of ROS activate signaling pathways to initiate biological processes, whereas high levels of ROS lead to oxidative damage to DNA, protein, and lipids, and even to cell death (53, 86). ROS also play dual roles in cellular inflammatory responses, dependent on concentration. A low elevated level of ROS in the immune system might enhance normal immune function, whereas high levels of ROS generation lead to elevated levels of proinflammatory cytokines, causing cell and tissue damage (39). In stem cells, moderate ROS levels are required for proper stem cell differentiation and renewal through activation of signaling pathways; however, much higher ROS levels lead to stem cell exhaustion and premature aging (86). In tumor cells, accumulating evidence suggests that treatment with low concentrations of ROS promotes the growth and metastasis of tumor cells. Only high concentrations of ROS, exceeding the damage threshold, can repress tumor cell growth (30, 70). In addition, different levels of ROS or ROS in different organelles have opposing effects on life span at the individual level. Increased superoxide levels arising from the addition of low levels of paraquat extend the life span of wild-type worms, whereas high doses decrease life span (100). As for different organelles, elevated ROS in mitochondria act to increase life span, whereas elevated ROS in the cytoplasm decrease life span (85). These two results indicate that mild elevation of mitochondrial superoxide levels can increase life span, whereas high levels of superoxide are toxic. This clearly shows that the effect of superoxide on life span is dependent on where and how much superoxide is present.
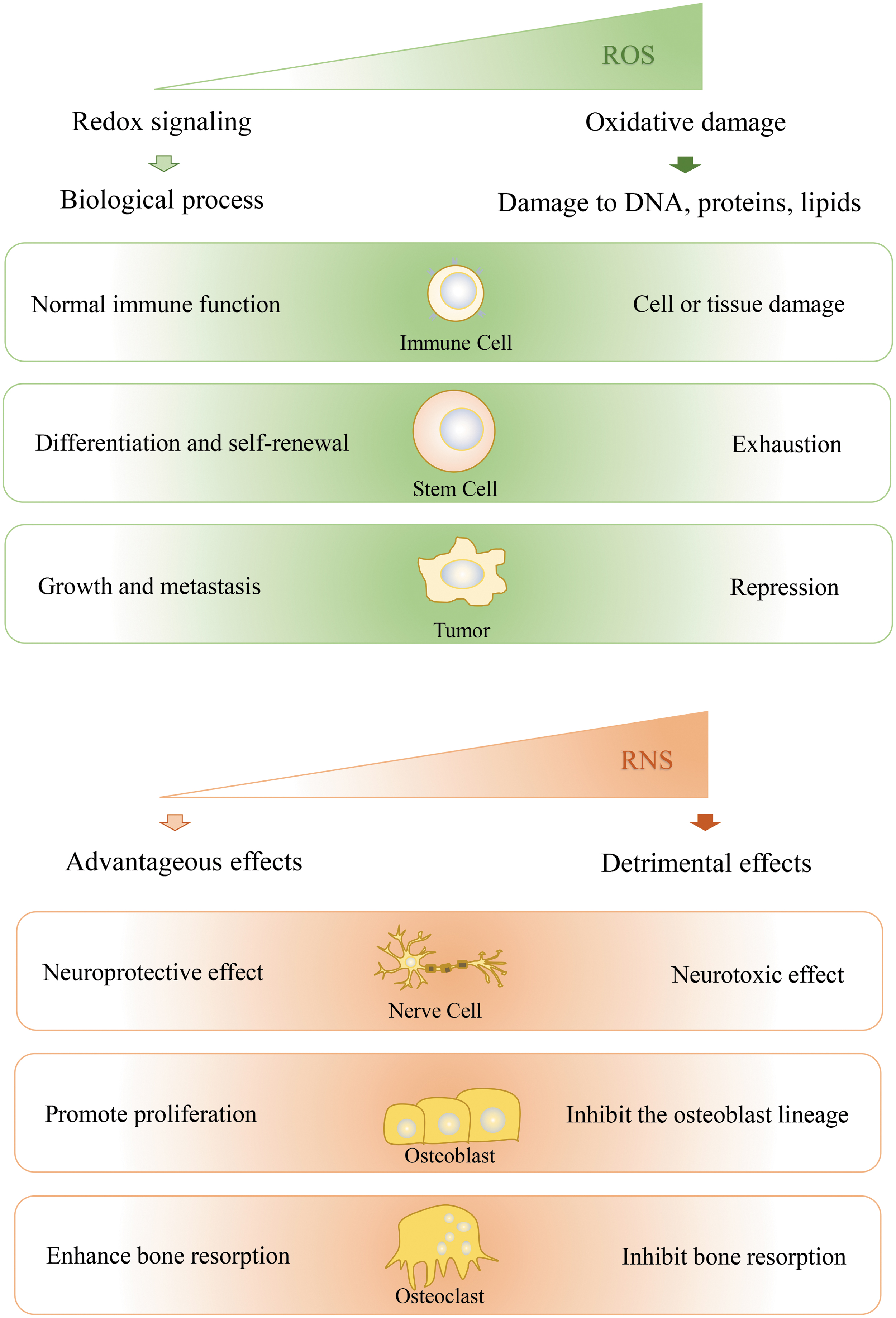
Similarly, the function of NO can also be advantageous or detrimental, depending on its concentration (Fig. 2). For example, in the nervous system, physiological levels of NO are neuroprotective, whereas higher concentrations are neurotoxic (15). NO has also emerged as an important biological mediator of bone cell function. At lower concentrations, NO promotes the proliferation of osteoblast cells, whereas at high concentrations, NO is inhibitory for cells of the osteoblast lineage (25). In addition, NO also has a biphasic effect on osteoclast activity and bone resorption: low concentrations enhance bone resorption while high concentrations inhibit bone resorption (45). Therefore, the level of a given redox molecule should be measured accurately, so as to evaluate its function accurately.
Although the impact of ROS/RNS in the cell varies with its level, the effect of ROS/RNS is not proportional to its concentration. There seems to be a concentration threshold that determines the transition from advantageous to detrimental effects. This concentration threshold, however, is not absolute and may be different in different cells, at different times or under different conditions. Generally, the physiological concentration of H2O2 in the cytosol, which functions as a signal, is in the submicromolar range (0.01–100 nM). Adaptive stress responses occur at higher concentrations from about 50 nM to 1 μM. Concentrations of H2O2 greater than 1 μM may cause growth arrest or cell death (88). When the cell is in a specific physiological process, such as the oxidative burst that is crucial to fertilization, ROS levels will also change. In addition, the same level of ROS may play different roles in different types of cells or in different individuals. For example, tumor and nontumor cells have different ROS susceptibilities. Normal cells have generated less ROS and have effective antioxidant systems. When ROS are further increased via therapeutic approaches, levels of ROS in tumor cells reach the death threshold earlier and the tumor cells are more easily killed. Therefore, to evaluate the concentration threshold correctly, multiple factors such as place, time, and condition should be considered comprehensively.
ROS/RNS function differs with different targets
ROS/RNS function as signaling molecules by covalently modifying specific cysteine residues in redox-sensitive target proteins. However, they have different effects on the modification of different target proteins. ROS oxidize and activate apoptosis signal-regulating kinase 1 (ASK1) to activate its downstream JNK pathway (91) but inhibit the activity of phosphatase and tensin homolog deleted on chromosome 10 (PTEN) by oxidative modification (73). The oxidative modification of Keap1 causes its conformational change and dissociation from NRF2 (111). NO can activate the autonomous activity of CaMKIIα by S-nitros(yl)ation, and GSNOR downregulates CaMKIIα signaling activity by decreasing S-nitros(yl)ation (113). NO inhibits the enzyme activity of ATG4B by nitros(yl)ation (63) and inhibits the aminoacylation and editing activities of mitochondrial threonyl-tRNA synthetase by S-nitros(yl)ation (115). In addition, NO controls the nuclear export of the DNA base excision repair protein adenine/pyrimidine endonuclease 1 (APE1) through S-nitros(yl)ation (81). The nitros(yl)ation of SUMO E3 ligase Pias3 promotes its degradation via the ubiquitin pathway, indicating that redox modification can affect other post-translational modifications (82). NO inhibits the interaction of nuclear export receptor CRM1 and NES by nitros(yl)ation, leading to the inhibition of the classical nuclear export pathway (103).
Oxidative modification of different members of the same family of proteins may even have different effects. For example, it is found that NO reduces NADPH oxidase 5 (Nox5) activity by reversible S-nitros(yl)ation. However, the inhibition of NO on the activity of Nox 1–5 isoforms is different, Nox1 being the most sensitive Nox isoform, and Nox4 the most resistant to the inhibitory actions of NO (80). Different extents of oxidative modification of the same protein may also result in different functions. For example, if Prxs are oxidized to form dimers, they will transduce ROS signals to target proteins, such as regulatory kinases or transcription factors. Hyperoxidation of Prx (Prx-SO2), however, may lead to the dynamic formation of multimeric complexes that function as chaperones to protect cells from protein aggregation (23). In conclusion, redox modification of different target proteins has different functions, including the regulation of protein activity, conformation, and location, and exerts an influence on other post-translational modifications and on protein–protein interactions.
In addition to their signaling role, excessive ROS/RNS lead to oxidative damage. The targets of oxidative damage include proteins, nucleic acids, and lipids. When the ROS level is too high, cysteine sulfhydryls within a protein can be directly oxidized to sulfonic acid (-SO2H) or sulfonic acid (-SO3H). These oxidations are generally irreversible and cause oxidative damage to cells and tissues (27). Tyrosine and tryptophan are also major ROS/RNS targets. Carbonylation of proteins, for example, directly affects the folding and degradation of proteins (99). Lipids are more susceptible to oxidative modification than proteins. The final products of lipid peroxidation include malondialdehyde and 4-hydroxy-2-nonenal. Oxidatively damaged phospholipids can cause cell death. Guanine is most susceptible of the DNA bases to oxidative damage; 8-OHdG, a product of DNA base oxidation, can be used as a marker for damage caused by oxidative stress (99). ROS/RNS, therefore, exert different functions on different targets by oxidative modification. It is thus necessary to be able to distinguish different targets to understand or regulate the redox status of cells.
Redox stress has different effects at different disease stages
Oxidative stress is considered to be closely related to many diseases including diabetes, neurodegenerative diseases, and cancers. Diabetes is caused by insulin resistance and insulin secretion impairment, and oxidative stress is thought to contribute to the development of diabetes. In fact, in the early stage of diabetes, reductive stress occurs due to high blood glucose levels. High blood glucose results in excessive production of acetyl-CoA that enters into the TCA cycle, producing excess NADH (98, 109). At the same time, the polyol pathway and overactivation of PARP [poly (ADP-ribose) polymerase]-mediated decrease in NAD+ levels also contribute to an increase in the ratio of NADH/NAD+. Oxidation of the excessive NADH then leads to the production of more ROS in the mitochondria, leading to the expansion of oxidative stress. Reductive stress followed by oxidative stress could serve as the major mechanism underlying diabetes (108, 109).
Alzheimer's disease (AD) is also closely related to redox. ApoE 4/4 and apoE 3/4 gene carriers are at high-risk of AD; they are in a state of reductive stress state before they develop AD (11). At the beginning of AD, there is chronic oxidative stress and the antioxidant system is activated, the GSH/GSSG level increasing significantly. The antioxidant system is unable to resist the oxidative stress, which eventually culminates in further oxidative stress, protein aggregation, mitochondrial damage, and cognitive damage (11, 66, 69). This suggests that reductive stress can be used as an indicator of pathogenesis in the early stages of AD and that redox states are different at different stages of AD.
Cancer is another disease closely related to redox. The biological functions of ROS appear to be dependent on tumor stage. At early precancerous and neoplastic stages, antioxidant activity decreases and the accumulation of intracellular ROS leads to oxidative DNA damage and mutations in prooncogenes and tumor-suppressor genes, promoting cancer development. In late stages of cancer progression, the activity of antioxidant enzymes and the intracellular pool of NADPH and GSH increase to buffer the excessive production of ROS and limit oxidative damage (8, 114). Thus, antioxidants are likely to help tumor cells escape apoptosis in the late stages of cancer. Antioxidant use should be prohibited in patients with advanced stages of cancer and/or undergoing radiotherapy.
Different Effects of Redox-Based Therapy
Failed antioxidant therapy clinical trials
As oxidative stress injury is closely related to aging and many diseases, antioxidant agents have been used therapeutically for these conditions, but their efficacy is often not satisfactory (Table 1). Although many redox-based drugs have achieved good results in cell experiments, they have failed in animal experiments or clinical trials. Oxidative stress is a key feature of the atherothrombotic process involved in the etiology of heart attacks, ischemic strokes, and peripheral arterial disease. It stands to reason that antioxidants represent a credible therapeutic option to prevent cardiovascular disease progression, but many studies and clinical trials have failed to show benefit (37, 38, 46, 58). For example, a clinical study recruited 2545 women and 6996 men, aged 55 years or older, who were at high risk for cardiovascular events. The results showed that there was no significant effect of vitamin E administration on cardiovascular events in these patients for an average of 4.5 years (46). Another high-risk study recruited 20,536 British adults with coronary heart disease, other occlusive artery disease, or diabetes who were randomly assigned to receive an antioxidant vitamin supplement (vitamin E, vitamin C, and carotene) or a placebo. While this therapy significantly improved the concentration of vitamins in the blood, it did not significantly reduce the incidence rate of any type of vascular disease (37). A trial in healthy men also showed that 12 years of supplementation with carotene was not beneficial to cardiovascular diseases (38).
Some Antioxidant Therapies that Failed in Clinical Trials
AD, Alzheimer's disease; CVD, cardiovascular disease; PD, Parkinson's disease.
PD, a common degenerative disease of the central nervous system, is also closely related to oxidative stress. However, as mentioned in the Introduction section, after 10 years of clinical trials, the antioxidant therapy Datotop was shown to have no significant effect on PD (43). Some clinical semi-quantitative food-frequency questionnaire studies have also reached similar conclusions; vitamin E and vitamin C intake from food and supplements is not related to the risk of PD (41, 112). In another neurodegenerative disease, AD, curcumin plays a role through antioxidant action. However, clinical trials have failed due to its low solubility, rapid metabolism, and clearance in vivo (4). In another clinical study, 769 subjects with amnesia subtypes of mild cognitive impairment were randomly assigned to receive 2000 IU of vitamin E daily for three consecutive years. Compared with the placebo group, there was no significant difference in the possibility of developing AD in the vitamin E group; that is to say, vitamin E was not beneficial to patients with mild cognitive impairment (78).
With respect to the use of antioxidants in cancer therapy, in addition to the fact that antioxidants (β-carotene, vitamin C, or vitamin E) have no inhibitory effect on gastrointestinal cancer (14), as mentioned earlier, a large number of clinical intervention studies have reported negative results. In the ATBC trial (the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study), 29,133 male smokers were randomly assigned to receive β-carotene, α-tocopherol, or both, daily for 5–8 years. Results showed that supplementation with α-tocopherol or β-carotene did not prevent lung cancer, and β-carotene even significantly increased the risk of lung cancer (2). In an epidemiological study based at a Japanese public health center in which 24,226 women aged 40–69 years answered a baseline questionnaire, no statistically significant correlation was found between plasma tea polyphenol levels and breast cancer risk (44). In another clinical study, 126 women at risk of breast cancer were treated with mixed soy isoflavones or placebo for 6 months, but the intervention did not reduce breast epithelial proliferation (54). Thus, it is worth reflecting on the failure of these antioxidants as clinical therapies; the general use of antioxidants does not appear to work as expected.
Examples of successful antioxidant therapies
What is the reason behind failed antioxidant strategies? Examples of successful antioxidant therapies provide some clues (Table 2). As cardiac and vascular tissues are highly dependent on mitochondrial homeostasis, several antioxidant therapies have targeted the mitochondria to treat cardiovascular diseases. Some small mitochondrially targeted peptides that can scavenge H2O2 have been used to inhibit lipid peroxidation in cardiac hypertension in animals (96). Hemigramicidin-TEMPO conjugates, novel mitochondria-targeted antioxidants, have been reported to prolong survival time in a rat model of hemorrhagic shock (28). Antioxidants that are targeted to the mitochondria thus seem to have a good therapeutic effect in cardiovascular diseases in animal models, emphasizing the importance of targeting the right place. The therapeutic effect of such novel mitochondria-targeted antioxidants on cardiovascular diseases thus deserves further clinical evaluation.
Some Examples of Successful Antioxidant Therapies
AAV, adeno-associated virus AMD, age-related macular degeneration; GEE, monoethyl-ester of GSH; GSH, glutathione; MnPs, Mn(III) N-alkylpyridylporphyrins; NAC, N-acetylcysteine; SOD, superoxide dismutase.
It is reported that PD is associated with a loss of total GSH that may contribute to progressive cell death. General antioxidants that do not specifically increase GSH levels may thus not have a very good therapeutic effect. Peripheral GSH administration has been shown to be effective in clinical trials (87), indicating the importance of the right redox species. The monoethyl-ester of GSH (GEE) has been reported to produce a dose-dependent elevation in GSH in rat mesencephalic cultures, protecting neurons from oxidative stress (110). However, in vivo, only central delivery of GEE resulted in significant elevations in brain GSH. In contrast, peripheral administration of GEE subcutaneously delivered by an osmotic pump did not result in the elevation of GSH levels in any of the brain regions examined (110). The manner of administration is thus important to ensure that there is a high enough dose of drug in the effective part, and it is not affected by metabolism or clearance.
With respect to tumor treatment, treatment strategies that target different redox species have been widely tested and applied. For example, Imexon, an agent that targets GSH, is in clinical trials for use in lung cancer (Phase I) and pancreatic cancer (Phase II) (24), and Tetrathiomolybdate, an agent that blocks cytosolic SOD1 and can inhibit melanoma, myeloma, prostate cancer, and breast cancer in Phase II clinical trials (50). In addition to different species, redox therapies that target specific organelles have also attracted attention in recent years. For example, nonmitochondria-targeted antioxidants N-acetylcysteine (NAC) and soluble vitamin E analog, Trolox, are reported to prevent tumorigenesis in a mouse hepatocarcinogenesis model, whereas mitochondria-targeted antioxidants SS-31 and Mito-Q (a derivative of ubiquinone) promote tumorigenesis (102).
In addition to targeting specific locations and using the right species, antioxidant treatments given at different time points also have different effects. In an experiment on antioxidant therapy for age-related macular degeneration, decreases in the electroretinography response and thinning retinal thickness were significantly delayed on early treatment with the Sod 2 vector but were not affected with the treatment when the mice were 6 months old (13). Similarly, in diabetes, Mn(III) N-alkylpyridylporphyrins (MnPs), analogs of SOD, have opposite effects at different stages in streptozotocin-induced diabetic mice. MnPs injected at the onset of diabetes inhibit oxidative damage and prolong life, whereas MnPs injected at the eighth day after diabetes onset aggravate oxidative damage (3). This means that antioxidant gene editing therapy may be effective in preventing the detrimental effects of oxidative stress but may not be beneficial once substantial tissue damage has occurred. In conclusion, different doses of antioxidants may show different therapeutic effects in different locations and at different times for different targets. To be successful, redox-based therapies must apply the right species in the right place, at the right level and at the right time.
Antioxidants in personalized nutrition and exercise
Antioxidant supplements have been discussed in the exercise nutrition field for several decades. At first, antioxidants were thought to be effective against the oxidative stress injury caused by exercise. Later, many views suggested that the use of antioxidants would inhibit exercise-induced adaptability. In 2018, some new ideas were put forward. NAC supplementation was found to enhance exercise performance by increasing GSH concentration and by reducing oxidative stress only in individuals with low resting levels of GSH (76). The differential effects of antioxidant supplementation on physical performance (VO2max), oxidative stress (F2-isoprostanes), and antioxidant concentrations were shown to depend on the baseline antioxidant concentration (68). That is to say, there are individual differences in baseline redox status and the effectiveness of antioxidant supplements to decrease oxidative stress and promote health depends on the baseline redox status. These authors proposed a novel strategy named the stratification of individuals based on their antioxidant profile. An “ideal” analytical tool could assess an individual's systemic antioxidant profile via analyzing a capillary blood sample, for example, design the most optimal nutritional redox treatment. Accurate recognition and evaluation of individual differences in redox status is thus also an important aspect requiring attention in antioxidant application.
In addition to being used as exercise supplements, antioxidants such as vitamin C are widely used as dietary supplements. So are antioxidant-rich foods good for everyone? A new study published in Nature on mutations in the p53 gene and intestinal cancer suggests that we should think twice about our use of antioxidants, bearing in mind individual circumstances (51). Mutant p53 fulfils different functions in different parts of the intestinal tract: in the colon, mutant p53 has an obvious carcinogenic effect, whereas in the small intestine, it has an obvious tumor-suppressive effect. The reason for this difference in function is that gut microbes in the colon produce a metabolite, the antioxidant gallic acid. This antioxidant is found in high levels in foods such as black tea, chocolate, nuts, and berries. When p53-mutant mice were fed with gallic acid, a malignant phenotype was detected throughout the gut (51). This study suggests that antioxidant-rich foods should be consumed with caution in people at high risk of colorectal cancer. The use of antioxidants should thus take into account not only individual differences in baseline redox status but also individual differences in other health indicators. The daily use of antioxidants as nutritional supplements thus also requires precision and should vary from person to person.
Summary
We have introduced the precise nature of redox from three aspects: differences in redox status, differences in function, and differences in therapeutic effects. It is obvious from the literature that redox status must be considered in the context of species, time, place, level, and target. The function of a biomacromolecule and its role in cellular signaling are closely dependent on redox status. Accurate evaluation of redox status and specific interventions are critical for the success of redox treatments. Precision redox is the key for antioxidant pharmacology.
We have thus proposed a “5R” principle of precision redox: “Right species, Right place, Right time, Right level, and Right target” (Fig. 3). Right species refers to the need to focus on specific redox species, including reactive species (ROS/RNS/RClS/RBrS/RSS/RCS/RSeS), antioxidant enzymes, and redox couples. Right time emphasizes the need to consider the dynamic changes in redox status at different times, such as the different stages of the cell cycle, circadian rhythms, seasons, aging, or diseases processes. Right place means that different organelles, different cell types, and different tissues should all be considered. Right level refers to the need to pay attention to the effective range of its effect when researching or applying redox, otherwise if the oxidation level is too high, it will lead to oxidative stress damage, or if the reduction level is too high, it will lead to reduction stress. Right target has two meanings: one is to regulate the oxidation level of specific substrates (such as proteins, lipids, and nucleic acids) at the micromolecular level, and the other is to establish different antioxidant application methods for different individuals at the macroindividual level.
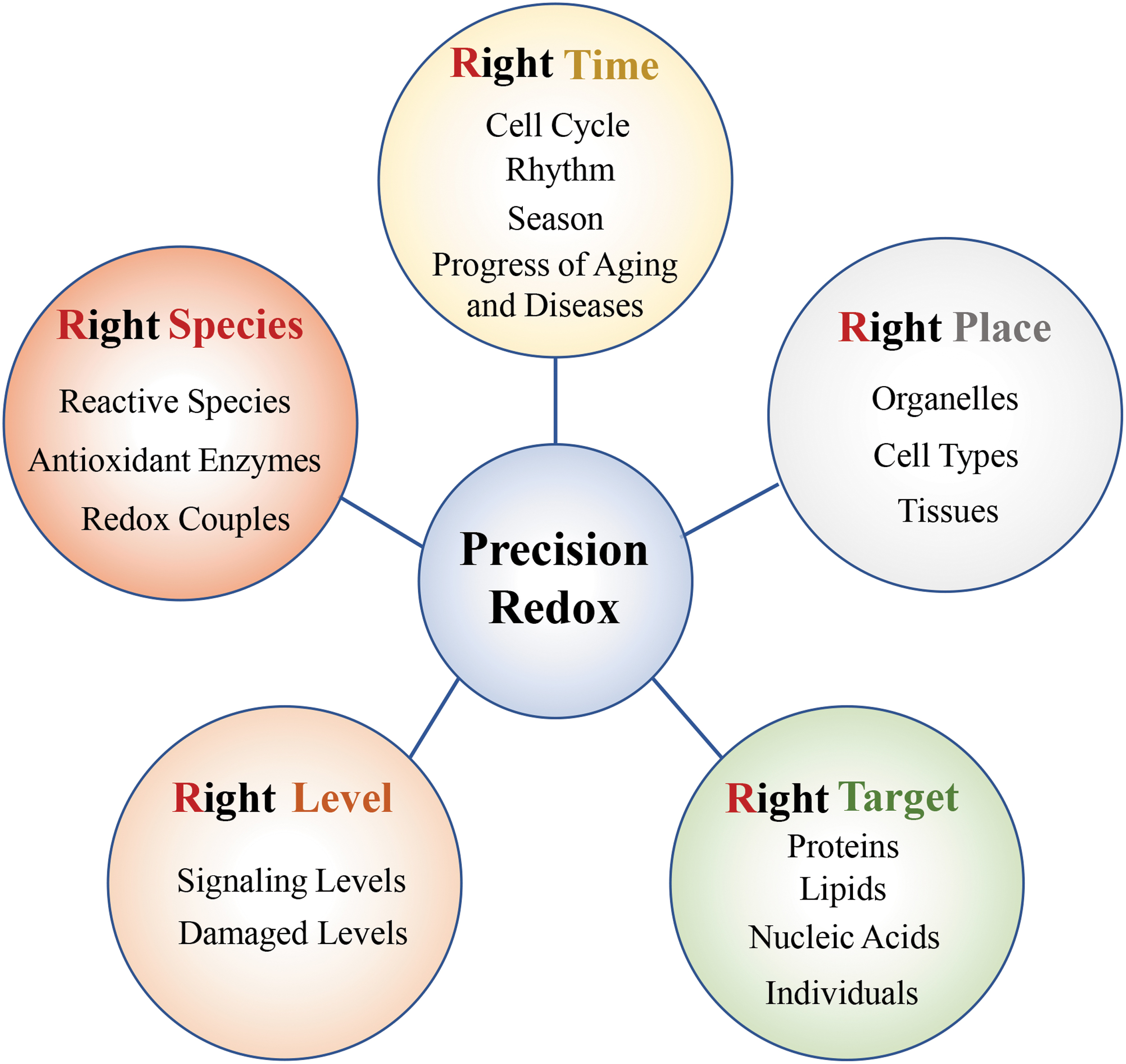
Previously, some scientists have mentioned the concept of redox compartmentalization and pointed out that the redox status is distinct between organelles and have suggested that accurately measuring subcellular redox will be facilitate the accurate regulation of redox in disease prevention and treatment (31, 47, 52). It is suggested that redox analysis should be specific rather than global, and it is necessary to quantify key redox molecules and pay attention to their spatiotemporal regulation (89, 90). With respect to redox-dependent tumor treatment, some scientists have also proposed that it is necessary to study redox couples with spatial, temporal, and chemical specificity, to precisely design redox drugs for specific stages in tumor growth, vitality, and migration (21). These points of view are consistent with our emphasis on the right species, right time, and right place. Since different concentrations of ROS/RNS play completely different functions, the importance of redox level has been recognized. In addition, the accumulation of protein oxidative modifications or S-nitros(yl)ation modifications has been found to be closely related to various diseases. Scientists have begun to advocate to improve or treat diseases by mutating oxidative modification sites of specific proteins. For example, specific mutations in the S-nitros(yl)ation site of IRE1α have been shown to improve glucose homeostasis in obese mice. This suggests the importance of specific targets in redox-based therapy. All these findings support our “5R” precision redox principle.
In view of the complexity of redox, the “5R” precision redox principles should be considered in antioxidant pharmacology rather than applying global nonspecific antioxidant treatments. As these five factors affect each other, it will be necessary to consider all or several factors at the same time to achieve real precision redox-based therapy and allow antioxidants to really play their therapeutic effects. With the development of redox detection technology, the application of this precision redox principle will help us to understand more accurately the overall picture of cell redox and should greatly improve the success rate of redox-based therapies.
Footnotes
Authors' Contributions
C.C. and J.M. conceived and wrote the review. Z.L. helped to search literature and draw figures. The other authors helped to search literature. All the authors read and approved the article.
Acknowledgment
We thank Dr. Joy Fleming for English editing of the article.
Funding Information
This study was supported by the National Key R&D Program of China (Grant No. 2017YFA0504000), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB39000000), and the National Natural Science Foundation of China (Grant Nos. 91849203 and 31900893).