
Abstract
Introduction
I
Early research clarified a fundamental issue in the assessment of kinetic versus thermodynamic properties in the analysis of redox processes in biology. Specifically, the steady states of deviation from thermodynamic equilibrium provide a source of order (85). The specific location of an enzymatic activity in subcellular structures or in distinct organs, that is, a kinetic potential, is used to influence the reduction–oxidation state of a given redox couple, defined by its thermodynamic redox potential as expressed by the Nernst equation. This enables multiple chemical and associated physical processes to occur simultaneously within the physical space, obeying the same thermodynamic forces, having distinct spatial and temporal response characteristics, and supporting differentiated functions. The two major systems addressed here, the nicotinamide nucleotide (NAD and NADP) systems and thiol systems (protein thiols, glutathione [GSH], and associated disulfides), substantially differ in organization and exemplify the central operational structures of redox biology.
The redox code of these metabolic and protein components provides a critical complement to the genetic code, the epigenetic code (80), and the histone code in the molecular logic of life. Unlike the genetic code and histone code, which underlie information storage and transfer for use, the epigenetic code and the redox code define the operations of the genetic and histone codes in the organizational structure, differentiation, and adaptation of an organism to the environment. Like the genetic and histone codes, the redox code has a fundamental simplicity, yet it is elaborated with considerable complexity in multicellular organisms. In particular, redox signaling and redox control in multicellular organisms evolved and diversified with the increase in atmospheric O2 about 600 million years ago (54, 93). The present review highlights key principles of the redox code derived from the extensive literature on redox sensing and signaling in diverse processes of circadian variation, cell cycle, apoptosis, and organismic defense (51). The term “Redox Code” applies to the redox organization of cells, tissues, and organisms. It is not confined to mammalian systems, and it extends ultimately to all living matter. While focusing mainly on eukaryotic mammalian systems in this review, it is understood that other eukaryotic, for example, plant and fungal systems, as well as prokaryotic, for example, bacterial, systems have elaborated redox codes describing their respective metabolic organization.
Principles of the Redox Code
The redox code consists of four principles by which biological systems are organized (Fig. 1). The first is the use of the reversible electron accepting and donating properties of nicotinamide in NAD and NADP to provide organization of metabolism, operating at near equilibrium. Substrate oxidations are linked to reduction of NAD+ and NADP+, which in turn are linked to ATP production, catabolism, and anabolism, respectively. The second principle is that metabolism is linked to protein structure through kinetically controlled redox switches in the proteome, which determine tertiary structure, macromolecular interactions and trafficking, activity, and function. The abundance of proteins and reactivity of sulfur switches with oxidants vary over several orders of magnitude to determine specificity in biological processes. The third principle is that of redox sensing, in that activation/deactivation cycles of redox metabolism, especially involving H2O2, support spatiotemporal sequencing in differentiation and life cycles of cells and organisms. The fourth principle is that redox networks form an adaptive system to respond to the environment from microcompartments through subcellular systems to the levels of cell and tissue organization. This adaptive redox network structure is required to maintain health in a changing environment and, if functionally impaired, contributes to disease and organism failure.
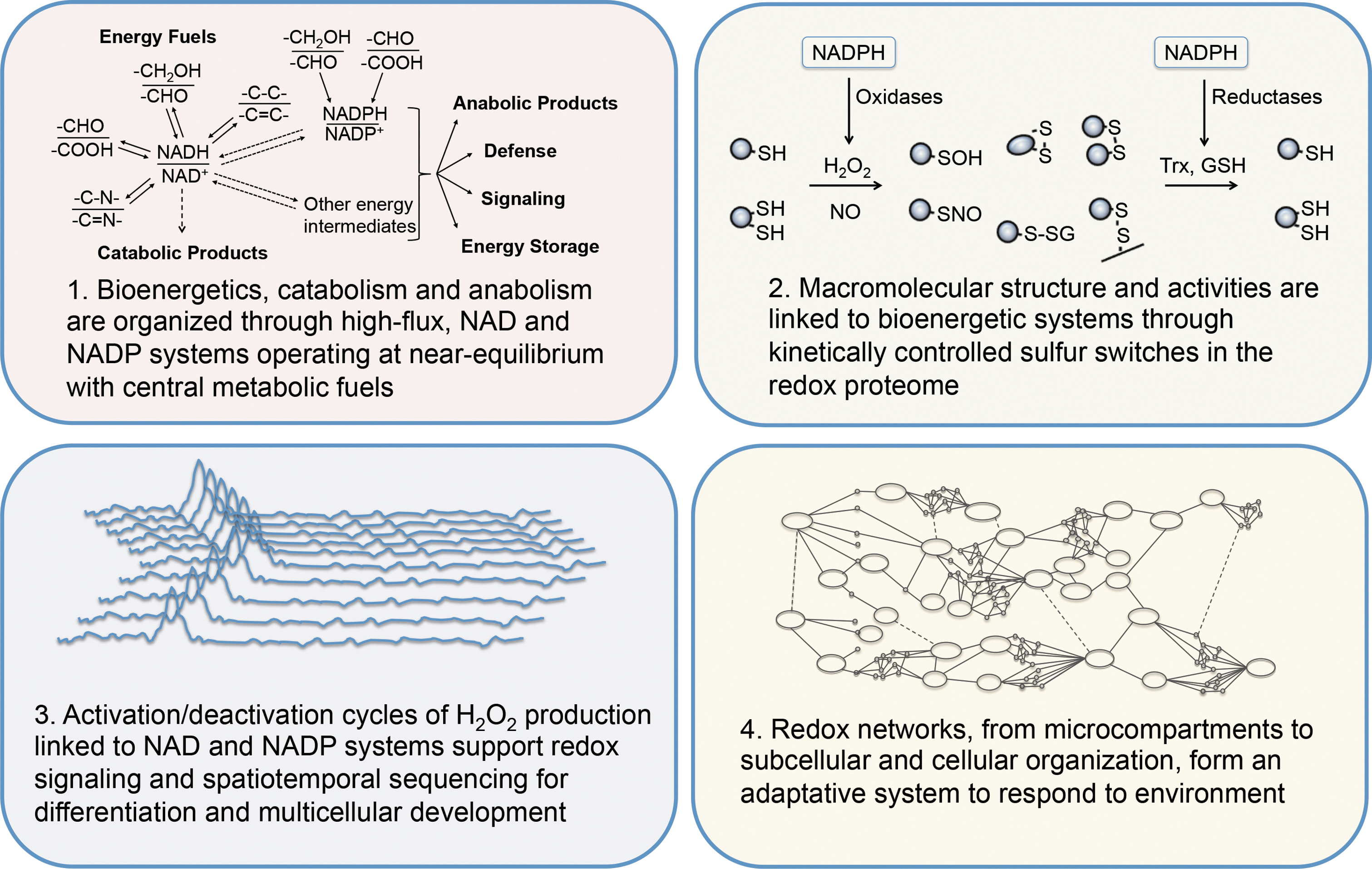
Redox Players
Redox biology occurs in a multidimensional space, in which the fundamental basis set, that is, the full spectrum of redox elements and dimensionality of this space, remains incompletely understood. In developing the redox code, we focused on a limited set of combinations to outline central principles for bioenergetics, metabolic and structural organization, and interaction of an organism with its environment. As indicated below, it does not incorporate the full spectrum of redundant and unique uses of specific redox systems [see also Buettner et al. (17)]. Consequently, the redox code should be viewed as a central framework rather than specific vectors leading to specific redox states and associated biological states as observed in the life cycles of diverse life forms. Future elaboration will be needed to fully encompass the selenoproteins, hemoproteins, cuproproteins, iron–sulfur proteins, flavoproteins, pteroproteins, molybdoproteins, manganoproteins, and other redox elements and interacting systems.
The NAD and NADP systems along with thiol/disulfide systems are central to the redox code. These are outlined in Table 1 and addressed in more detail below. The NAD system uses powerful dehydrogenases, active in reversible two-electron transfer catalysis, in the metabolism of biomolecules with covalent bonds of C, O, S, N, and H. [NADH]/[NAD+], the ratio of concentrations of free NADH and NAD+, is in equilibrium with the reduced and oxidized substrates of major cellular dehydrogenases (11). The [NADH]/[NAD+] couple is central to catabolism and energy supply. The [NADPH]/[NADP+] system is set apart from the [NADH]/[NAD+] and is central to anabolism, defense, and control of thiol/disulfide systems by controlling the redox state, for example, in the GSH and the thioredoxin (Trx) systems. The [NADPH]/[NADP+] and [NADH]/[NAD+] systems are linked by activity of mitochondrial energy-linked transhydrogenase (40, 127). One-electron transfer is biologically less relevant in the NAD and NADP systems, but important in associated components, such as FAD and FMN systems, the flavin-linked reactions (30), and in iron–sulfur clusters (7). Redox chemistry of thiols involves both one-electron and two-electron transfer reactions of cysteine (Cys)/cystine (CySS), GSH/glutathione disulfide (GSSG), protein thiols and oxidized forms, and protein-bound cysteines/disulfides. The selenium analog of cysteine, selenocysteine, also plays an important role in controlling the redox function of proteins [see Jacob et al. (47)]. The role of selenium in selenoprotein biology [see Refs. (44, 84)] and the chemical properties of selenocysteine versus cysteine (76, 94) have been studied intensively. Protein thiols are subject to other modifications (e.g., nitrosylation, sulfhydration, metal ion binding) impacting redox activities (51). These and other redox players provide an enriched diversity to the central redox organizational structure described here.
For discussion, see text.
There are exceptions, for example, the lipoamide in pyruvate dehydrogenase is high flux, likewise ribonucleotide reductase in an S-phase T cell.
HIF-1α, hypoxia-inducible factor-1α; NAD, NADP, nicotinamide adenine dinucleotide; NF-κB, nuclear factor-κB; Nrf2, nuclear factor-E2-related factor 2.
First Principle: Metabolic Organization
Metabolism is organized through high-flux thermodynamically controlled NAD and NADP systems. The relevant property for redox systems is the activity of the redox partners, which is closely dependent upon their free concentrations within cellular and subcellular compartments. This concept is emphasized by the nonequivalence of tissue levels (nmol/g wet weight) with the free concentrations (nM) (101). Differences in free concentrations among compartments may result from permeability barriers, binding to macromolecules, nonhomogeneous distribution, and also heterogeneity of cell types in organs and tissues (Fig. 2A). Recent technological progress allows the resolution of redox partners within microcompartments (37) and such resolution can be expected to enable refined investigation in redox biology.
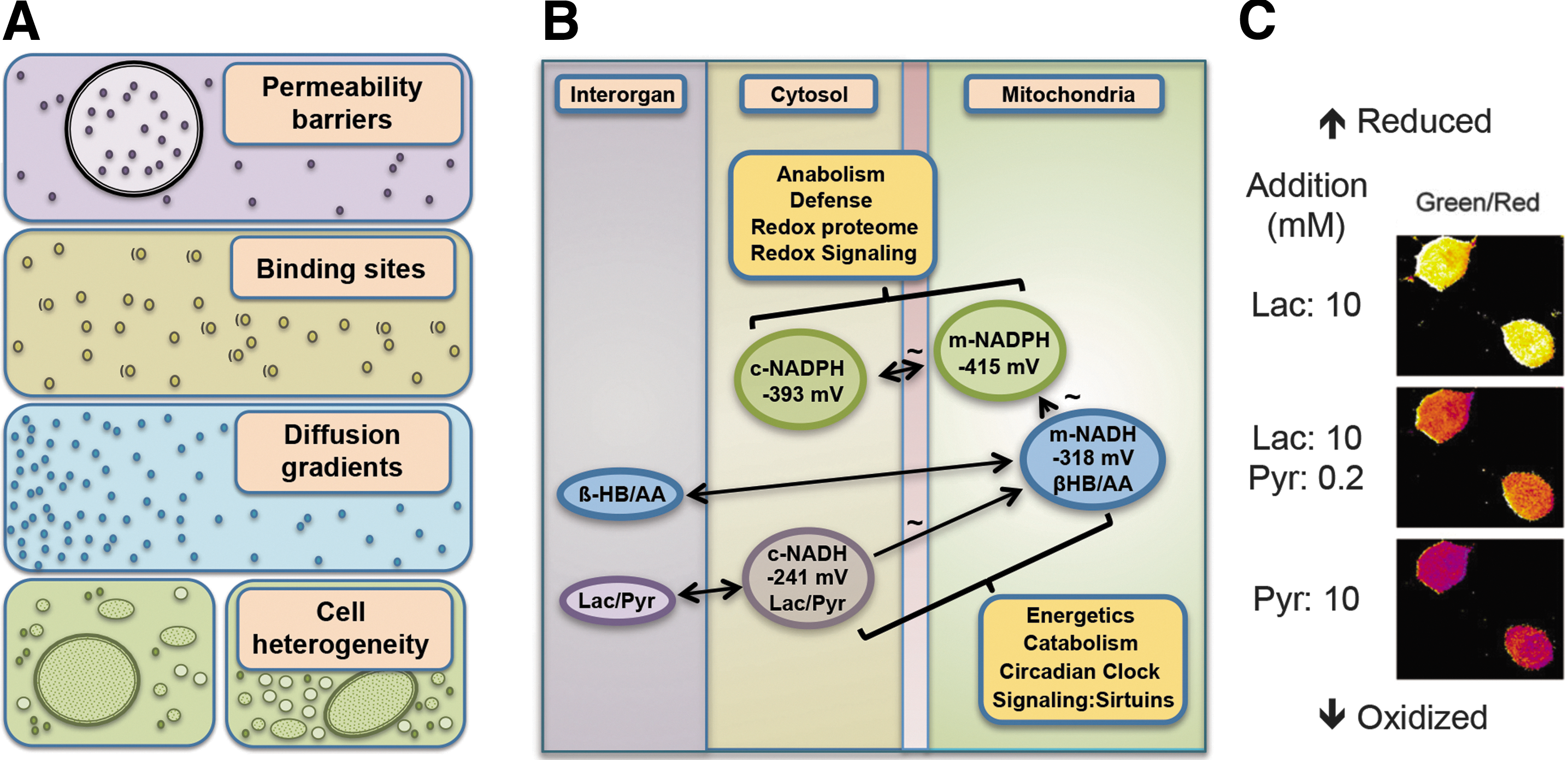
Detailed studies of the three major NAD-linked dehydrogenases showed that the systems operate near equilibrium (15, 42, 43). The cytosolic redox poise of [NADH]/[NAD+] is at a set point of −241 mV (Fig. 2B), substantially more oxidized than expected from the total cytosolic NADH and NAD+ content (14, 100). This difference is attributable to powerful protein-binding sites for NADH; the redox buffer capacity of these titratable NADH-binding sites provides a cytosolic set point at about 1 μM free NADH, despite much higher total cytosolic NADH (pNADH=6). Operation of the NAD system at near equilibrium allows cells to support catabolic needs while maintaining capability to supply ATP energy according to cellular needs. Studies of subcellular compartments showed that the redox potential of the cytosolic and the mitochondrial [NADH]/[NAD+] are vastly different by about two orders of magnitude or ca. 60 mV (15, 122). The mitochondrial matrix NAD operates at a more negative redox potential (86), −318 mV, providing a greater reductive force for mitochondrial ATP production. Noninvasive redox titrations have been extensively applied to study dynamic steady states using redox indicator metabolites in combination with organ spectrophotometry (14); newer methods use genetically encoded fluorescent biosensors (Fig. 2C) (46).
The NADP system provides an important counterpart to the NAD system, supporting anabolic reactions of metabolism, organismic defense, and maintenance of the thiol/disulfide systems. The [NADPH]/[NADP+] system is equilibrated with NADP-linked dehydrogenases and operates at a more negative redox potential than the NAD system, that is, −393 mV in the cytosol and −415 mV in the mitochondrial matrix (Fig. 2B) (100). As mentioned above, the mitochondrial NAD and NADP systems are connected by energy-linked transhydrogenase. This enzyme may act as a sensor that determines the positioning of either NAD or NADP at the near-equilibrium steady state [see Yin et al. (127)]. The reducing force provided by these potentials supports biosynthesis concurrently with NAD-dependent catabolism and energy production; the specificity of the NAD- or NADP-linked systems provides a central logic to these divergent needs in metabolism. This molecular logic is further illustrated by the use of the NADP system for defensive purposes, especially activation of O2 for otherwise unfavorable oxidation of xenobiotics and reduction of O2 for the generation of superoxide anion used in phagocytic killing of pathogens.
Second Principle: Linkage of Metabolism to Structure
The linkage of redox metabolism to structure involves the redox proteome. The redox proteome is a subset of the epiproteome, which includes phosphorylation, acylation, methylation, and other modifications of the translated proteome. Although functionally integrated with the systems controlling these other covalent modifications, the redox proteome provides a distinct basis to link metabolism to structure through kinetically controlled sulfur switches. These molecular redox switches determine tertiary structure, molecular activity, macromolecular interactions, and trafficking principally through changes of thiol and disulfide structures.
The nature of the thiol systems differs from the nicotinamide nucleotide systems, in that most elements operate at low flux rather than near equilibrium (Table 1). This characteristic is exemplified by steady-state redox potentials of extracellular low-molecular-weight thiol/disulfide couples, including Cys/CySS and GSH/GSSG pools (55). Using the Nernst equation, the calculated redox potentials in human plasma are −75 and −150 mV, respectively. While compartmentation and other factors (Fig. 2A) complicate interpretation of thiol/disulfide systems in cells and tissues, lack of equilibration in this cell-free aqueous space emphasizes the kinetic limitation. Reaction rates calculated from rate constants and concentrations for thiol/disulfide couples are too slow for most redox elements to equilibrate in living organisms. This has recently been emphasized in a discussion of the GSH redox system (8).
Cellular Cys/CySS and GSH/GSSG couples are maintained at a substantially more reduced state than the extracellular counterparts. Like the NAD and NADP systems, redox potentials differ in subcellular compartments, with the mitochondria more negative (reducing) and endoplasmic reticulum more positive (oxidizing) than cytosol. The thiol/disulfide set point for specific Cys residues in proteins is analogous to the NADH-binding sites mentioned above, with the thiol reactivity of individual protein Cys being tuned by location within the protein structure. Unlike the NADH sites, most of which have catalytic functions supporting energetics and metabolism, most protein thiols have noncatalytic functions, for example, protein folding, regulation of activity, control of macromolecular interactions, and directional trafficking (47). The low-molecular-weight thiols interact both enzymatically and nonenzymatically with proteins, presenting challenges to identify corresponding thiol/disulfide systems connected to them (26). Additionally, protein thiols undergo other chemical reactions and interact with metal ions, either of which can impact biological function.
Despite the extensive number of specific Cys (>200,000 encoded in the human genome) (50) and variation in reactivity (>6 orders of magnitude) and abundance (>6 orders of magnitude), progress is being made in understanding the redox organizational structure. Current evidence points to evolution of the Cys proteome (70) during the same period as evolution of multicellular organisms, advanced O2 sensing, and H2O2 signaling systems (54, 93). The Cys proteome increased with organismic complexity, from 0.5% of the proteome in prokaryotes to 2.2% in mammals (70). The expected Cys content in proteins based upon the random use of tRNAs is (3.1%), indicating that increase in Cys in the proteome occurred, despite selection pressure against use (51). This coevolution of the Cys proteome with the O2 sensing and H2O2 signaling in multicellular evolution is consistent with known structure–function relationships involving protein thiols in complex organisms.
Measurements of cells and tissues show that Cys occurs with different steady-state oxidation in proteins (33, 60). Use of thiol-trapping agents and mass spectrometry in the absence of added oxidants shows that median percentage oxidation of protein Cys is 6%–14%. These findings are consistent with measures of steady-state redox potentials of the central thiol antioxidants, Trx and GSH, which show that these couples are not in thermodynamic equilibrium with the reductant couple, NADPH/NADP+. Inhibitors of GSH and Trx antioxidant systems increase fractional oxidation (6), and extensive studies with nutrient deprivation, mitochondrial respiratory inhibitors, and oxidant challenges show that oxidation of the thiol proteome is dynamic and reversible. Thus, the redox proteome is characterized by widespread occurrence of protein Cys with partial oxidation.
A common operational structure governs the redox-sensitive Cys through metabolism dependent upon the NAD and NADP systems. In this structure, H2O2 is the most central oxidant, and Trx and GSH are the most central reductants (Fig. 3A). Measurements of steady-state concentration show that H2O2 in the mammalian liver is 1–10 nM (18, 82, 103, 105). Constitutive H2O2 is derived from mitochondrial respiration dependent upon NADH (10, 34, 87) and activities of NADPH oxidase (Nox) enzymes dependent upon NADPH (2, 13, 87). As a consequence, H2O2 production depends upon activity levels in cells. H2O2 production is countered by Trx and GSH/glutaredoxin (Grx) systems, both functioning to reduce disulfides, but with differing specificities for protein Cys (62, 65). Importantly, both Trx and GSH systems depend upon NADPH-linked reductases, creating a system in which protein oxidation and reduction are inherently balanced yet responsive to cellular activity through NADPH availability. This links energetics and metabolism to redox structure, which in turn uses oxidant signals through nuclear factor-E2-related factor 2 (Nrf2), peroxisome proliferator-activated receptor γ coactivator 1-α (PGC-1α), nuclear factor-κB (NF-κB), and other transcription factor systems to more broadly integrate energy efficiency, resistance of cells to injury, and cell repair (11, 23, 25, 28, 56, 61, 86, 92, 98, 118). The stability of this redox structure is further assured by the use of Trx and GSH (with GSH peroxidases) as reductants for hydroperoxides, thereby providing two levels of opposing reactions to assure stable steady-state operation of this redox network structure (Fig. 3A).
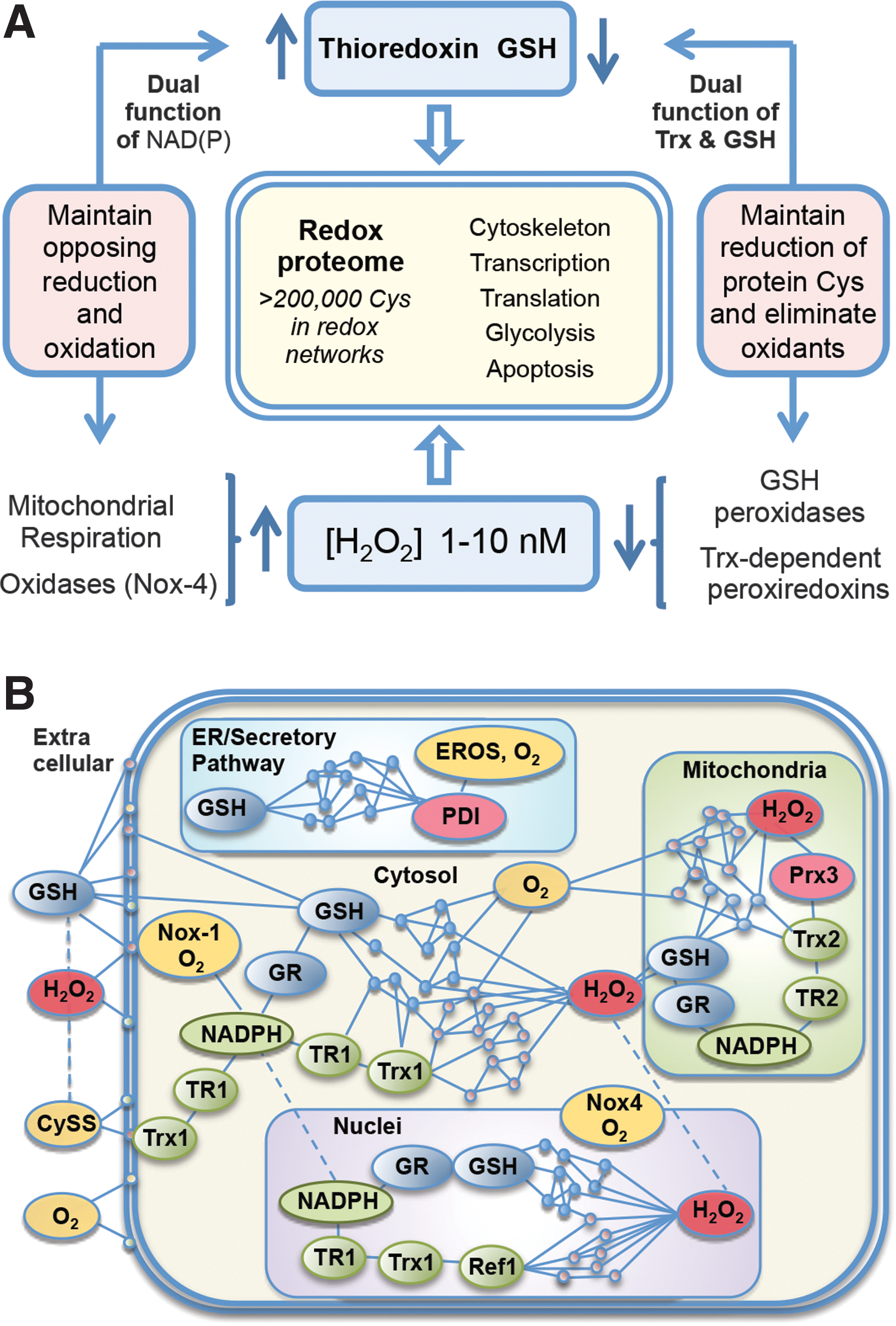
The fraction of the Cys proteome that undergoes reversible oxidation is unknown. Redox proteomics data provide evidence for thousands of redox-sensitive protein thiols (Fig. 3B), but a much lesser number have descriptions of associated function. Some have detailed understanding, such as those involved in transcription control via OxyR in prokaryotes (91), Nrf-2 (56), NF-κB (115), P53 (88), AP-1 (1), glucocorticoid receptor (78), and other transcriptional systems. In NF-κB, for instance, oxidation of Cys-38 in p65 or Cys-62 in p50 serves to control DNA binding. The redox pathway controlling reduction uses NADPH, thioredoxin reductase-1 (TR1), Trx1, and the nuclear redox factor-1 (Ref-1) to maintain the DNA-binding structure (115, 126). Nuclear peroxiredoxin-1 enhances activity, suggesting that endogenous H2O2 supports oxidative inactivation. Measurement of steady-state oxidation of each component in the pathway and sensitivity to changes in NADPH supply further show that the redox pathway is kinetically limited (50). Other examples of specific redox-controlled proteins occur in the respiratory chain NADH dehydrogenase (6), mitochondrial thioredoxin reductase-2 (TR2), and Trx2 control activation of apoptosis (113) and actin-associated proteins (33). The GSH systems provide alternative reductases that not only have some redundancy to Trx systems but also have some specific functions in parallel with Trx. By itself, GSH is a slow reductant. However, Grx uses GSH to catalyze reduction of disulfides. Grx1 in the cytosol/nucleus and Grx2 in mitochondria remove S-glutathionyl groups formed during cell signaling, regulation, or oxidative stress. Functional modules for translation, glycolysis, lipid metabolism, and mRNA and miRNA processing and trafficking Cys are associated within functional networks in subcellular compartments according to steady-state oxidation (Fig. 3B) (33). The central logic to the dynamic operational structure of these diverse systems lies within the stable redox proteome linked to NADH/NAD+ and NADPH/NADP+ metabolism.
Third Principle: Redox Signaling and Spatiotemporal Differentiation
The evolution of O2 sensing, H2O2 signaling, and the Cys proteome following appearance of an O2 atmosphere naturally suggests a relationship between these processes; the significance of such a relationship is crystallized by studies with modern redox probe technologies. Since the discovery of nonphagocytic Nox enzymes in mammalian systems, considerable research has focused on the role of H2O2 and Nox in cell signaling (35, 59). Redox probe studies now show that during Caenorhabditis elegans development, a temporal wave of H2O2 production occurs in the larval stage, followed by a decrease in H2O2 upon emergence of the adult (Fig. 4A) (58). Selective oxidation of protein thiols accompanies this H2O2 production (Fig. 4B). The data highlight the dynamic redox organizational structure that occurs within the temporal framework of multicellular development.
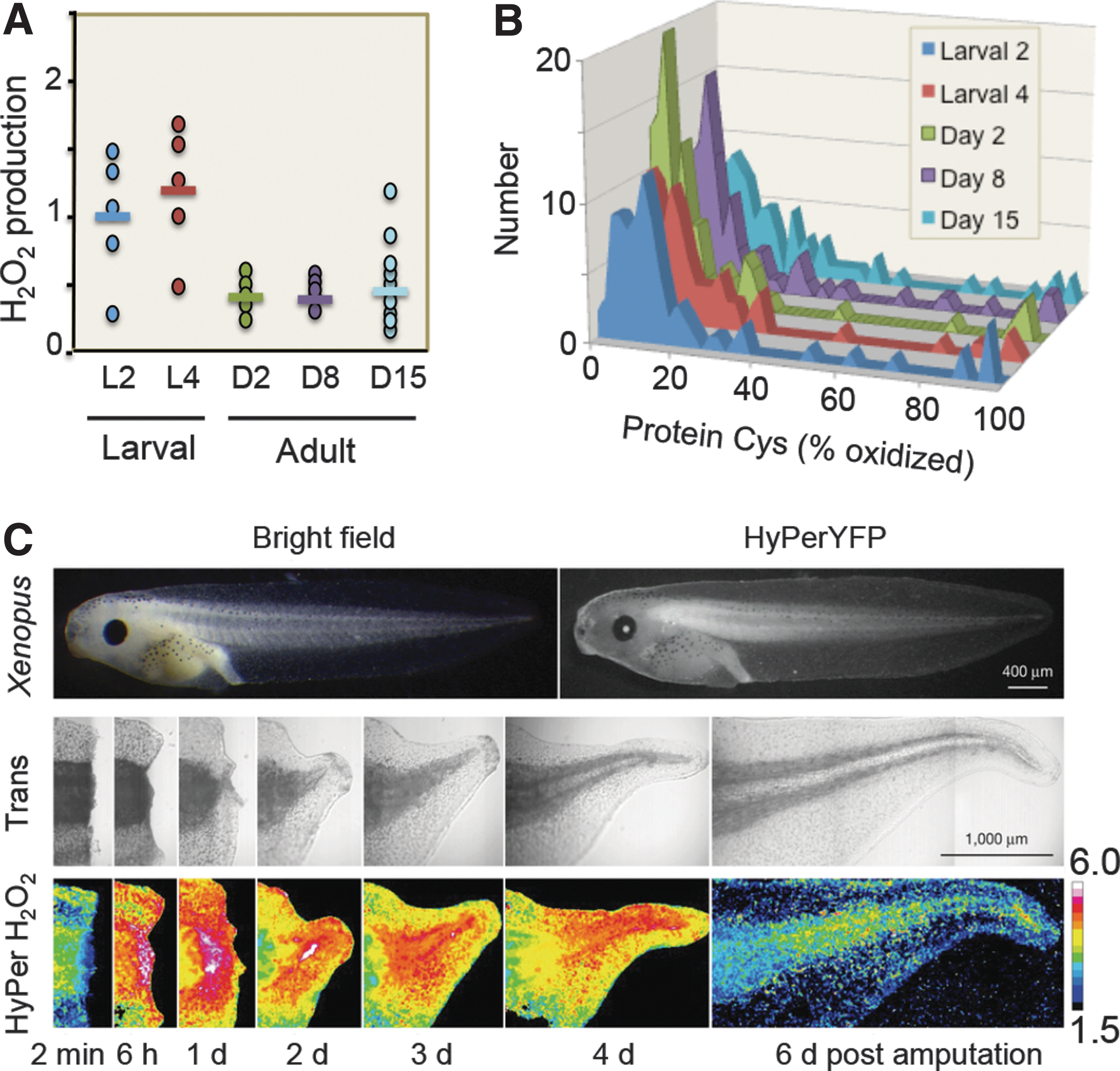
Redox probe studies of tadpole tail regeneration extend this to a spatiotemporal framework (Fig. 4C) (64). Assuming that the indicator responses measure H2O2 and not pH or other effects, the results show that enhanced production of H2O2 occurs at the wound site following tail amputation and progresses as a wave associated with regeneration. A tissue-scale gradient of hydrogen peroxide also mediates rapid wound detection in zebrafish (77). While molecular and pharmacologic manipulations have effectively demonstrated cellular redox mechanisms and roles in disease, the advanced bioimaging methods have enabled recognition of the third principle of the redox code, that is, activation/deactivation cycles of H2O2 metabolism support complex time-dependent processes in the life cycle of development of cells and organisms. Indeed, this echoes O2 sensing via hypoxia-inducible factor-1α (HIF-1α), a signaling system that requires O2 rather than H2O2 and provides signaling in multicellular organisms (99).
In spatiotemporal redox signaling, relatively high rates of local H2O2 production (2) provide an important contrast to the slow reaction rates that govern the bulk of the redox proteome described above (second principle). Localized H2O2 production may represent only a small fraction of total cellular O2 utilization, but results in locally high H2O2 concentrations (2). Diffusion of H2O2 and its rapid metabolism confines the redox signal to the vicinity of production. Specific proteins, such as peroxiredoxins, will react with hydroperoxides with substantial velocity. In principle, specificity could be obtained by the rate constants of redox elements for reaction with H2O2. An alternative is that peroxiredoxins or other proteins with high reactivity with H2O2 could serve as oxidative catalysts to provide specific oxidation of target proteins. This could occur in linear pathways or with a network reaction structure, with scaling of reactivities and abundance of nodes within the network. In such a network structure, one oxidative catalyst could have multiple protein targets, each with relatively high reactivity toward subsets of the cysteine or methionine proteome. Such a hierarchical structure could coordinate functional subsets of the proteome broadly over space and time.
Multiple other factors could also provide specificity. For instance, cationic and other proximal amino acids have a considerable effect on the reactivity of thiols with H2O2. pH is regionally controlled, especially in the vicinity of membranes, and thiol redox reactions are highly pH dependent. Metal ions are also important to stimulate (e.g., Cu2+) or inhibit (e.g., Zn2+) oxidation and these could impart specificity through protein–protein or other domain-specific structural interactions. The detailed mechanism and specificity of redox signaling by hydrogen peroxide needs to be further elaborated (57, 123).
Redox organization is well established in association with organellar function. For instance, in the endoplasmic reticulum, an endoplasmic reticulum oxidase system uses O2 and protein disulfide isomerase to introduce disulfides into proteins for the cell surface and secretion (116). This vectorial processing with successive oxidation results in relatively oxidized steady states of thiol/disulfide couples in extracellular compartments. Import into mitochondria uses similar redox processing, and multiple redox processes control transcription factor translocation into nuclei (32). One can extend these concepts to the cross talk between signaling mechanisms, for example, the widespread use of redox switches in kinase signaling pathways, such as oxidation of active site 215Cys in protein-tyrosine phosphatase 1B (PTP1B), underscores the spatiotemporal organizational structure. Subsets of protein Cys provide additional functional diversity through selective nitrosylation, sulfhydration, and binding to metal ions (51, 125) and irreversible modification of Cys by reaction with electrophilic metabolites (39). Reversible oxidation of Met to MetSO at the protein surface also occurs (29) and may provide a parallel redox network. Refined redox imaging tools will enable detailed mapping to enhance understanding of specialized redox functions.
O2 sensing allows maintaining NAD and NADP systems for energetics and metabolism, as defined above (first principle), while activation/deactivation cycles of H2O2 signaling direct targeted activities within a multicellular structure. Complementary NAD functions have recently been reviewed and highlight the importance of the subcellular location of NAD synthesis and the diverse roles of NAD in chromatin stability, DNA repair, circadian rhythms, and longevity (22). As an example, NAD+ activates sirtuins, which catalyze deacetylation of histones (68). Sirtuins are regulated by the [NADH]/[NAD+] ratio (27), and the [NADH]/[NAD+] ratio activates SIRT1-mediated autophagy (48). NAD plays a role in mitonuclear protein imbalance, which has been described as a conserved mechanism in the regulation of energy metabolism (45).
Thus, the molecular logic of differentiation and development in multicellular systems is fundamentally linked to three simple principles of redox biology defining the organization of energetics and metabolism, the linkage of metabolism to macromolecular structure and function, and the activation/deactivation cycles of H2O2 production that direct spatiotemporal organization. Although governed at all levels by the structural design of the NAD system to support energetics and metabolism, detailed knowledge of activation/deactivation cycles of H2O2 production and responsive redox protein systems will enhance the understanding of the differentiated functions and functional limitations in redox biology.
Fourth Principle: Adaptation to the Environment
The intricate relationships of NAD metabolism and circadian rhythms (74, 89, 96) underscore the central logic of redox systems in supporting organismic adaptation to environmental changes, including availability of nutrient resources, exposure to hazardous agents, and vulnerability to predators. This highlights the fourth principle of the redox code that redox networks, from microcompartments through subcellular systems to the levels of cell and tissue organization, form an adaptive system to respond to the environment. This adaptive redox network structure is required to maintain health in a changing environment and, if functionally impaired, contributes to disease and organism failure.
The fundamental linkage of diurnal changes to the light cycle is evident for photosynthetic organisms, but day–night cycles also determine the timing for food access in animals, and associated activities entrain susceptibility to environmental hazards and predation. Recently elucidated circadian redox clocks emphasize the conserved nature of diurnal variation of redox control in eukaryotes (Fig. 5A) (24). Coordination of NAD control of metabolism with NADPH/H2O2 control of spatiotemporal organization of the redox proteome supports key cellular activities, including mitosis and meiosis, differentiation, activation, senescence, and apoptosis, and complex multicellular processes, including fertilization, development, wound healing, and repair (95). The central positioning of H2O2 and thiol redox states in space and time is emphasized by the circadian variation in peroxiredoxins, abundant proteins that metabolize H2O2 and are central to redox signaling, and control of the thiol proteome (Fig. 5A).
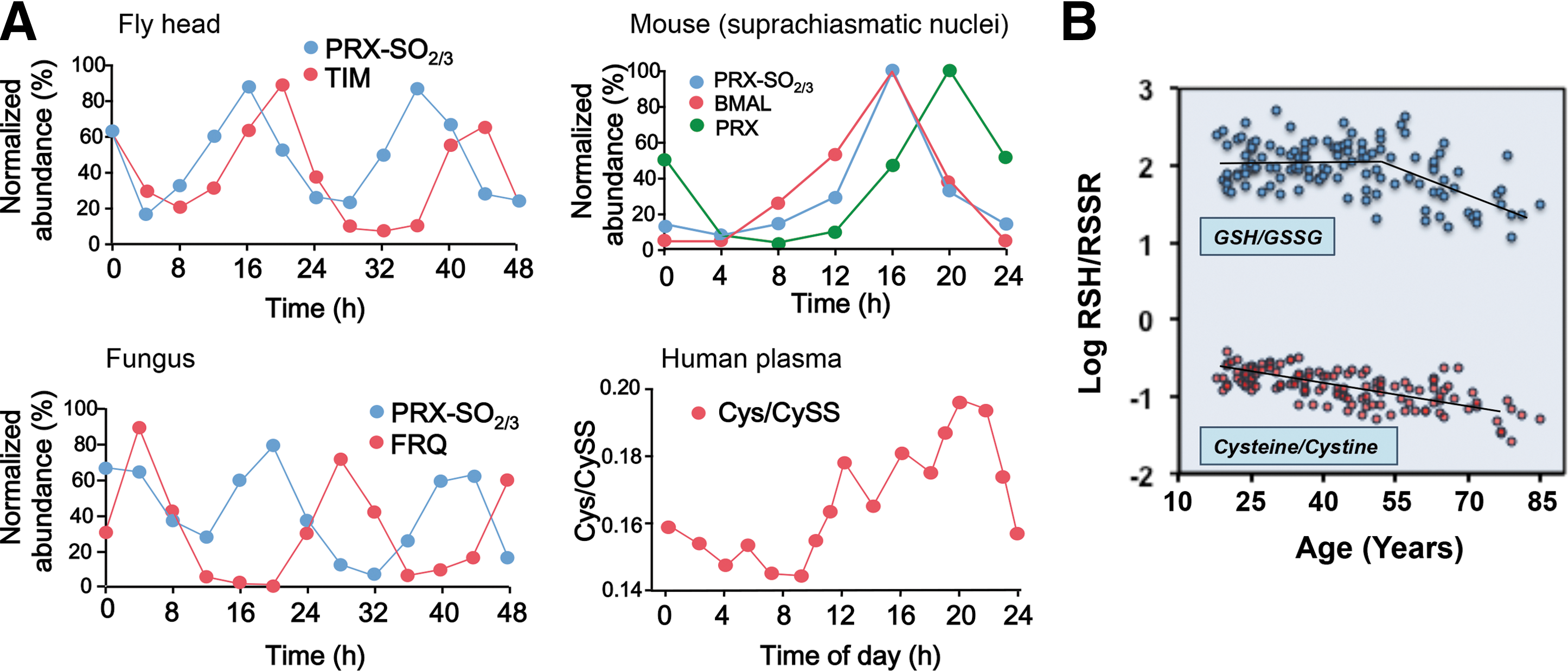
Mechanistic studies also provide knowledge of GSH/GSSG oxidation during the life cycle of mammalian cells (32), reversible glutathionylation of actin (69), mitochondrial NADH dehydrogenase (6) to integrate structure and metabolism, and genetic control to modulate GSH redox systems. A yeast model system studied with a fluorescence imaging probe (71) showed a substantially more negative GSH redox potential (very low free GSSG); the relevance for mammalian cell physiology will need to be addressed. Diurnal variation in the GSH system occurs in humans and is linked to the timing of food intake and the precursor amino acid, Cys (Fig. 5A).
Activity cycles are connected to nutritional needs and environmental threats, linking all specialized adaptive mechanisms ultimately to central homeostatic control of energetics, metabolism, and proteomic structure. Thus, redox integration is fundamental to the logic of these adaptive mechanisms, evident in the activity and response characteristics of the immune system, redox gradients within physical barriers, and adaptive responses, such as thermogenesis. GSH-dependent detoxification of reactive chemicals, protection from oxidants, and control of toxic metals, each depend upon this integrated redox network to tolerate and thrive in the presence of environmental challenges. NAD+ and the redox proteome networks are linked to life span with an incipient oxidation of steady-state thiol redox systems with age, providing a redox indicator for aging (Fig. 5B).
Together, the four principles of the redox code approach the central theory for the redox organizational structure of life. Along with the evolved genetic code to define an organism and epigenetic and histone codes to guide the use of the genetic code, the redox code supports organized implementation of the genetic code within energetic, metabolic, and environmental constraints of the organism. Knowledge of the network structure embodied in this code provides new avenues for redox medicine to understand and treat diseases.
Implications for Redox Medicine
Health and disease are distinguished, among other parameters, by maintenance or disruption of the network of redox steady states. The concept of oxidative stress describes deviation from physiological redox steady state toward oxidation [see Sies (104) for recent discussion]. As outlined above, the very functioning of cell metabolism is governed by redox processes. Fine-tuning of reaction cascades and/or direct involvement in signaling pathways involves molecular redox switches and participation of redox-active metabolites, such as H2O2, nitric oxide, or hydrogen sulfide. Considerable efforts have been deployed in redox research in biology and medicine. The following brief comments are meant to exemplify the current state and opportunities in this rapidly developing field. Since enzymes rather than low-molecular-weight micronutrients play the major role in redox biology (102), aspects of genetic polymorphisms of crucial redox enzymes and of transport systems are important (e.g., responders vs. nonresponders). While genetic polymorphisms are not treated here in detail, just one example might illustrate the point: the set point of the concentration of GSH, a low-molecular-weight component, has been described as a heritable trait (117).
An example relating nutrition to redox biology is given by the micronutrient, selenium, which is present as selenocysteine in selenoproteins, as mentioned above, and also as selenomethionine in place of methionine in some proteins. There is a diversity of selenium functions in health and disease (12), including epigenetic effects (107) and a role in adjuvant therapy of viral and bacterial infections (5, 108).
Aging
Irrespective of the question of cause and effect, an association of a more oxidized set point in the redox balance with increasing age has been amply documented. The fraction of cellular proteins present with oxidized side chains increases with age and is more prominent in diseases of premature aging (79). In blood plasma, the small-molecule thiols, cysteine and GSH, are increasingly present as disulfides as individuals are more aged (Fig. 5B) (9). Thus, protein oxidation in aging and the machinery of removal of oxidized proteins has attracted attention (41). Autophagy and its potential regulation by dietary micronutrients have been examined in terms of healthy aging (83). Apart from loss of proteostasis, the hallmarks of aging include genomic instability, telomere attrition, epigenetic alterations, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intercellular communication (63). Most, if not all, of these processes comprise a redox component (52). The NAD system is increasingly recognized as an attractive target for intervention in age-associated disorders (72). NAD precursors slow down aging and extend the life span in yeast and protect severed axons from degeneration in disease models.
Aspects of intermediary metabolism
Reactive oxygen species, H2O2 in particular, have been identified as signaling molecules in responses to extracellular stimuli such as insulin. Indeed, the insulin-dependent production of H2O2 is indispensable for propagation of intracellular signaling triggered by insulin (112). However, conversely, high levels of intracellular reactive oxygen species inhibit insulin activity and allow activation of stress responses, making for what has been called the peroxide dilemma (112). Interference with insulin signaling can also result from overexpression of hydroperoxide-reducing capacity, for example, by selenoproteins, and there has been an association between excessive selenium intake and incidence of diabetes type 2 (111), pointing to an interplay between selenium and carbohydrate metabolism (109). Likewise, strategies to improve the redox state in terms of obesity-associated oxidative stress have been examined (97). Diabetes type 2 has even been called a redox disease (120).
Cardiovascular arteriosclerosis
Oxidative damage from elevated production of reactive oxygen species contributes to ischemia–reperfusion injury in myocardial infarction and stroke. The enormous literature on this topic is not to be covered here [see Tocchetti et al. (114)]. One recent aspect might deserve special attention in the present context, linking protein redox modification to energy production: a specific cysteine switch on mitochondrial complex I has been identified by selective S-nitrosation of Cys39 on the ND3 unit of complex I (19).
Wound healing
Upon occurrence of a wound, the healing process begins as soon as damage arises. This process starts with conserved transcription-independent damage signals, such as H2O2 (Fig. 4C), calcium ions, and ATP, which initiate cell shape changes, functional actomyosin structures, and recruitment of immune cells (20). There are important redox signals in wound healing, and the recent identification of miRNA in wound inflammation and angiogenesis has led to the concept of OxymiRs, in particular miR-210 and miR-21, in their roles in responses to tissue oxygenation, skin development, and repair, as well as wound healing (91).
Immunology
Host defense is intimately linked to bacterial and virucidal oxidants generated by specialized cells, which are recruited and activated at the appropriate sites. Among the biological defense mechanisms, the production of superoxide by leukocytes was an early hallmark (3). Human fibroblasts release reactive oxygen species in response to interleukin-1 and tumor necrosis factor-α (67). The multiple aspects of redox processes in immunology are beyond the scope of the present article [see Nathan and Cunningham-Bussel (75)]. In inflammation, H2O2 is recognized as the messenger, guide, and assassin (124).
Proliferation of cancer
The relationship of redox processes with cell proliferation and apoptosis has been studied extensively and will not be presented here in detail. A recent overview on oxidative stress and cancer illustrates the range of the topic (106). The use of redox cycling drugs (53) has been a mainstay in cancer therapy.
Circadian rhythm
Chronobiological research has revealed tight coupling of redox reactions to circadian rhythmicity (36). As mentioned above, peroxiredoxins are conserved markers of circadian rhythms (Fig. 5A) (24), and there is diurnal variation in GSH and cysteine redox states (9). Physiological functions such as neuronal excitability are regulated by rhythmic changes in the redox state, and there are numerous implications of rhythmicity to human health (121). This burgeoning field of research has been circumscribed as the circadian topology of metabolism (4), with clocks impacting on fitness in multicellular organisms, ranging in medical aspects from nutrition to chronopharmacology and chronotherapeutics (81). Novel chronoassays as the diurnal exhalome may facilitate practical applications (66).
Concluding Remarks: The Redox Code
Research in redox biology has experienced progress to new levels by emerging methods, which identify redox states and allow monitoring in time and space. Central reactions of energetics and metabolism are controlled through near-equilibrium NAD reactions. The structure and function of cells, including genomics and epigenomics, are controlled through nonequilibrium reactions of the cysteine proteome. The spatiotemporal organization of complex systems is coordinated through activation/deactivation cycles of H2O2 production. Redox changes are a common feature of aging, and disruption is a common factor in disease. Hence, application of the new redox tools to define targets and target interventions affecting the NAD and redox proteome systems opens a new horizon for therapeutics impacting a broad range of human disease. The concept of the redox code integrates spatiotemporal set points in cellular organization.
Footnotes
Acknowledgments
The authors are grateful to Dr. Young-Mi Go and Dr. Wilhelm Stahl for critical comments during article development. H.S. is a Fellow of the National Foundation for Cancer Research (NFCR), Bethesda, MD, and expresses gratitude for the longstanding research support as well as support by Deutsche Forschungsgemeinschaft, Bonn, Germany. D.P.J. is supported, in part, by NIH grants, ES023485, ES009047, AG038746, HL113451, and ES019776.