
Abstract
Introduction and Focus
O
The enormous variety of different chemical structures of oxidized phospholipids complicates their study considerably. Advances in our understanding have depended on the ability to separate heterogeneous mixtures of oxidized phospholipids derived from biological samples and elucidate their structures, which has largely relied on analytical techniques that can provide molecular-level detail such as mass spectrometry (MS), and developments in MS technology have been critical to the increased recognition of the importance of oxidized phospholipids.
This article focuses on the analysis of oxidized glycerophospholipids, and it explores why MS is such an informative and useful method to study them. Recent developments in MS technology that enhance its application to complex biological samples are described, and examples of the knowledge obtained regarding specific oxidized phospholipids and their levels in vivo are given. The major focus is on the use of liquid chromatography mass spectrometry to study oxidized phosphatidylcholines (PCs) and ethanolamines, reflecting the balance within the literature, but a brief mention of other approaches and phospholipids is included. The aim of the article is to give nonexperts interested in studying phospholipid oxidation an overview of both the potential and the complexity of MS approaches.
Types of Lipid Oxidation Products
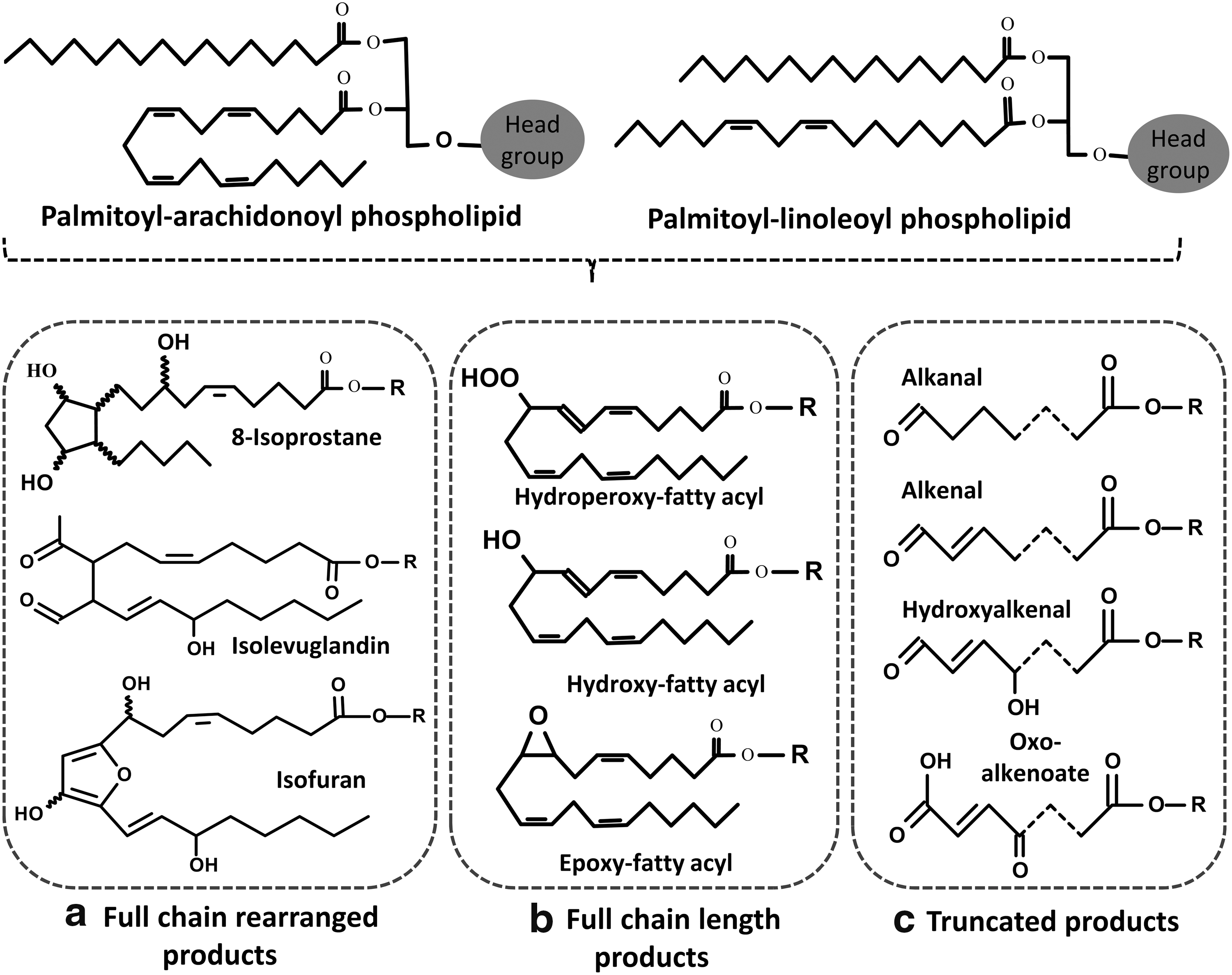
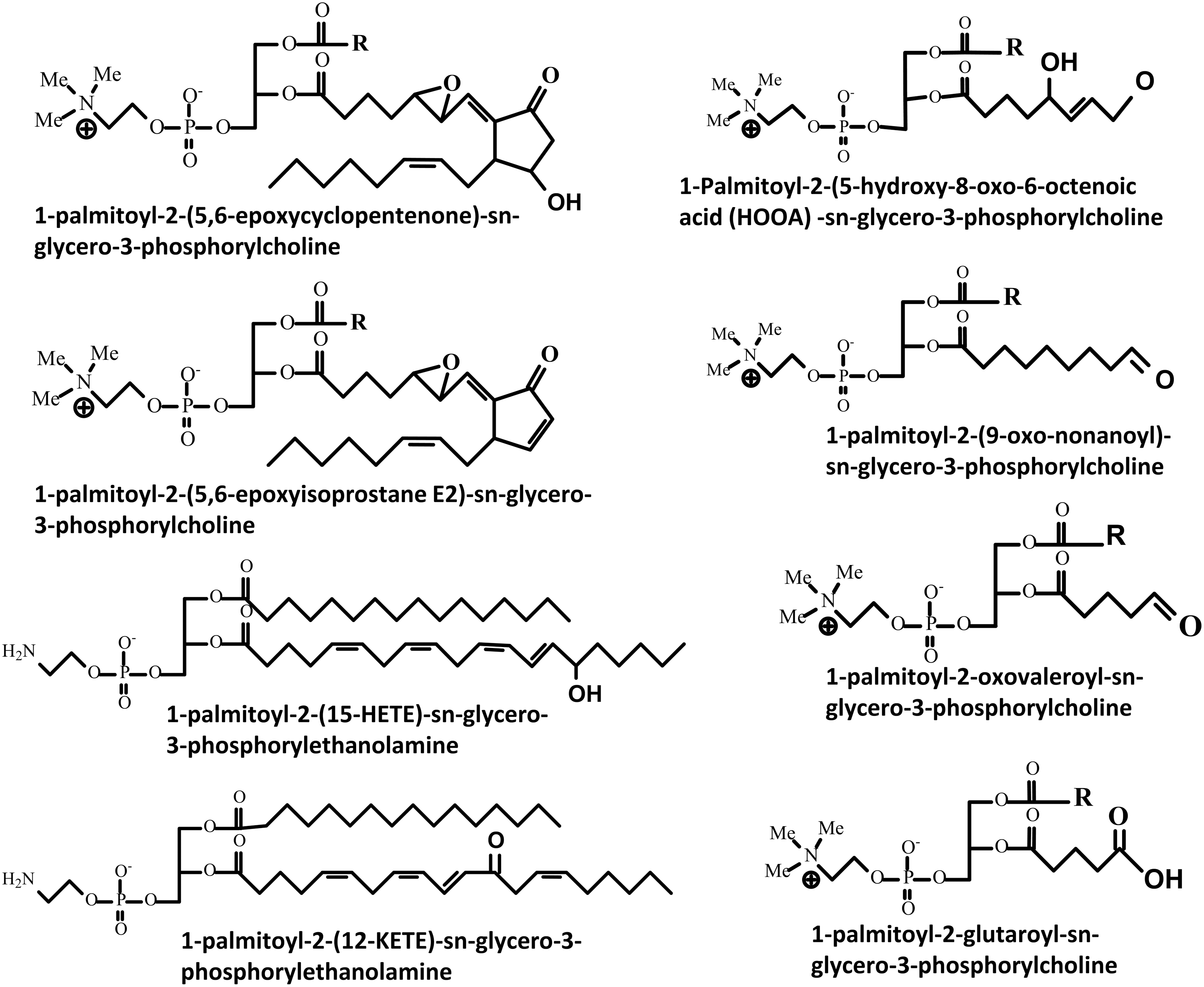
One of the challenges in the analysis of oxidized phospholipids is their extensive heterogeneity, which is a consequence of the large number of phospholipids that exist and the variety of oxidants that can modify them. The oxidation products formed from phospholipids depend on the type of oxidation (free radical versus two electron attack) and the structure of the phospholipid in question. Free radical attack leads to lipid peroxidation through well-characterized pathways that have been extensively reviewed (87, 94, 105, 115, 120). Phospholipids containing polyunsaturated fatty acyl chains (PUFAs), such as arachidonic, eicosapentaenoic, or docosahexaenoic acids, are most vulnerable to this sort of oxidative attack. In contrast, hypohalous acids (derived from the myeloid cell enzymes myeloperoxidase and eosinophil peroxidase) and some reactive nitrogen species can additionally attack lipids containing mono-unsaturated fatty acids, the vinyl ether bond of plasmalogens, or reactive headgroups such as phosphoethanolamine or phosphoserine. The extensive variety of possible products includes full-length oxidation products, chain-shortened phospholipids, and the corresponding fragments of the oxidized fatty acyl chains, of which the aldehydes malondialdehyde and 4-hydroxy-trans-2-nonenal are well-known examples (7, 8). The full-length oxidation products vary from simple modifications (hydroperoxides, hydroxides, epoxides, and ketones) to more complex products such as those resulting from cyclization (e.g., isoprostane-like structures and cyclopentenone rings) or further rearrangements (e.g., ring opening to yield isolevuglandins) (Fig. 1). The structures of some of the most commonly studied and detected oxidized phospholipids are shown in Figure 2.
Approaches to Phospholipid Analysis
Traditional methods for detecting lipid oxidation products can broadly be divided into those that depend on chemical reactions with reagents, immunoassays, or separation techniques (Fig. 3). Initial products of radical attack on phospholipids contain carbon-centered or oxygen-centered radicals, so electron spin resonance spectroscopy (ESR or EPR) with spin traps can be used to detect them (77). A recent example is the use of α-(4-pyridyl-1-oxide)-N-tert-butyl nitrone (POBN) to stabilize the radical products of docosapentaenoic acid (C22:5) in an online LC-ESR system (96). However, it remains difficult to apply this technique to complex biological samples, and a more common approach is to use colorimetric or fluorimetric assays for nonradical products. Most phospholipid oxidation products do not contain extended chromophores and cannot be detected in the visible range without a reaction with compounds that generate better chromophores or fluorophores. There are assays for the detection of hydroperoxide groups, such as iodometric titration (54), FOX assays (143), and isoluminol-dependent assays (80, 146). Commonly used assays for aldehydes or ketones are based on a reaction with 2,4-dinitrophenyl hydrazine (81, 137), cyclohexanedione, or pentafluorobenzyl hydroxylamine (81, 120). Many variations of these assays exist and have been used to investigate the occurrence of lipid oxidation in extracts of biological samples (120). Hydroperoxide-reactive fluorogens also offer the possibility of tissue imaging of lipid peroxidation. The hydrophobic compound diphenyl-1-pyrenylphosphine (DPPP) is oxidized fairly selectively by lipid hydroperoxides (owing to its localization in nonpolar areas) and has been used to demonstrate the formation of lipid peroxides in rat pulmonary microvascular endothelium during ischemia ex vivo (76). However, it is difficult to extend this approach to whole animal studies because of the toxicity of DPPP.
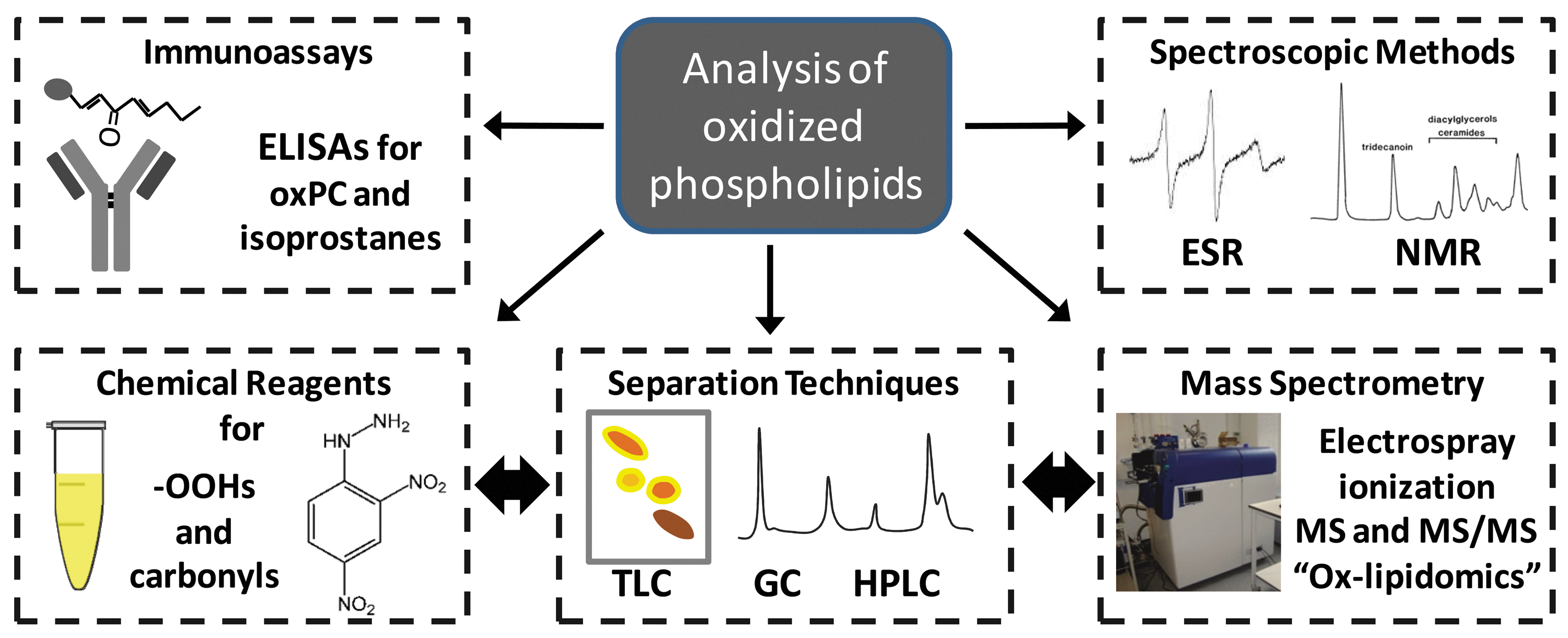
A limitation of all of these assays is that they are not specific for individual products, but rather for classes of products, so they are only suitable if a global measure of oxidative damage is required. Depending on the complexity of the sample, this issue can be partly addressed during chromatographic separation before detection. Basic methods such as thin layer chromatography continue to be used for separating different classes of lipids (70, 110, 135). Gas chromatography (GC) is a popular separation method for modified fatty acids and improved methods continue to be reported, for example, in separation of epoxy fatty acids from hydroxy and un-modified fatty acids (83). Coupling GC to MS provides additional structural information based on the fragmentation profile of the analytes. The ionization methods typically used in such instruments are high energy, such as electron impact or chemical ionization, and should not be confused with the low energy ionization methods discussed later. GC-MS is widely regarded as the best method for quantification of isoprostanes (17, 63) and has also been used for hydroperoxides and hydroxides of fatty acids (120, 130). However, it is less convenient for oxidized phospholipids, which must first be hydrolyzed to free fatty acids (150), so important structural information is lost. In contrast, high-performance liquid chromatography (HPLC) is ideal for separating oxidized and native phospholipids, and it is readily interfaced to a range of detector types. Moreover, it interfaces well to soft ionization MS, which, as discussed subsequently in this review, has advantages for analysis of phospholipid oxidation products.
Immunoassays such as ELISAs and radioimmunoassays offer an entirely different approach to detecting oxidized phospholipids. They are usually highly sensitive but depend on the availability of selective antibodies. Immunoassays for nonesterified lipid oxidation products such as F2-isoprostanes (139) and 8-iso-prostaglandin F2α (63, 137) have been available for many years, but there are several problems with them (130) and the best method for analysis of free isoprostanes is either GC-MS (67, 79, 103) or LC-MS methods (93, 150). Natural antibodies of the IgM class that recognize phosphocholine-containing lipids (e.g., T15; (22) and refs therein) were found to have identical variable regions (and therefore antigens) to more recently characterized natural antibodies (the E0 series), found at high levels in ApoE-deficient mice (29). The antibody E06 has been thoroughly characterized and found to recognize both free oxidized PCs (especially POVPC) and PC adducts to LDL and albumin. A chemiluminscent ELISA for oxPC-modified LDL has now been developed using E06 and applied extensively to studies of lipid oxidation in cardiovascular and other diseases (127).
While all of these methods yield useful information about lipid oxidation in general, most of them cannot identify specific molecular oxidized phospholipid species, which is important for understanding the biological effects. To achieve this level of detail, advanced LC-MS/MS approaches are the best option, and their application will be discussed in the next few sections.
MS in Modern Oxidative Lipidomics
Electrospray ionization (ESI) and matrix-assisted laser desorption/ionization (MALDI) are considered “soft” (low energy) ionization techniques, and they have been used for more than 50 years for the analysis of labile biomolecules, including phospholipids (33, 95). They are a convenient method to identify oxidized phospholipids, most of which are readily ionizable. An important advantage compared with GC-MS is that analytes can be detected directly, without the chemical manipulation necessary to release the fatty acid chains and derivatize them to generate the volatile species required for GC (116). Thus, these soft ionization methods tend to be simpler, less open to handling artifacts, and information on the intact phospholipid structure is not lost. However, it is worth noting that in some cases it is currently impossible to distinguish the positional distribution of oxygenated groups (e.g., in oxygenated derivatives of arachidonic acid such as LTB4, Hepoxilin A3 and 5, 12, and 15-HpETEs) using these methods, without resorting to hydrolysis and analysis of the fatty acids.
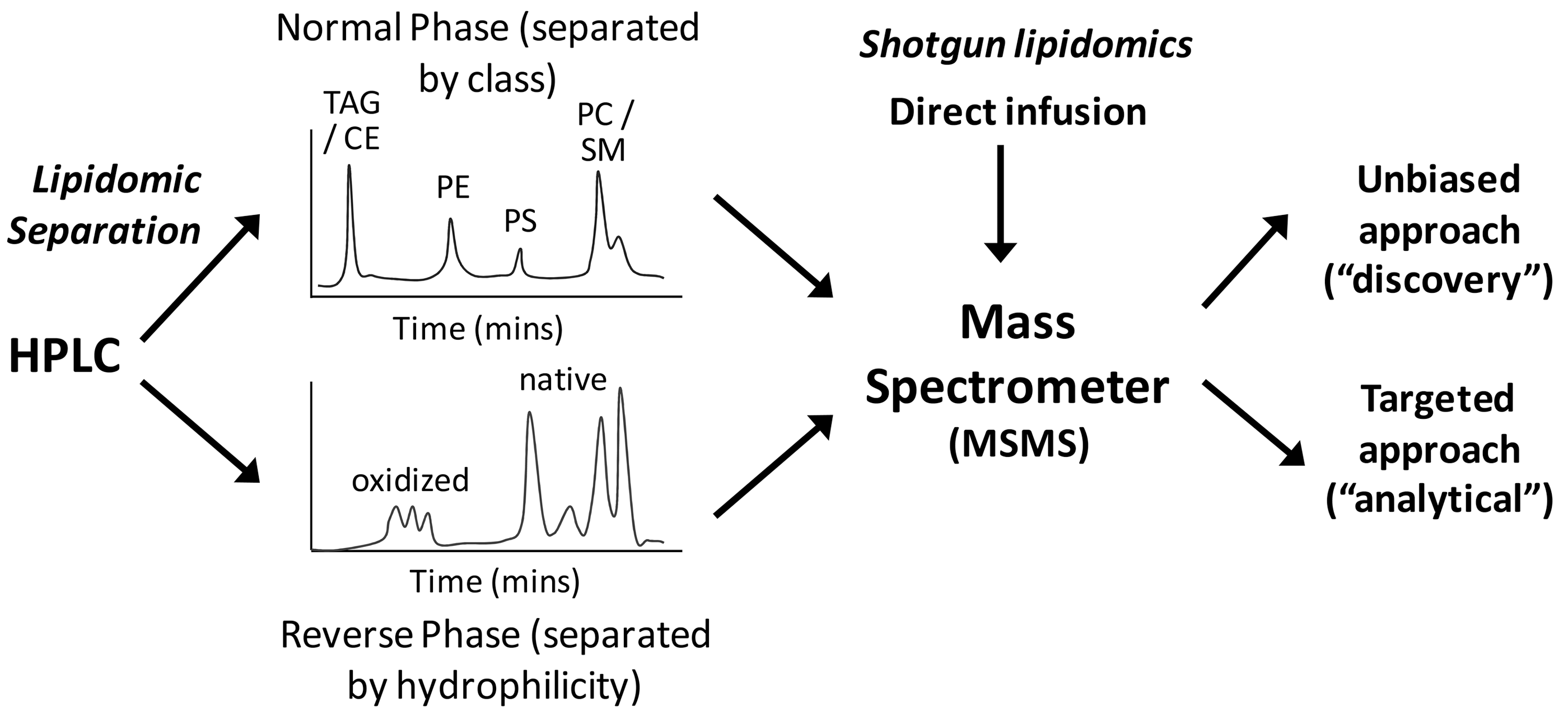
The ability of ESI and MALDI MS to analyze multiple components in the sample simultaneously allows the profiling of many lipid classes in biological tissue in parallel, and has led to the development of shotgun lipidomics, in which the sample is introduced into the mass spectrometer without separation, and the components are scanned and, for MSMS modes, fragmented (43). While this is a relatively powerful technique, the variety of lipids is so large, and the ratio of oxidized to un-modified lipids usually so small, that preseparation techniques are important to ensure representative profiles are obtained; otherwise, minor signals can be obscured or suppressed by more abundant molecular ions. As ESI requires the infusion of the sample in solution, it can be coupled very readily to HPLC, with normal phase and reverse phase columns being most common, although other column types have also been used (Fig. 4). Thus, LC-MS(MS) has become the method of choice for most lipidomics work, although in some cases the higher throughput achievable with shotgun approaches is desirable; there are several informative reviews on this topic (9, 43, 58, 142). The use of MS in biomolecule analysis has advanced enormously in the past 10 years, especially not only in terms of the sensitivity, resolution, and scanning speed of mass spectrometers but also in terms of identifying fragmentation patterns that can be used for smart scanning approaches (111), and these advances are now being translated to oxidative lipidomics (73, 90). For example, LC linked to high-resolution MS has been used to demonstrate the occurrence of oxidized PCs and ethanolamines in plasma from control and obese human subjects (21).
The application of ESI in lipidomics far exceeds the use of MALDI, largely because of the potential for interfacing directly with liquid chromatography and simplicity of sample preparation; MALDI usually requires co-crystallization of a matrix with the sample, the quality of which affects the signal. This means that MALDI is intrinsically less amenable to absolute quantification than ESI. However, procedures for MALDI-MS of phospholipids and oxidized or chlorinated phospholipids have been developed (32, 112, 122): Recently, MALDI-MS has been applied to a wider range of oxidized phospholipids (26, 27, 151) and has been used to investigate the fragmentation of cardiolipin by copper in a mouse model of the copper overload condition Wilson's disease (151), as well as demonstrating the occurrence of truncated oxidized PCs in spinal cord tissue of Sprague–Dawley rats (124). It has been argued that MALDI has advantages compared with ESI for biological samples, as it is more tolerant of the presence of salts or detergents, whereas ESI usually requires sample cleanup or the application of LC. MALDI has also been the technique most commonly used for MS imaging to gain information on spatial distributions of molecular species across tissues, whereas ESI requires extraction of the tissue.
Principles of Soft-Ionization MS Analysis
To be able to make best use of information on oxidized phospholipids obtained from MS, it is necessary to understand some basic principles of the techniques involved. Mass spectrometers contain mass analyzers, which discriminate between ions based on their mass-to-charge (m/z) ratio. In a tandem mass spectrometer, which can be used to gain structural information, two analyzers are connected together via a third component, the collision cell; a molecule selected in the first mass analyzer, the precursor (parent) ion, is broken down by collision with an inert gas (He, N2, Ar, etc) in the collision cell into fragment (daughter) ions that are analyzed in the second mass analyzer. The same effect can be obtained in ion trap mass spectrometers by capturing all of the ions, selecting the one required for further analysis by ejecting all the others, fragmenting this in the trap by collision with an inert gas, and then scanning the resulting ions out of the trap to obtain the mass spectrum. There is now a bewildering array of different mass analyzers, connected together in an even wider number of combinations, and some additional methods of ion fragmentation. Each has its own merits, and further specific information can be found in a number of reviews and books (and many on-line resources) (4, 15, 23, 39). For the purposes of this review, it is important to understand the operation of the quadrupole (Q) mass analyzer, as this is the most common first analyzer in tandem mass spectrometers. It operates by allowing only ions of a specific m/z value to pass through to the detector. This can be scanned through the m/z range of a defined mass window, but as this scanning is slow and of a relatively low resolution, quadrupoles are more commonly used as mass filters by setting them up to select individual ions rather than in scanning mode. An important recent advance has been in the wide availability of high-resolution (ability to distinguish two ions of similar mass), high-mass accuracy (ability to accurately and reproducibly measure the mass of an individual ion) instruments that routinely achieve resolutions of 40–100 k and mass accuracies of 0.5–5 ppm. This is important in the analysis of oxidized phospholipids, as it allows oxidized phospholipids to be distinguished from native phospholipids of the same nominal mass (such as PC18:0/18:0 and PC16:0/18:2-OOH) at high resolution, owing to the unusual negative mass defect of oxygen, which has an accurate mass of 15.9949 amu (138).
Every ion of a different m/z yields a unique signal in the resulting spectrum, so this technique is ideal for investigating complex mixtures. Although there may be compounds with different elemental compositions but very similar masses (isobaric compounds), these can often be differentiated by high-resolution instruments, as mentioned earlier. In contrast, isomeric compounds, which have the same elemental composition but different structures, can only be distinguished by fragmentation, or separated chromatographically (102), or using ion mobility MS. Ion mobility mass spectrometry (IMS) is an increasingly valuable technique for lipid analysis, after the recent introduction of commercial technologies (such as the Synapt™ mass spectrometer and the SelexION™ source). Ion mobility separates ions based on their shape (more correctly on their average collisional cross-section) and has proved particularly useful in lipidomics for distinguishing isobaric lipids from different classes or of different double bond configurations (2, 14, 56, 66, 69, 93, 109). It also has great potential for removing interfering chemical background from the mass spectra, allowing better assignment and more sensitive detection, and has proved particularly useful in mass spectrometric imaging applications (16, 52, 144). This technique has not yet been applied specifically to oxidized lipid species, although there are some reports that include oxidized lipids (51), but it will have great potential for adding an additional dimension of separation. As oxidation of a lipid or phospholipid to form a hydroperoxide or a hydroxide involves addition of oxygen to the native structure, the mass of the molecule is changed, which can readily be detected by MS (118). Likewise, modification through addition of HOCl to form chlorohydrins (53) or NO2 to form nitro-fatty acids (3, 65) results in new signals at higher m/z. Breakdown products, including lysolipids and chain-shortened phospholipids, appear at lower m/z windows.
The ionization mode, and to a lesser extent the solvent system, determine the lipid class that can be observed: Phosphatidylcholines (PC), sphingomyelins (SM), and phosphatidylethanolamines (PEs) are observed in positive ion mode, whereas phosphatidylserines (PS), phosphatidylinositols (PI), phosphatidylglycerols (PG), and cardiolipins (CL; diphosphatidylglycerols) can best be observed in negative ion mode (Table 1). However, adjustment of the solvent systems with ionization modifiers can enable detection in the opposite ionization mode if required: for example, although PC has a constitutive positive charge on the choline moiety, phosphocholine-containing lipids can be observed in negative ion mode as formate or acetate adducts (20). As PC is both very abundant and easily observable in either mode, much research had previously focused on identifying oxidation products of unsaturated PCs (20, 116), although to address this knowledge gap increasing studies on oxidation of phopholipids with other headgroups are being undertaken, including PE, PS, and CL (26, 70, 74, 132). The principles of fatty acyl group oxidation are common to all the phospholipids, and it is well established that hydroperoxides, hydroxides, epoxides, and chlorinated species can be detected, according to the mass increments and fragmentation patterns.
However, when using fragmentation patterns to identify the lipids present, it is important to be aware that the ionization state and the counter ion present alter the fragmentation. Protonated glycerophospholipids undergo different collision-induced decomposition to sodium adducts of the same lipid, owing to their differing behavior in gas phase chemistry (20, 34); likewise, negatively charged formate and acetate adducts of PC fragment differently to the same PC observed under positive ionization conditions. Characterization of fragmentation patterns for oxidized phospholipids of interest facilitates the identification of these analytes in biological samples and is essential for the development of targeted MS approaches, as described in the next section. In many studies, fragmentations have been determined experimentally (99), although once the principles for one type of oxidized product have been ascertained, they can be used to generate theoretically expanded lists for other species of analogous structure (85).
Types of Targeted Scanning Routines for Oxidized Phospholipids
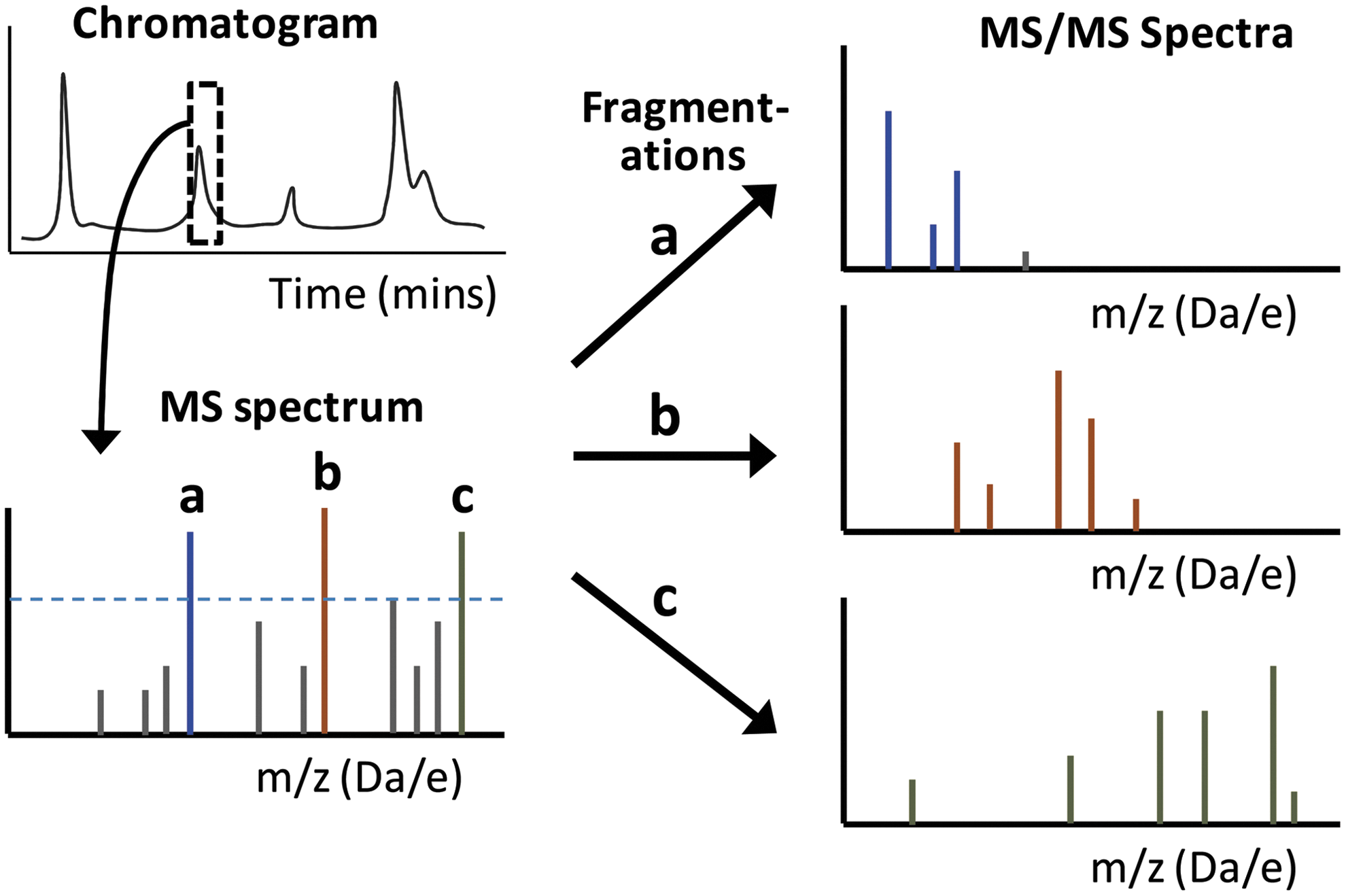
Tandem MS allows the use of targeted scanning routines, which are important as they facilitate selective identification and often quantification in more complex samples (58). In much of the early research on identification of lipid oxidation products, individual lipids or simple mixtures of lipids were oxidized in vitro; thus, it was feasible to select and fragment individual parent ions to identify the analyte by the pattern of fragment ions. This MS approach is referred to as product ion scanning (Fig. 5), and it has proved extremely useful for structurally characterizing many phospholipid oxidation products. For example, important early work on the identification of chain-shortened PCs and PCs with esterified isoprostanes in atherosclerotic plaques depended on this approach (48, 97, 125, 140, 141), and it continues to be used (55). It has been used to identify novel lipid oxidation products such as isolevuglandins as well as to determine mechanisms of oxidative fragmentation (68, 104). In ion trap instruments, multiple sequential fragmentations (MSn) can be carried out to provide more detailed analysis of the structure in question (149). Product ion scanning also provides information on diagnostic fragment ions for individual oxidized products, which can subsequently be used in the targeted scanning approaches. Several reviews have described typical fragmentation products and have provided useful summaries of the information (20, 99, 117). In these LC-MSMS lipidomic approaches to discover oxidized lipids present in complex samples, data-dependent acquisition is often used, whereby throughout the LC run, ions for fragmentation and product ion scanning are selected based on intensity, which is prone to missing low-abundance ions that may be biologically important (Fig. 6). Hence, targeted methods with the potential to overcome this problem have been developed.
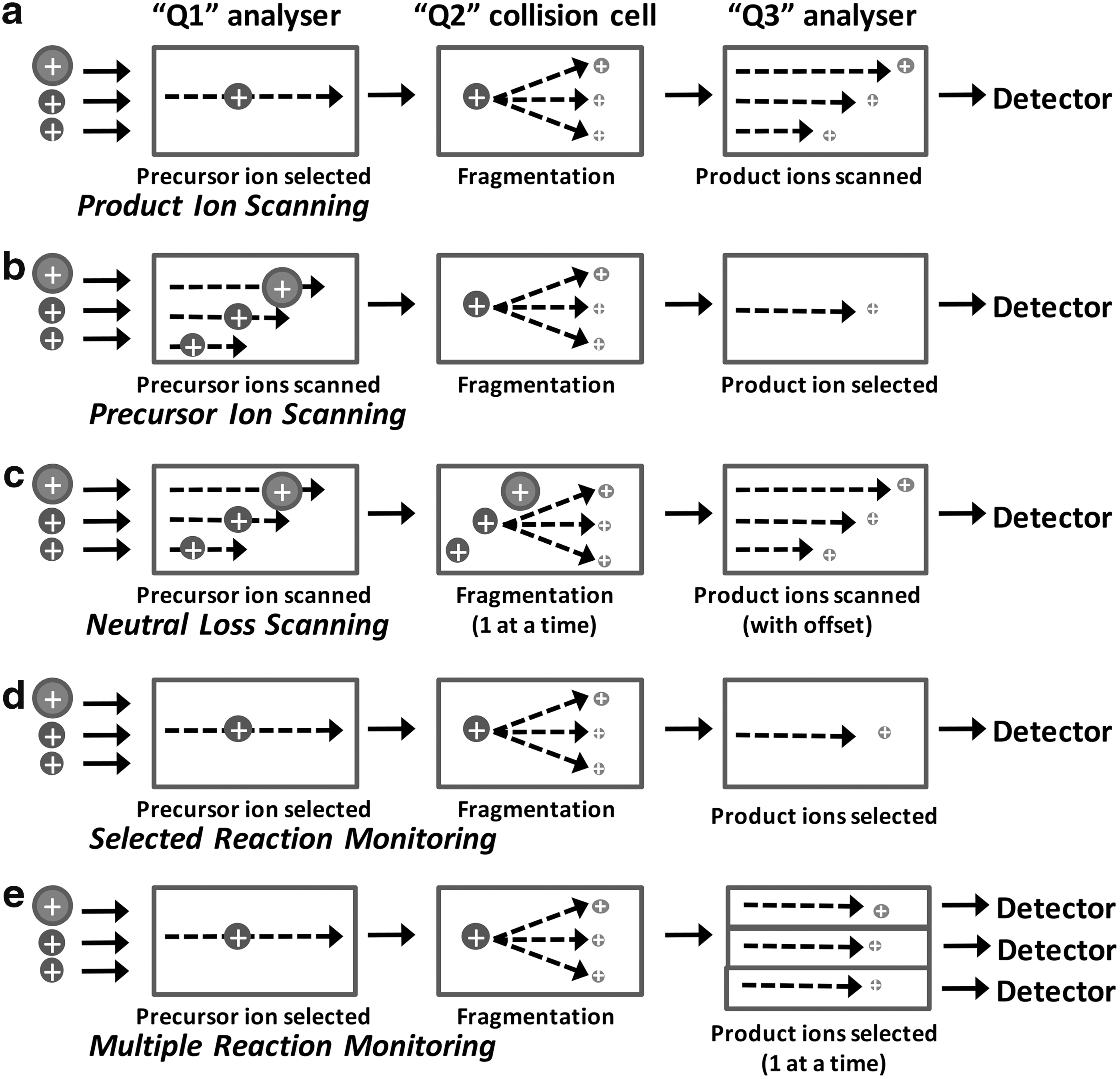
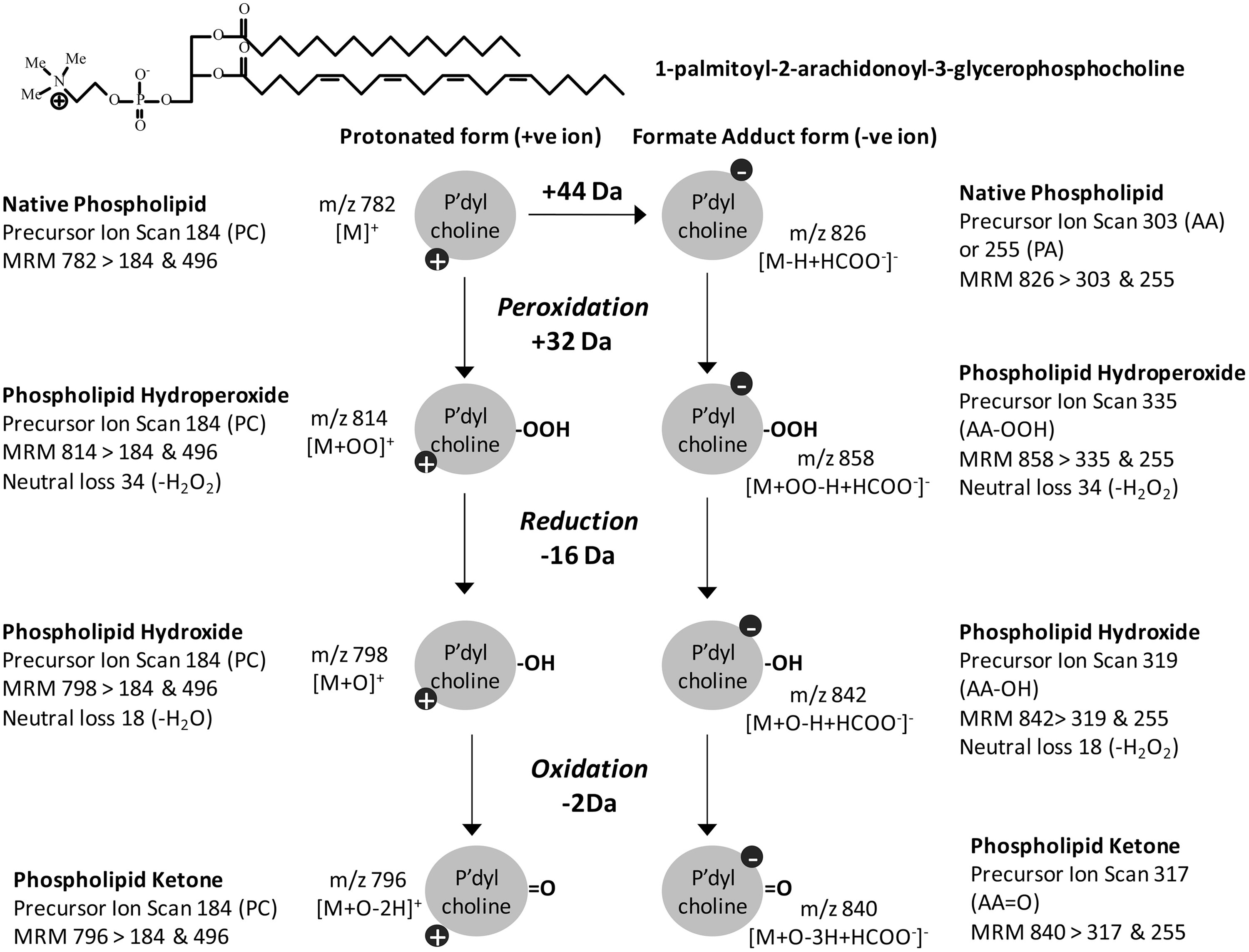
There are three main targeted MS approaches, which have different levels of selectivity (Fig. 5). Precursor ion scanning identifies precursors (parent ions) that fragment to give a specific fragment ion (daughter ion) of interest by scanning through the masses in the first analyzer with the second analyzer set to detect a specific m/z. Neutral loss scanning identifies precursors that fragment by loss of a specific uncharged fragment, regardless of the mass of the parent ion. This routine scans the first and second mass analyzers with a fixed mass offset between them corresponding to the mass of the neutral fragment lost. Both of these methods can identify families of analytes containing a particular structural feature. For example, precursor ion scanning for m/z 184 identifies phosphocholine-containing lipids such as SM or PC (126), while scanning for neutral loss of 18 amu, corresponding to water, and indicates the presence of hydroxyl groups (119). In contrast, selected single reaction monitoring (SRM) should be specific for a single molecular species, as both the m/z of the parent ion and a diagnostic fragment ion are set, in the first and second analyzers, respectively. The specificity of the identification can be improved by selecting multiple “transitions” from precursor to fragment ions, assuming that several strong fragmentation products occur; this technique is known as multiple reaction monitoring (MRM), although MRM has also come to refer to SRM experiments on multiple precursors run in parallel. The principle in all these methods is essentially the same, but SRM and MRM can be considered specific cases of precursor ion scanning (for a single precursor of interest) (43). The fundamental difference between SRM/MRM and the scanning routines is that the latter can be used as discovery approaches, searching more broadly for types of products [hydroperoxides, chlorohydrins, hydroxides (119)], whereas reaction monitoring routines tend to be used for analytical work when the presence of a specific analyte has been established and a quantitative approach is needed. SRM and MRM have much higher duty cycles (more time is spent collecting data on the species of interest), so they generally have higher sensitivity than scanning experiments. Consequently, they are well suited to quantification of oxidized lipids. Some typical diagnostic routines for 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphocholine are given as an example in Figure 7.
Application of Targeted MS Methods for Oxidized Phospholipids
Although the MS routines described earlier have been known for many years (as long as tandem MS instruments), their application to the study of oxidized phospholipids has expanded greatly recently as new instrumentation and software have become available, and appropriate diagnostic ions have been identified. This section reviews recent developments in the methodology in model biological systems.
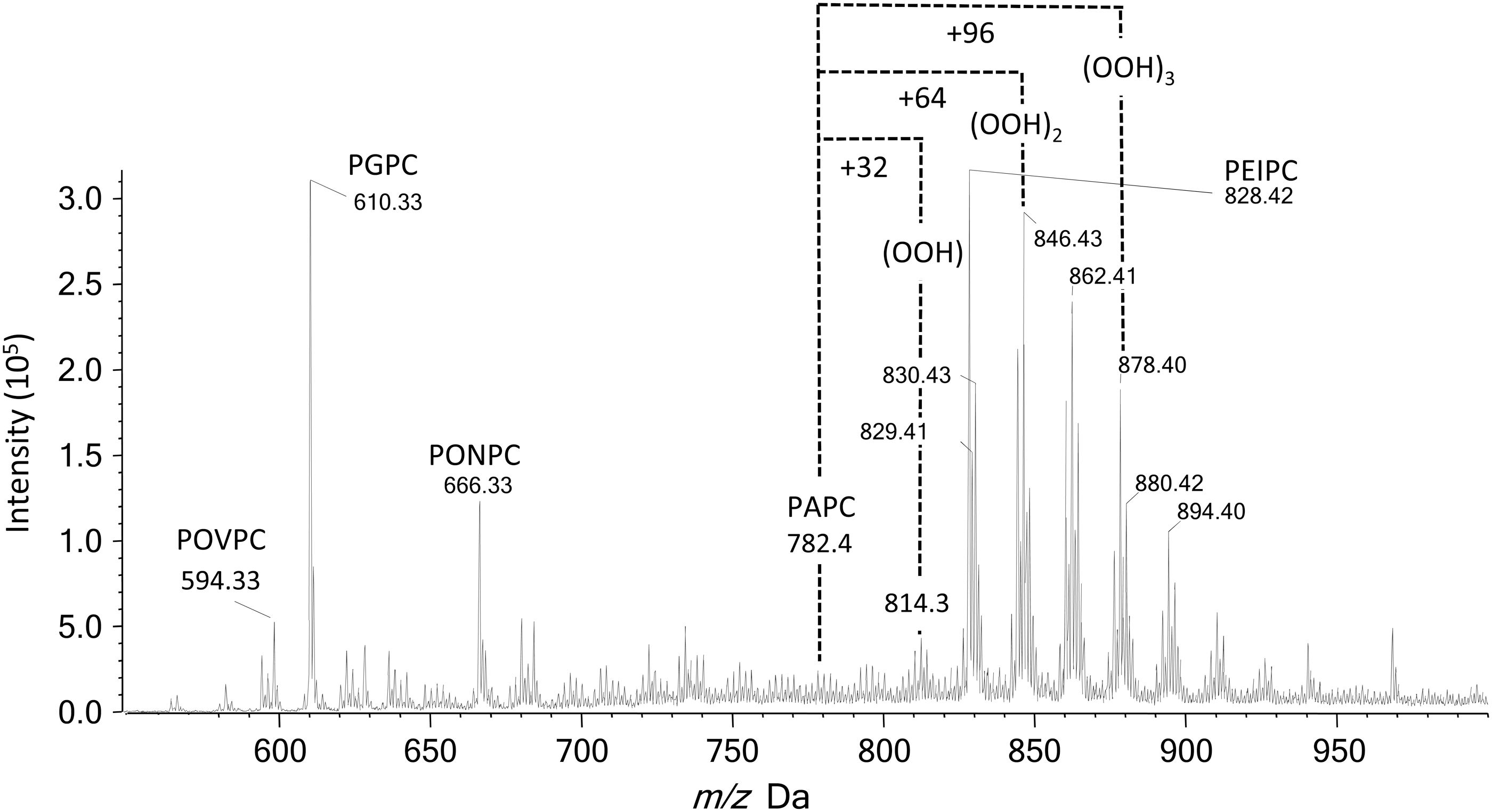
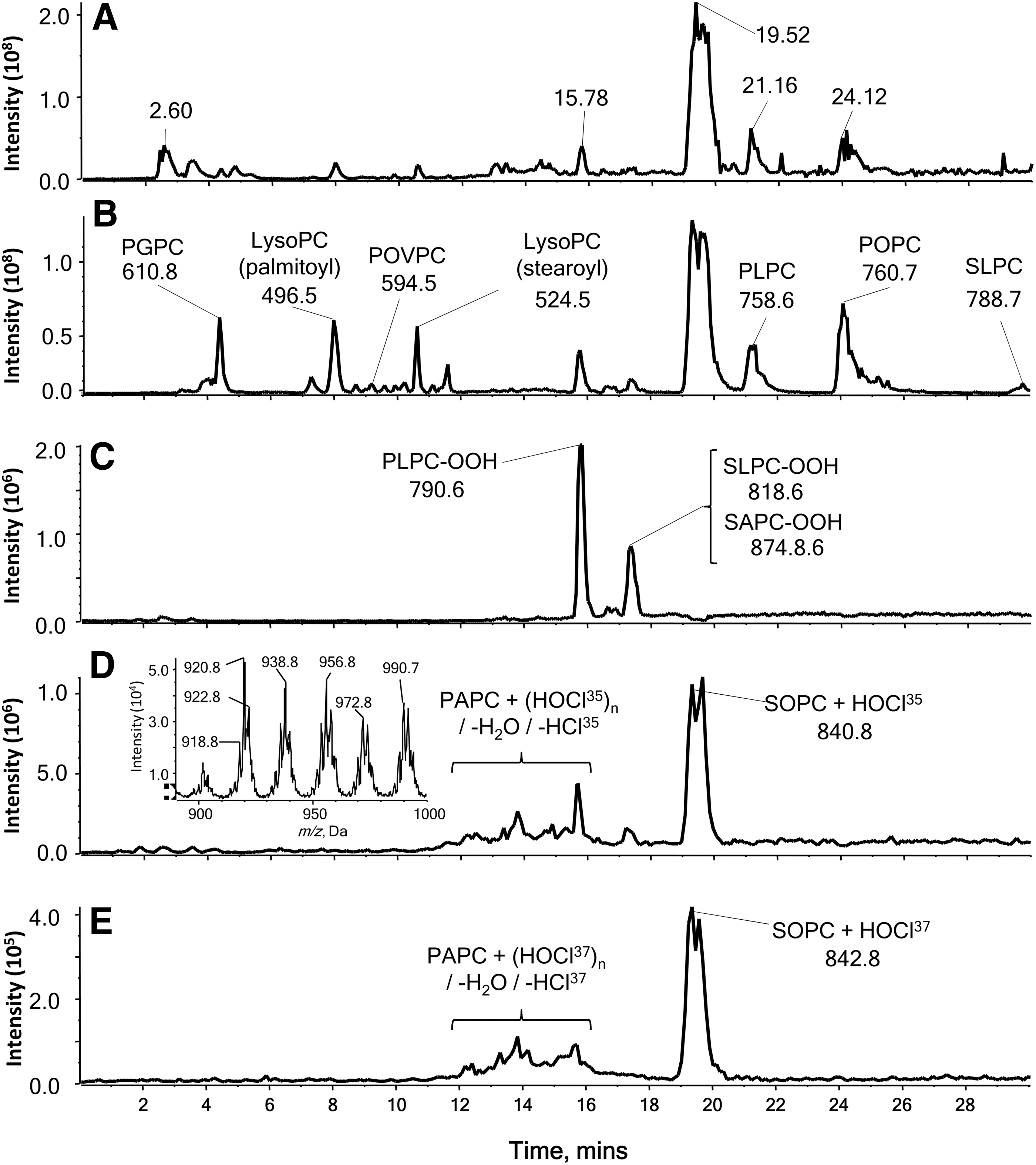
Much of the work carried out to date has focused on the analysis of PCs, as these are abundant phospholipids and have a constitutive positive charge, so they yield strong ions in the positive ion mode. The most common diagnostic scan is for m/z 184.1 (the phosphocholine headgroup) in the positive ion mode, either as a precursor ion scan or transitions from specific parent ions (SRM). Scanning for precursors of m/z 184.1 allows detection of all PCs, not just oxidized forms (Fig. 8), and also detects SMs as they contain a phosphocholine moiety, but nevertheless this is useful in studies wanting to characterize the range of native and oxidized species present (46, 98, 119). In contrast, SRM and MRM can select specific oxidized forms. For example, SRMs of m/z 790>184 and 818>184 have been used to quantify monohydroperoxides of 1-palmitoyl-2-linoleoyl-PC and 1-stearoyl-2-linoleoyl-PC in healthy human plasma by comparison with a calibration curve of synthetic standards (49). Similarly, hydroperoxyoctadecadienoyl phosphatidylcholine (HpODE-PC) and hydroxyoctadecadienoyl phosphatidylcholine (HODE-PC) were analyzed in rat and human plasma by transitions from the parent ions (790 and 774, respectively) to phosphocholine at m/z 184 (148). Other types of oxidized PC products, including the epoxyisoprostane PEIPC (m/z 828.6>184) and lysolipids such as 1-palmitoyl-2-hydroxyl-sn-glycero-3-phosphorylcholine m/z 496.3>184), have also been detected by SRM (18). More recently, an extensive list of SRM transitions was reported for the quantification of oxidized products derived from the most abundant species of phosphocholines in human dermal fibroblasts subjected to UVA irradiation, which clearly caused increased levels of many of these compounds (40). However, quite a few of these transitions were not specific for an individual oxidized product, owing to the existence of isobaric or isomeric PCs. This is a limitation of detection in the positive ion mode using a single transition to m/z 184. The specificity can be improved by MRM where additional product ions resulting from the loss of water (−18 Da) are used for hydroxides and loss of H2O2 (34 Da) for hydroperoxides. Uchikata et al. used this approach to measure oxidized PCs in liver from mice fed on a high-fat diet: As an example, the monoperoxide of SAPC was monitored by transitions from m/z 842.6 to 184.1, 808.4 (loss of H2O2), and 824.4 (loss of H2O) (136). It is important to note that the loss of water is not a very specific transition, as it occurs for hydroxides and hydroperoxides, although it has been used in an SRM approach (50).
While positive ion detection works very well for PCs, most other glycerophospholipids can be more readily observed in the negative ion mode, and this also has an advantage that a wide range of fatty acyl fragments can be observed relatively strongly in the MSMS spectra and used as reporters in precursor ion scanning. This approach has been applied effectively in several studies on the generation of oxidized products of PEs by lipoxygenases (89, 90). Precursor ion scanning for phospholipids containing hydroxyeicosatetraenoic acid (HETEs) (fragment ion at m/z 319.2) has been carried out and identified precursor ions at m/z 738, 764, 766, and 782 that were found to correspond to a family of plasmalogen PEs containing 15-HETE (74). Similarly, scanning for precursors of ions with m/z 317.2 can be used to identify KETE-containing phospholipids (42). Subsequently, SRM methods were set up and applied for a wide range of PEs from several cell types; appropriate selection of collision energies also allowed positional isomers (e.g., 5-HETE, 12-HETE, 15-HETE) to be analyzed (82). In fact, PCs containing equivalent oxidative modifications of the fatty acyl chains can also be analyzed in the negative ion mode, if solvents containing acetate or formate are used to generate [M-H+HCOOH]− or [M-H+CH3COOH]− adducts, respectively (82, 85), and an advantage is that two transitions of comparable intensity, one for each fatty acyl chain, should be detectable. The corresponding neutral loss of fatty acyl fragments is observed in the positive ion mode, and facilitates structural determination, though these signals are generally weak and very dependent on lipid structure. This neutral loss can be used as a reporter scan, but is generally less sensitive and selective than precursor ion scanning.
Recently, there have been studies on detection of oxidized PS by targeted MS approaches (70, 71, 131, 135). Lipids containing the phosphoserine headgroup can be selectively detected by a neutral loss of 87 amu, and this has been applied for the detection of native PSs and PSs containing an oxidized fatty acyl chain (72). However, it has also been observed that the amine moiety of the headgroup is susceptible to oxidation, leading to formation of glycerophosphoacetic acid (GPAA). Alternative selective scanning procedures for this modification to the head group have been reported, involving a neutral loss of 58 amu or scanning for precursor ions of m/z 137.1, and used to detect PS oxidation in keratinocytes subjected to radical oxidation in vitro (Fig. 9) (72).
It is well recognized that lipids containing hydroxides can be identified by loss of 18 Da and hydroperoxides by loss of 34 Da (1), and some studies have used the resulting transitions for SRM or MRM with observation of the charged fragment, as described earlier. In addition, these characteristic fragmentations can be used in neutral loss scanning to identify all analytes containing hydroperoxide moieties. This has been demonstrated for lipids oxidized in vitro (117, 119) (Fig. 10), but has not as yet been much utilized in biological samples, as neutral loss scanning tends to be less sensitive than precursor ion scanning or SRMs. The same is true for the use of the neutral loss of 36 Da (H35Cl), which has been suggested as diagnostic for chlorinated ions such as chlorohydrins of PCs. The presence of chlorine can be checked by neutral loss of 38 Da for the isotopic H37Cl form; this profile should match that of neutral loss of 36 Da, although weaker owing to the 35Cl:37Cl isotope ratio (78, 92). However, caution is required, as bis-hydroperoxides may also undergo sequential loss of two water molecules, corresponding to a decrease of 36 Da (50).
Novel Strategies for Detecting or Quantifying Oxidized Phospholipids
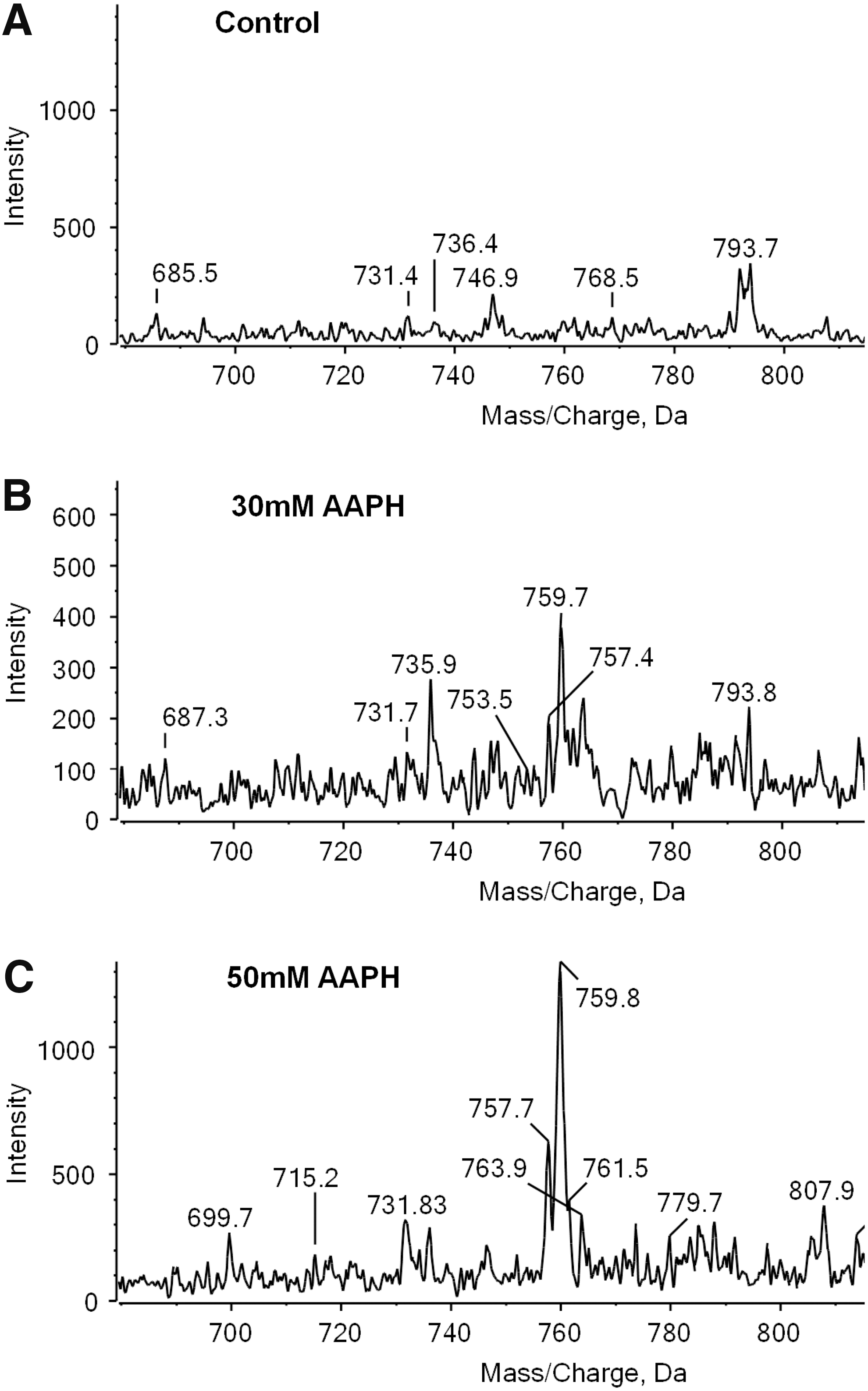
There is increasing interest in oxidation products of PEs, which also commonly contain unsaturated fatty acids. Although the neutral loss of 141 amu can be used to identify PEs, this targeted approach underestimates contributions from vinyl ether containing PEs, which fragment differently owing to the more labile nature of the vinyl ether bond. This can be overcome by chemically labeling the PE headgroup with 4-(dimethylamino)benzoic acid, which fragments to yield a strong signal at m/z 191.1 that can be used for precursor ion scanning (152). This fragmentation is favored compared with the loss of the ether-linked chain, and therefore plasmalogen PEs are also detected. An interesting application of this labeling approach is that isotope-labeled DMABA (D0, D4, D6, and D10) can be used for differential isotope labeling of samples, allowing relative quantification of the PE profiles in different samples by precursor ion scanning for m/z 191.1, 195.1, 197.1, or 201.1, respectively. This has been used to demonstrate changes in profile of PEs extracted from RAW264.7 cells and subjected to oxidation with the radical initiator AAPH in vitro (153). An ethyl-labeled PC hydroperoxide has also been synthesized with the objective of following structural changes to hydroperoxides in biological systems, in this case human blood (108). The phospholipid could be monitored selectively by scanning for precursors of the ethyl-labeled phosphocholine headgroup at m/z 198. Although this approach cannot be used in vivo, as it involves a synthetic lipid rather than a chemical labeling protocol, it represents an interesting method for studying metabolism of individual oxidized PCs in vitro, as a contrast to the use of expensive isotopically labeled lipids.
An interesting method for improving the extraction and enrichment of oxidized phospholipids in LDL has been recently reported by Hinterwirth et al. (46). Gold nanoparticles (GNPs) with anti-OxLDL antibodies immobilized on their surface were used to trap oxidized LDL selectively from plasma samples, before extraction of the lipids and analysis of the oxidized phospholipids by precursor ion scanning and SRM for transitions to m/z 184.1. Four different antibodies were tested: E06, an anti-Cu oxidized LDL antibody, an anti-MDA-LDL antibody, and an anti-carboxymethyllysine-LDL antibody; all the GNP-Ab conjugates showed specific binding of oxLDL and significantly enhanced the levels of oxidized phospholipids detected in plasma. This methodology also has the advantage that it can be used to measure dissociation constants for oxidized phospholipids, thus allowing the antigenicity of multiple lipid biomarkers to be screened. Other enrichment strategies compatible with MS have also been reported to improve the analysis of oxidized lipids in complex samples. For example, aldehyde products of fatty acyl chain oxidation have been trapped using hydrazine base reagents conjugated to solid surfaces such as nanoparticles (41), and the acidity of carboxylic acid oxidation products has been exploited using a nanoparticle enrichment strategy (123).
Advances in Chromatographic Separation of Oxidized Phospholipids
In addition to the advances in scanning approaches, there have been new developments in chromatographic separation for phospholipid oxidation products. LC-MS(MS) of oxidized phospholipids is commonly carried out with conventional (4.6 mm i.d.) or microbore (1 mm i.d.) columns, in contrast to proteomics analysis where nanoflow is commonly used; Lee et al. (60) demonstrated that nano-flow LC-MSn was effective for separating a variety of native and oxidized phospholipids. They used a laboratory-prepared 7 cm length silica capillary of 75 μm i.d. containing ODS-P C18 resin and gradient elution from water:acetonitrile (10:90 v/v) to acetonitrile: methanol isopropanol (20:20:60 v/v), and with data-dependent CID were able to detect the presence of 139 oxidized PLs species in commercially available oxLDL. The chromatographic separation achieved by this method appeared to be excellent, despite the column being packed in-house. Several types of nano-flow columns with C18 resins are available from commercial suppliers, and there is no reason that these could not be used more extensively in ox-lipidomics. Two-dimensional chromatography can also bring some significant benefit in increasing sensitivity (106), although at the price of more extensive sample manipulation and data generation and analysis.
A completely different approach to improving separation of oxidized phospholipids was taken by Uchikata et al. (136), using supercritical fluid chromatography (SFC) on a 2-ethylpyridine column. SFC uses carbon dioxide that is cooled and pressurized to maintain it in a supercritical fluid state as the mobile phase; it is suitable for nonpolar, labile analytes and is able to separate chiral compounds. For interfacing with MS, methanol containing 0.1% ammonium formate was used as a modifier solvent, which is essential for generating ions of the analyte. An advantage of SFC compared with reverse-phase chromatography was that hydroxides, epoxides, and hydroperoxides of PC could be separated; under the conditions used, hydroxides eluted at 5–6 min, epoxides eluted from 7.5 to 9 min, and hydroperoxides eluted at 10–10.6 min. While this method offers potential for advanced separation and analysis of full-chain oxidation products, it requires specialized chromatographic equipment and cannot yet be considered a routine analytical technique, although commercial systems are available.
Knowledge on Disease Physiology from Lipidomics Approaches to Lipid Oxidation
A critical question in the development of methods to identify and measure oxidized lipid products is to what extent they have been applied to obtain novel information about physiological systems. Fully targeted MRM-based methods have been in use for a number of years and there are a substantial number of publications related to physiology and disease, especially for nonesterified lipid oxidation products (oxylipins) (19, 121, 147, 154), but the semi-targeted methods are newer and currently there are fewer applications. As oxidized phospholipids are formed in inflammatory situations, many of the applications have been to cardiovascular conditions, such as ischemia-reperfusion injury and atherosclerosis, or infectious diseases, leading to systemic or chronic inflammation.
Oxidized phospholipids are known to influence inflammatory processes, and, consequently, several studies have used MRM to quantify their levels in disease and control conditions. For example, POVPC, PGPC, and PEIPC were found in macrophages infected with Mycobacterium bovis using transitions from the native masses to 184; using relative quantitation, the level of PEIPC, in particular, was found to be increased approximately 3.5-fold, and lyso PC also increased (18). The antibody E06 was used to investigate the occurrence of oxidized phospholipids in the less controlled lepromatous lesions, compared with the well-contained tuberculoid leprosy lesions, demonstrating the value of these complementary methods. Although absolute quantification was not carried out, the analysis of these oxidized PLs by targeted MS provided evidence of their role in controlling inflammatory outcomes during infection and the similarity of these inflammatory lesions with atherosclerotic lesions (18).
Nakanishi et al. (85) used MRM in the negative ion mode to detect oxidized PCs, with transitions from their native m/z to those of both the sn-1 and sn-2 chains (full length and chain-shortened respectively). The use of two or, in some cases, even three transitions gave extra confidence in the identification, but a disadvantage of analysis in the negative ion mode is that the overall sensitivity is often lower than in the positive mode. The transitions were based on theoretically expanded data sets; that is, they identified transitions of each PL and oxidation type, and extrapolated them to analogous oxidized phospholipids of different chain lengths. The method was used to compare 44 different oxPLs in ischemic and nonischemic myocardium. It was found that levels of these oxPLs varied, with hydroperoxides, aldehydes, and carboxylic acids the most abundant in both tissues, but there appeared to be greater increases in ischemic tissue of products derived from C20:4 and C22:6 containing PLs, where the level approximately doubled according to the peak area. However, a limitation of the study was that quantitative and statistical analyses were not carried out. The authors concluded that C22:6 lipids provide a protective barrier against oxidative damage, but further work would be required to demonstrate the benefit of these lipids in ischemia.
Increased levels of oxidized CL were detected in mitochondria from cardiac muscle after ischemia in the aged heart (62). Data-dependent acquisition was used, which is a nontargeted MS methodology: at each sampling point of the chromatogram, the most intense peak in the spectrum was selected for fragmentation (MS2) and the most intense signal in the MS2 spectrum was fragmented further (MS3). The MS data obtained were searched for masses of interest related to oxidized CL. A particularly interesting finding was that oxidation of CL appeared to occur in a nonstochastic manner, with the addition of three oxygen atoms apparently on a single acyl chain, rather than a mixture of oxidation products and sites. The implication is that ischemia injury may lead to an enzymatic oxidation of CL, which could be instrumental in initiating apoptosis; oxidation of CL is known to favor release of cytochrome c from mitochondria and decrease the activity of the electron transport chain complexes (91, 107). Nonrandom oxidation of CL has also been reported in other tissues and conditions (133). Thus, MS analysis can provide important information about cellular and biochemical processes.
Coronary and peripheral artery interventions are used to treat obstructive vascular diseases and acute coronary syndromes, but release of material from the plaque can cause distal complications such as embolisms. Ravandi et al. (98) used distal embolic protection filters to collect material released during these procedures and demonstrate the presence of oxidized PLs. Scanning for precursors of m/z 184 (phosphocholine headgroup) in the positive ion mode was used to identify PC-containing PLs in the extracted filter material, before quantitative analysis using SRM. The method was established using six commercially available oxidized lipids with 1,2-dinonanoyl-sn-glycero-3-phosphocholine as an internal standard, so the oxPLs could be quantified in terms of ng of material collected or as a % of the total oxidized PC analyzed. The filter material was enriched in oxidized lipids compared with unoxidized LDL, with PONPC as the most abundant component; around 75 ng was found to be present in filters for saphenous vein grafts, although peripheral and carotid interventions released less material (∼50 ng). This is important, as aldehyde and carboxylate-containing products have been found to have different effects on vascular tissues (7, 30, 38). The study also demonstrated the value of protection filters to reduce the amounts of oxidized PLs reaching the microvasculature (98).
Essentially the same MS approach, involving transitions from the parent molecular ion to m/z 184, also demonstrated that levels of hydroxy, hydroperoxy, and truncated PCs were increased in plasma from rats and patients suffering from chronic alcohol exposure and alcoholic liver disease. Analysis of the temporal appearance of these products suggested that rather than being related to initial oxidative stress occurring during ethanol metabolism, they were produced later during the critical transition to pathologic steatohepatitis, when hepatic inflammation occurs. As oxidatively truncated phospholipids are pro-inflammatory and pro-apoptotic, it was suggested that they were likely to contribute to adverse systemic effects of chronic ethanol exposure, as well as being markers of steatohepatitis (148).
Oxidized PEs were found to occur in bronchoaveolar lavage fluid from patients with lung diseases, using negative ion MS analysis (42). Precursor ion scanning was used as a discovery tool to find precursors of m/z 317.2 (KETE-containing lipids). The analysis of individual oxPEs was then carried out by monitoring transitions from parent ions to m/z 153.2 (12-KETE), 219.2 (15-HETE), or 179.2 (12-HETE), using higher collision energies to induce dissociation within the chain and provide information on positional isomers. Patients with cystic fibrosis had significantly higher levels of three oxidized plasmenyl phosphoethanolamines: 18:0p/15-KETE-PE, 18:1p/15-KETE-PE, and 16:0p/15-KETE-PE, at levels from approximately 0.05 to 3.5 ng/ml; although similar levels of the ester-linked compound 18:0a/15-KETE-PE were observed, the difference was not significant. The HETE- and KETE-containing PEs are mainly generated by the action of 15-lipoxygenase in macrophages, and they were shown to cause stimulation of PPARγ and upregulate CD36 expression in peritoneal macrophages (42). The increased levels of these oxidized lipids are thought to have a role in damping down the acute pulmonary inflammation characteristic of cystic fibrosis and may offer therapeutic potential for treating the disease.
The formation of oxidized CL and PS has also been observed in animal models of lung damage, such as that induced by hyperoxia, γ-radiation, or carbon nano-tubes, which can induce chronic pulmonary inflammation and fibrosis (5, 133, 135). Using shotgun lipidomics for initial identification followed by LC-MSMS for quantitative analysis, these conditions were found to result in much greater % increases in hydroperoxides and hydroxides of PS and CL compared with the other more abundant lipid classes PC and PE. This selective pattern of phospholipid peroxidation involving primarily CL and/or PS has also been reported in traumatic brain injury, and the observation that only certain polyunsaturated species within each of these classes became oxidized is in agreement with the study of CL oxidation in cardiac muscle (62). These studies concluded that mitochondrial lipid peroxidation pathways contributed to the chronic damage involved in inflammatory lung damage by triggering apoptotic pathways, and that mitochondrially targeted electron scavengers or heme-iron ligands might be able to protect against this damage (133).
Future Perspectives
A great deal of information currently available on the extensive range of individual oxidized phospholipids that occur both in vitro and in vivo has been provided by MS analysis. Interestingly, although products such as hydroperoxides and epoxides are often regarded as unstable, they have been detected in tissue samples from many acute and chronic inflammation conditions, as have the more stable truncated phospholipids containing reactive aldehyde or ketone moieties that can also form adducts to proteins, and a range of other oxidized products with known or potential biological activity. Using accurate mass or fragmentation patterns, distinct oxidized molecular species and isomeric compounds have been identified. In fact, the major problem with nontargeted MS analysis is the amount of data available, exemplified by the size of a typical data file for a 1 h LC-MSMS run of 0.5 GB, and the consequent complexity of its analysis. For example, recent lipidomic studies of LDL have identified between 150 and 275 native molecular lipid species (13, 100), a study of mouse plasma using SFC identified 461 lipids (145), many lipids have been comprehensively profiled in brown adipose tissues (47), and other reports have identified even higher numbers (11); the likely number of different oxidized forms is considerably higher than this, and many different oxidized phospholipids have been reported in diseased tissues. The manual identification of such large numbers of species becomes an impossible task, and other approaches are required to make the data manageable.
One approach, as described earlier, is the use of targeted methods to find specific relevant modified lipids. An alternative approach is the use of software to cross-search the MS data against databases of lipid masses and fragmentation patterns for identifications. The most established open access database for lipidomics is LIPIDMAPS (
An issue that will be important for the future is the availability of standards. In contrast to the large number of available isotope-labeled or unlabelled nonesterified lipids, standards for oxidized phospholipids are fewer and often prohibitively expensive, owing to the greater synthetic difficulty. Relative quantification, involving comparison of lipid profiles between samples or ratioing of oxidized products versus native precursors, can be carried out without labeled standards (101). However, absolute quantification of oxidized lipid levels requires standards of all analytes of interest, as the ionization efficiency varies with chain length and phospholipid class. Quantification by comparison with a small number of standards that may not have directly comparable MS behavior has limitations; especially in discovery applications, absolute quantification continues to be a challenge.
Unlike protein aldehydes (generated from direct oxidation or short-chain lipid adducts, such as HNE), there are very few reported methods for the enrichment of oxidized phospholipids, which is especially surprising given their generally low abundance. Of the few there are, the most effective appear to be nanoparticle-based enrichment of acidic products generated either directly from the oxidation or via reaction of lipid aldehydes with an acidic aniline based reagent (4-aminobenzoic acid) using either ZrO2 or surface functionalized Fe3O4 core-shell superparamagnetic nanoparticles (123), or a bifunctional thiol hydrazide that can be captured on GNPs for isolation and then the aldehyde is released using hydroxylamine (41). The former have the advantage that they can be directly analyzed by adding the particles to the MALDI matrix.
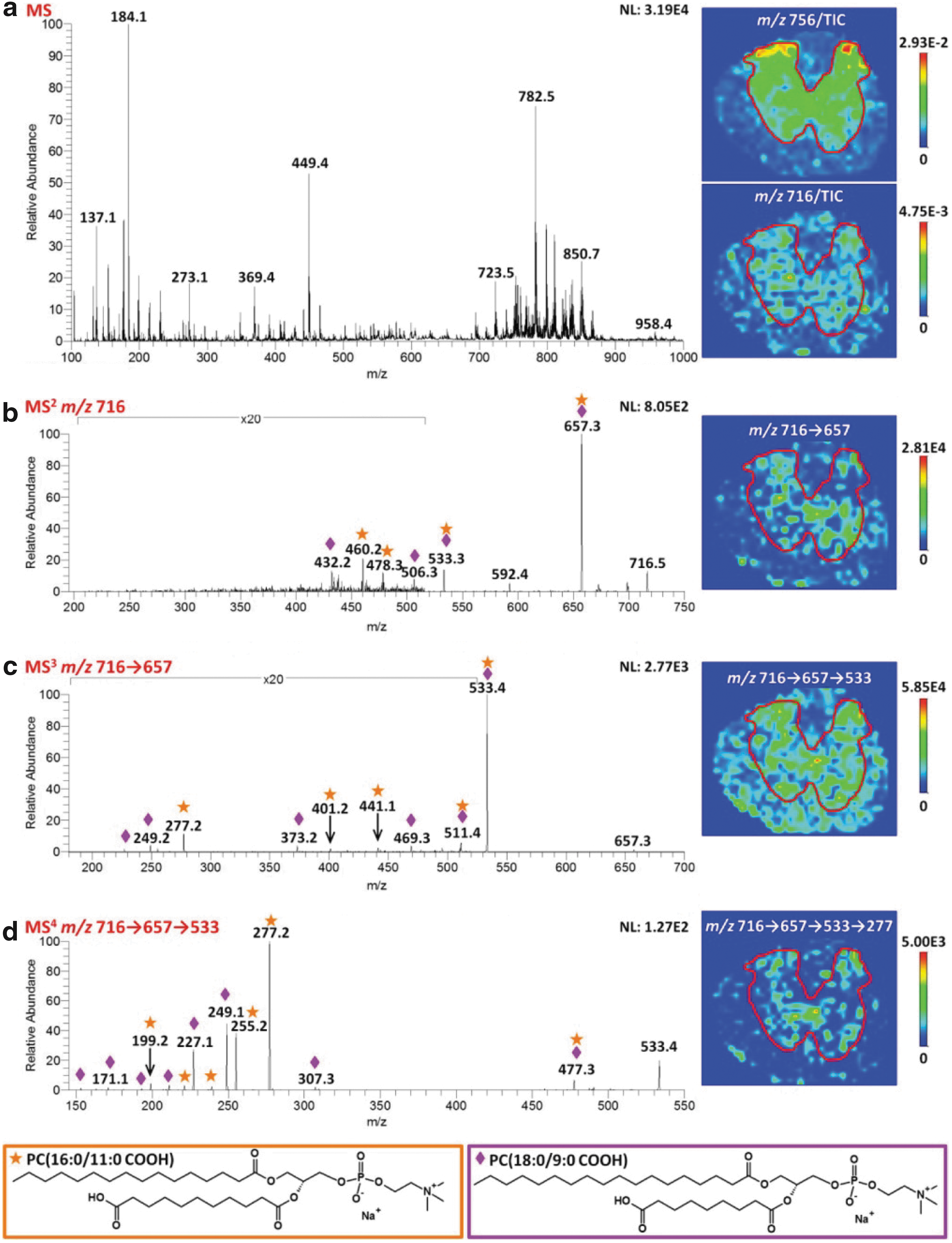
An area that is likely to grow considerably in the next few years is MS imaging of oxidized lipids. MS imaging involves collection MS or MRM data at a number of raster points across a sample, and reconstructing an abundance map from this using ion intensities (36, 84), and its potential for phospholipids has been recently reviewed (113). The most commonly used method to date is MALDI, although a range of newer methods that operate at atmospheric pressure, such as desorption ESI (24), laser ablation electrospray ionization (LAESI) (86), laserspray ionization (LSI) (44), and liquid extraction surface analysis (LESA) (10), which generally have limiting resolutions between 20 and 100 μm, and higher-resolution methods such as secondary-ion mass spectrometry (SIMS, sub-micron resolution) (6) are showing promise as alternative methods. The use of ToF-ToF instruments, and especially the coupling of imaging methods to other high-resolution MS instruments where ion analysis and fragmentation are fully decoupled from the ionization method, along with the application of ion mobility MS are revolutionizing the capabilities of this methodology. MALDI imaging appears to work particularly well for easily ionized lipids (PC, PE) as well as for other metabolites or peptides (88), and studies on the lipid content of a number of animal tissues have been reported, especially in the brain and liver, which have a high lipid content; this has been comprehensively reviewed recently (35, 36). As yet, very few studies in which oxidized lipids were detected have been reported (114), but Stutts et al. have used sophisticated MSn methodology to image truncated carboxylate-containing PCs in rat spinal cord and show they were mainly localized to the gray matter (Fig. 11) (124), and the analysis of oxidized lipids in traumatic brain injury has been recently reviewed (114). This technology is an area of considerable interest and value for understanding the localization of oxidized phospholipids within diseased and inflamed tissue, although currently spatial resolution of the image is limited by the laser spot diameter and other factors to 10–50 μm.
In conclusion, oxidative lipidomics is an exciting and biomedically important area that depends largely on MS technology, and in which we can expect to see significant advances in the near future as the technology and supporting methodological refinements are developed further.