
Abstract
Introduction
O
The physiological role of the redox active CXXC motif of human branched-chain aminotransferase (hBCAT) remains unclear. This report demonstrated a novel functional role of hBCAT in redox protein folding. Moreover, this is the first study which reports that hBCATm can bind to protein disulfide isomerase (PDI) in the human brain and that PDI is co-localized with hBCATm in mitochondria. Overall, this has key implications, as both proteins are modified in Alzheimer's disease (AD) brains, which could impact the regulation of protein folding by possibly contributing to the pathogenesis of AD. Understanding the role of dys-regulated redox mechanisms will assist in the identification of therapeutic targets for the treatment of neurodegeneration.
The human branched-chain aminotransferase proteins (hBCATs) (EC 2.6.1.42) catalyze the reversible transamination of the α-amino group of the branched-chain amino acids to α-ketoglutarate, forming their respective branched chain α-keto acids and glutamate (30, 32). The branched chain α-keto acids are further metabolized through irreversible decarboxylation that is catalyzed by the branched chain keto acid dehydrogenase complex (BCKDH) (25). Although other isoforms exist, the cytosolic (hBCATc) and mitochondrial (hBCATm) form predominate, with hBCATm expressed in most tissues and hBCATc primarily expressed in the brain and peripheral nervous system (22, 29, 30, 57). The hBCAT proteins operate as homodimers with a subunit molecular mass of 41.73 and 43.40 kDa for hBCATm and hBCATc, respectively (20). Transamination by the BCAT proteins in the brain contributes to more than 30% of de novo brain glutamate (30). In human and rat brain, hBCATc is specific to glutamatergic and GABAergic neurons, where expression is predominantly in the soma and proximal dendrites, in keeping with its role in contributing to the storage and metabolic pools of glutamate, which is, in turn, decarboxylated in GABAergic neurons to form GABA (22, 57). The mitochondrial isoform is localized specifically to the human brain vasculature with localized neuronal staining, where a role in brain glutamate metabolism has been suggested (28); whereas it is located in astrocytes of rat models (8, 9, 12).
Unique to the mammalian BCAT proteins is a conserved CXXC motif, which is ∼10 Å from the active site (23, 72). X-ray crystallography and biochemical studies of overexpressed hBCAT have shown that these thiol groups confer redox-linked regulation to the hBCAT proteins and that the active and inactive form results from the reduced and oxidized CXXC motif, respectively (18, 23, 72). In response to oxidation, the thiol at the N-terminal position acts as a redox sensor, whereas the C-terminal residue permits reversible regulation through the formation of a disulfide bond via a sulphenic acid intermediate, resulting in a loss of hBCAT activity (16, 17). X-ray crystallography studies and kinetic analysis demonstrated that the predominant effect of oxidation was on the second half reaction rather than on the first half reaction, where disruption of the CXXC center results in altered substrate orientation and an unprotonated pyridoxal mono-phosphate amino group, thus rendering the enzyme catalytically inactive (73). Both isoforms were differentially inactivated through S-nitrosation; however, prolonged exposure to S-nitrosoglutathione (GSNO) resulted in a shift from S-nitrosation to S-glutathionylation, suggesting that S-glutathionylation plays a protective role in preserving hBCAT function during periods of cellular stress (13). Physiologically, the reduced state of the CXXC motif of the hBCAT proteins is essential for the binding of hBCATm to the E1 subunit of the BCKDH complex, facilitating substrate channeling of the α-keto acids (33). In a separate study, using extracts from neuronal cells, hBCATc was shown to have specific peroxide-related redox associations with several proteins known to be involved in protein cell signaling, indicating a novel role for hBCATc in cellular redox control (15). However, neither the cellular mechanisms that govern this redox sensitivity nor their redox substrates are completely understood.
In these studies, we have found that the hBCAT proteins have dithiol-disulfide isomerase activity that is regulated through S-glutathionylation. We provide in-vitro, ex-vivo, and in-vivo evidence that hBCAT associates with PDI, the interaction of which is altered through oxidative and S-nitrosative stress-induced conditions, where we propose that hBCAT may have a novel chaperone role in mediating protein folding. The localization of these proteins to neuronal and endothelial cells of the vasculature of the human brain along with their co-immunoprecipitation demonstrates that the proposed chaperone role may have physiological relevance. These findings provide the first evidence that the hBCAT proteins have oxidoreductase properties similar to PDI and that these proteins may act as neuro-protectors during the cellular stress response, which may become compromised by contributing to protein misfolding, a key feature of neurodegenerative conditions such as AD.
Results
Endogenous PDI-like activity of the hBCAT proteins in altered redox environments
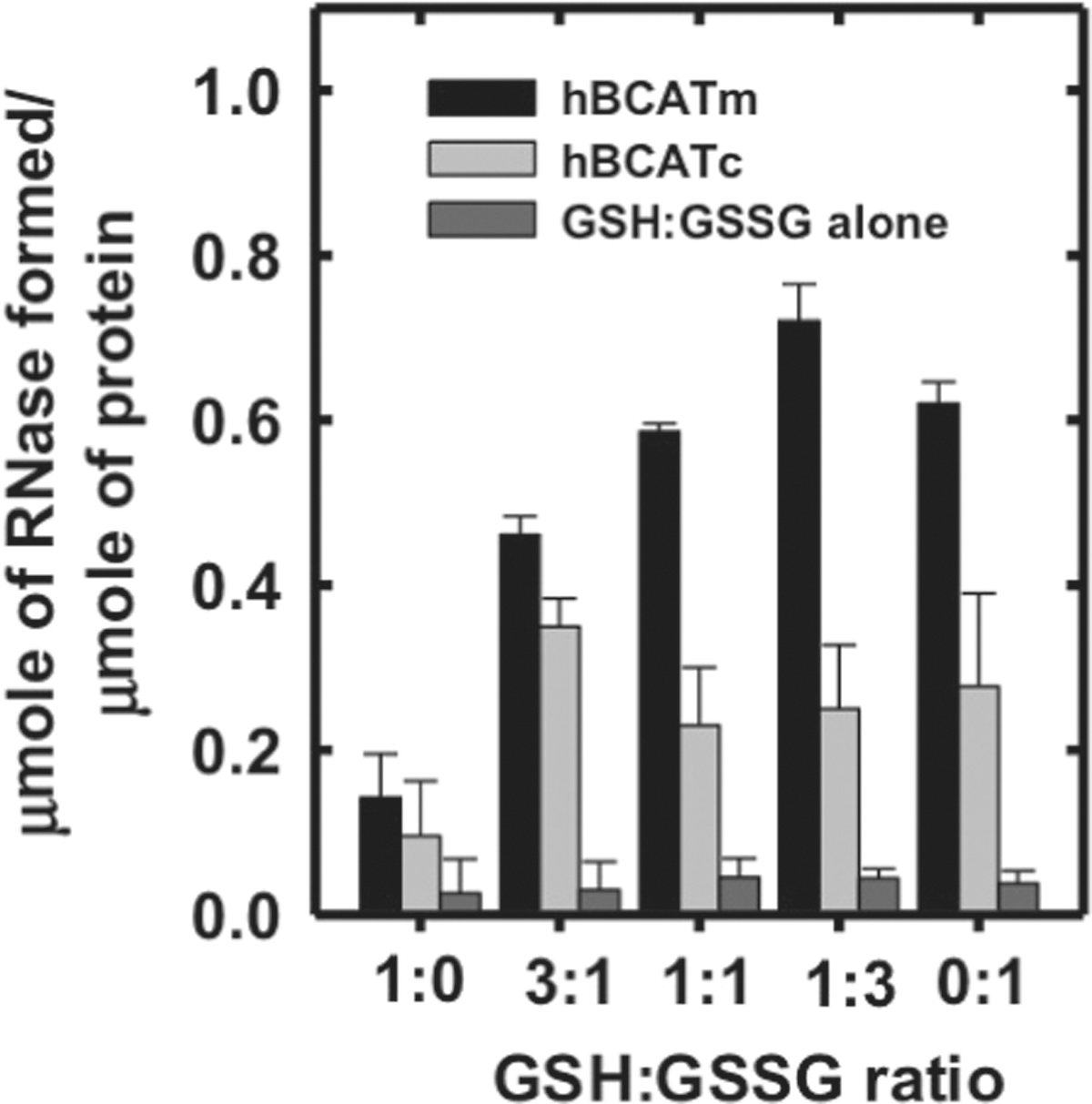
Both hBCATm and hBCATc catalyzed the refolding of reduced and denatured RNase (rdRNase) through the introduction of disulfide bonds at a rate significantly greater than air oxidation alone, with activity at 55%–60% relative to PDI (Fig. 1A), confirming their oxidase activity. Ovalbumin, which does not have a redox-active CXXC motif but contains free thiols and disulfide bonds, did not exhibit measureable activity over 24 h. The refolding of rdRNase by the hBCAT proteins was found to be both time- and concentration dependent and saturated at concentrations of 12 μM (Fig. 1B, C, respectively). Since protein folding is inherently influenced by the redox environment (31), we assessed the influence of various glutathione (GSH) buffers on hBCAT oxidase activity. Although GSH buffers alone offer partial refolding of rdRNase, the oxidase activity of both hBCAT isoforms was increased two-fold at a 3:1 ratio of GSH/Glutathione disulphide (GSSG) compared with GSH alone (Fig. 2). However, as the environment became more oxidizing, the oxidase activity of hBCATm continued to increase relative to hBCATc, which itself remained unchanged at these concentrations.
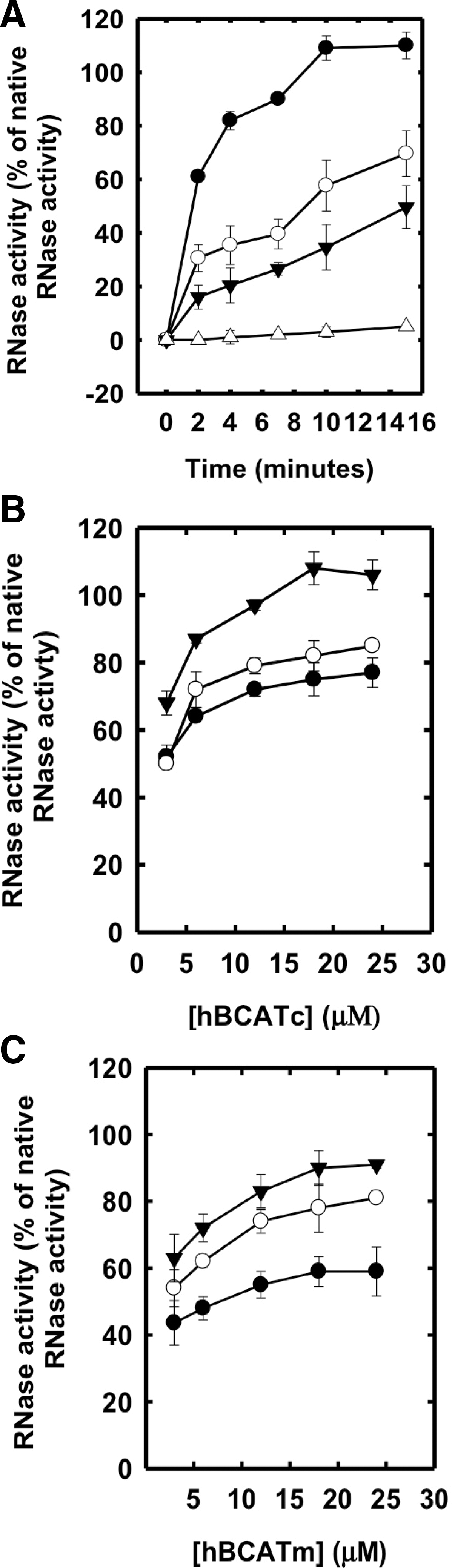
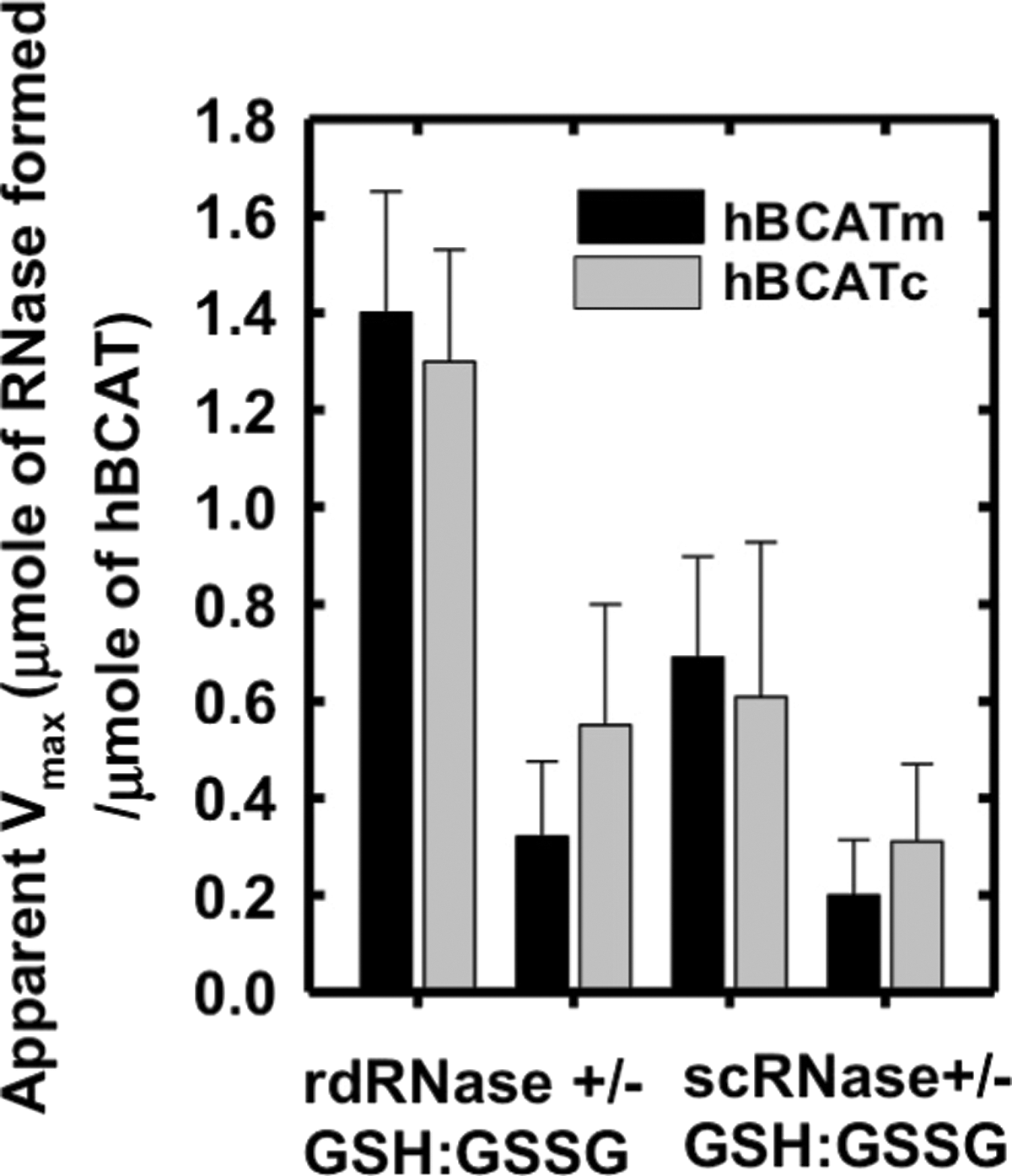
In the absence of a GSH buffer, the apparent Vmax was 4.4- and 2.4-fold less for hBCATm and hBCATc, respectively (Fig. 3). To investigate the ability of hBCAT proteins to correctly align mis-paired disulfides, we used scrambled RNase (scRNase) as a substrate, which is rdRNase with random disulfide bond formation. Titration of scRNase with 5,5′-dithiobis(2-nitrobenzoic acid) (DTNB) before and after oxidation resulted in a loss of thiols to 0.04 μmoles/μmole of protein with <1% RNase activity. Although in the presence of GSH/GSSG the apparent Vmax for refolding of scRNase was 3.4- and 2.4-fold higher relative to the reaction without the redox buffer, the dithiol isomerase activity was ∼50% less than that reported for their oxidase activity with rdRNase (Fig. 3). These results demonstrate that the hBCAT proteins have oxidase and thiol isomerase activity, the kinetics of which is dependent on the redox environment.
Target site of dithiol-disulfide exchange
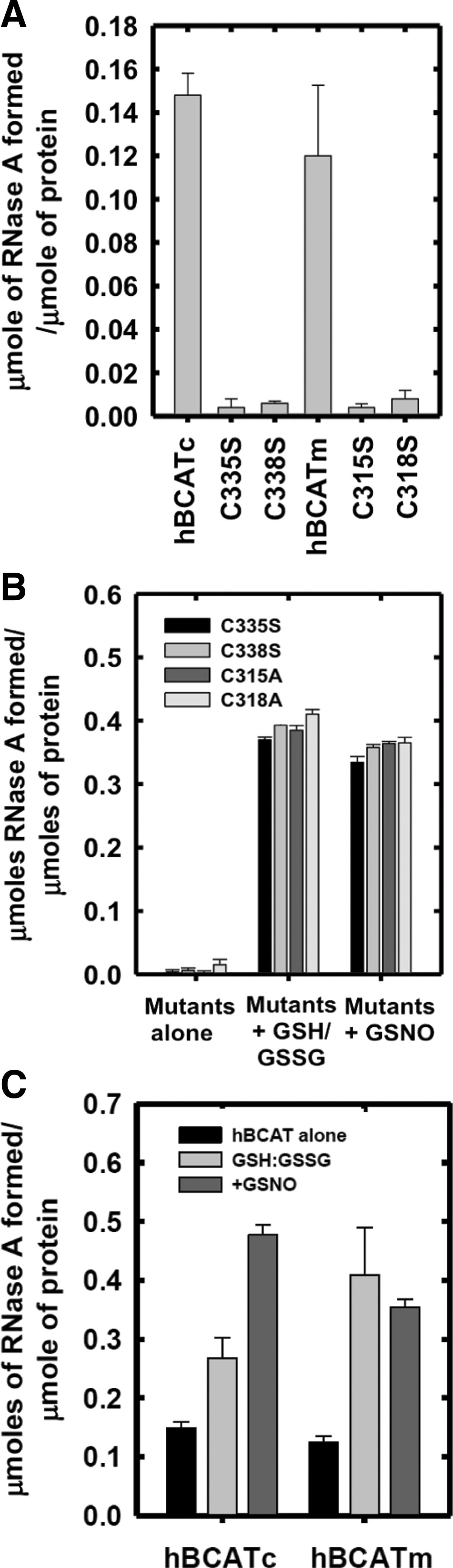
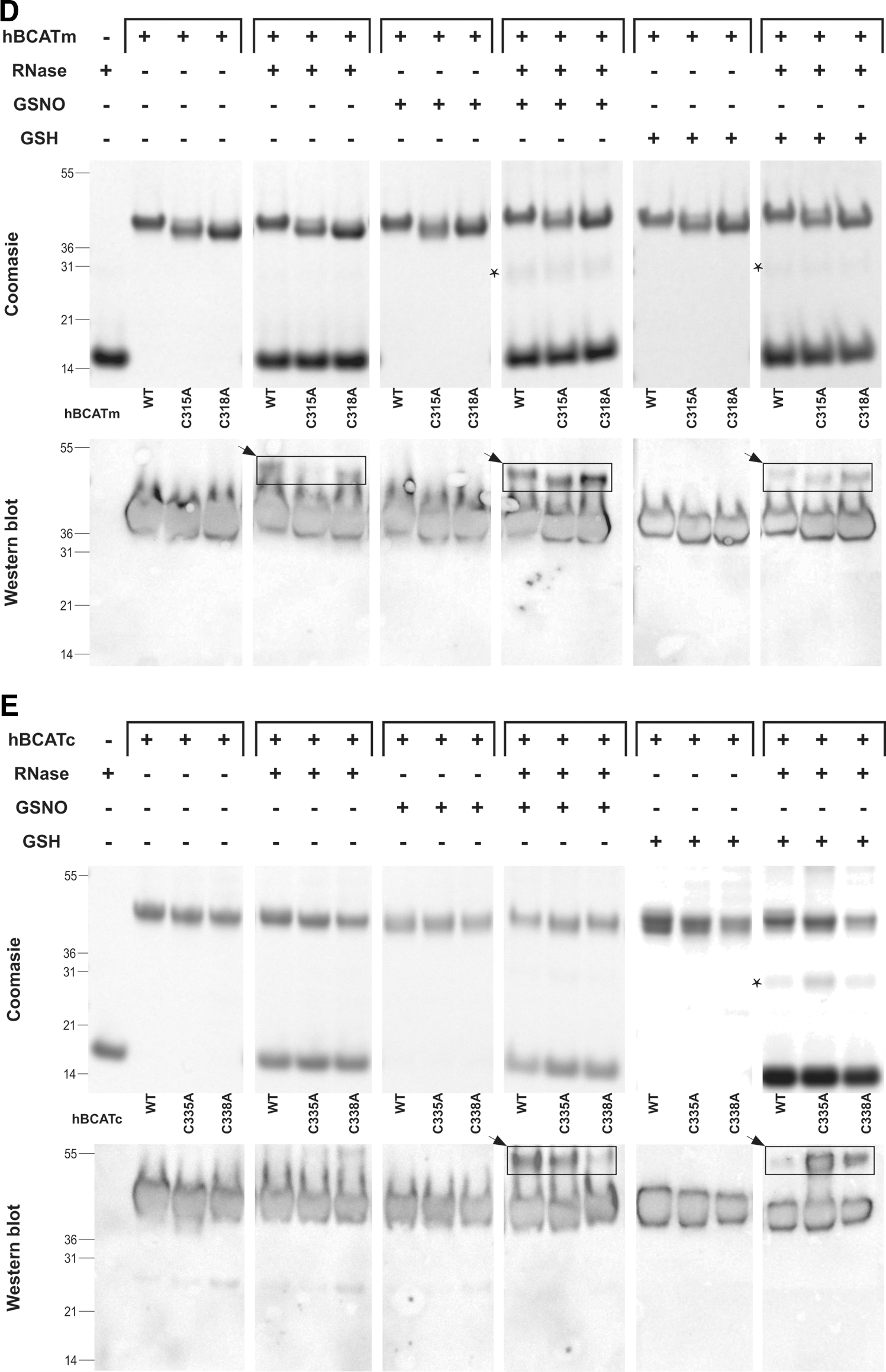
To determine the target site of these refolding mechanisms, active-site cysteine mutants of the hBCAT proteins were assessed for their ability to catalyze the refolding of rdRNase. At 24 h, the oxidase activity of cysteine active-site mutants was negligible (Fig. 4A), but was recovered by the addition of a GSH buffer or GSNO, similar to native hBCAT under these conditions (Fig. 4B, C, respectively). Western blot analysis confirmed that native hBCAT associates and forms a complex with RNase (estimated M.Wt.=53 kDa), which is not evident in the N-terminal mutant proteins unless S-glutathionylated (Fig. 4D, E, respectively, arrow). Also observed were multimers when hBCATc was incubated with GSNO or the GSH buffer (Supplementary Fig. S1; Supplementary Data are available online at
Association of the hBCAT proteins with human PDI
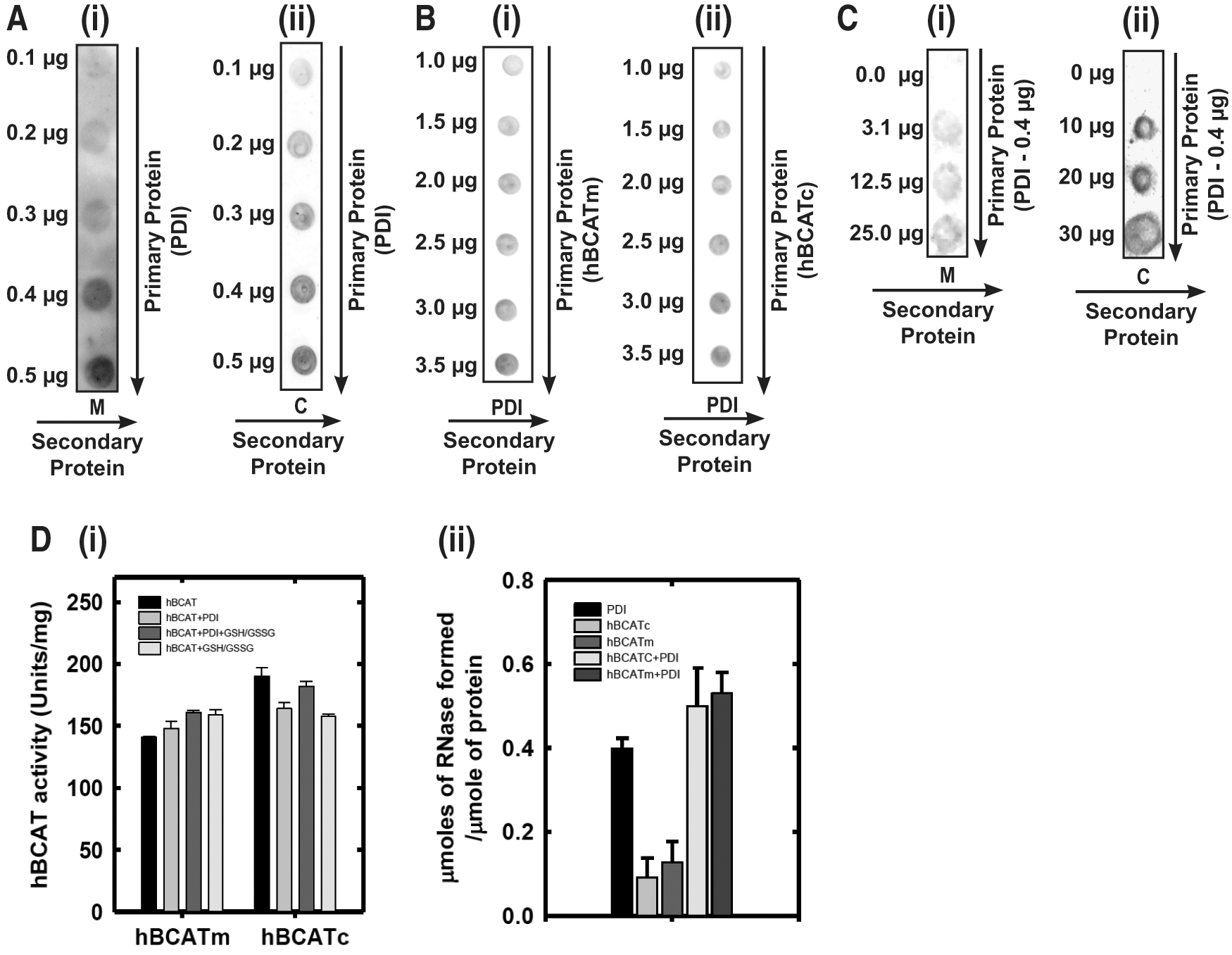
Far Western blot analysis demonstrated that there is a dose-dependent direct binding of hBCATm and hBCATc with hPDI (Fig. 5A [i, ii]). Binding is also demonstrated using a reverse dot blot for PDI with hBCATm and hBCATc, respectively (Fig. 5B [i, ii], varying concentrations of PDI and Fig. 5C, PDI constant at 7 nmol). Although PDI did not significantly influence hBCAT transaminase activity (Fig. 5D [i]), incubation of hBCAT with PDI resulted in an increase in oxidase activity (Fig. 5D [ii]). Since the initial phase of the reaction occurs earlier relative to the proteins in their independent reactions, we conclude that hBCATm acts in a synergistic manner along with PDI in the initial stages of the refolding reaction but has less influence during the linear phase (Supplementary Fig. S2). Thus, these proteins can associate with each other in a dose-dependent manner, and protein folding is influenced by this binding.
Co-localization of hBCATm with PDI and Mia40 in IMR-32 neuronal cells using confocal microscopy and electron microscopy
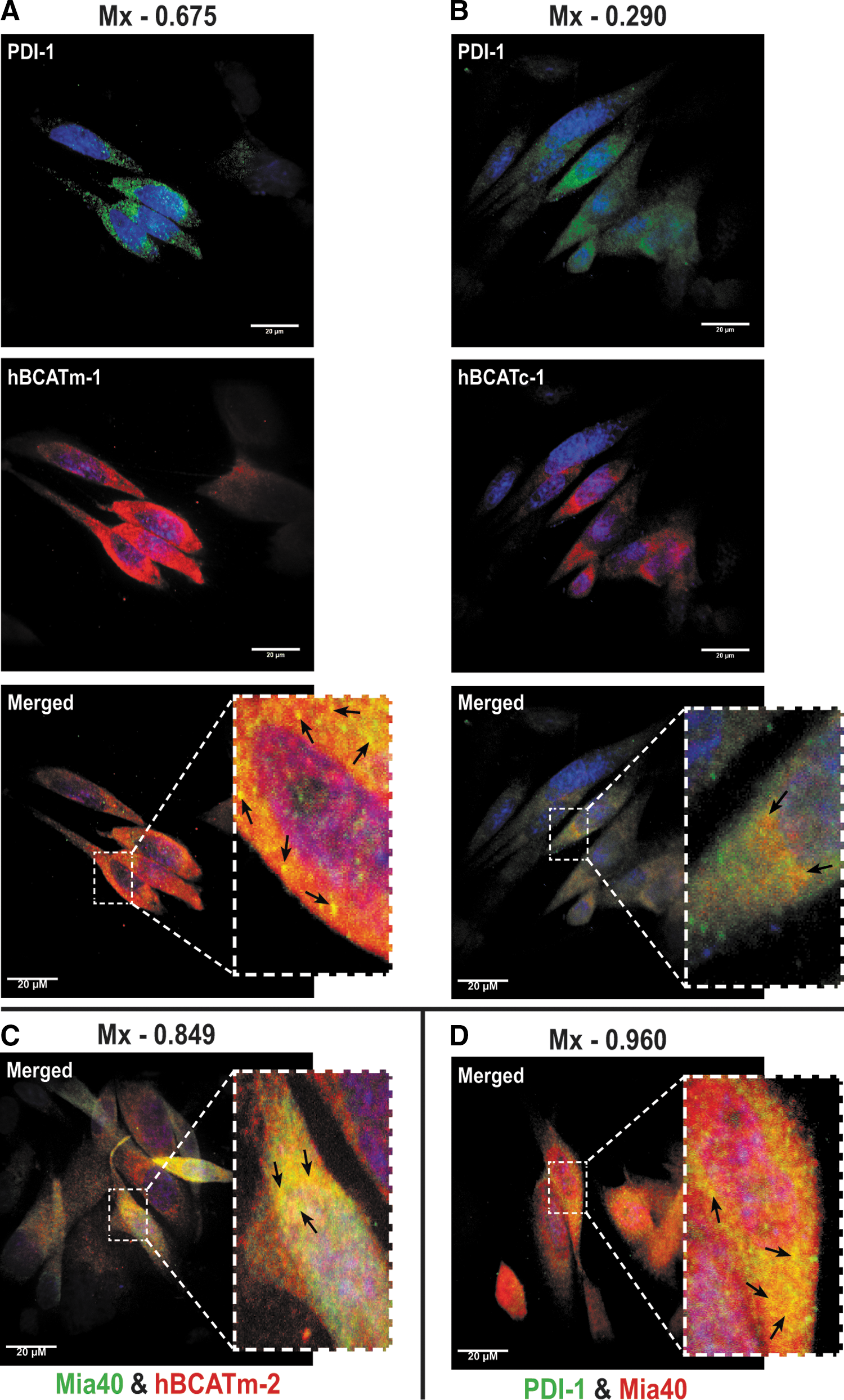
Confocal microscopy established that both isoforms differentially co-localize with PDI, suggesting that they occupy the same space within the cell and are interacting (Fig. 6A, B, merged image). The co-localization of hBCATm/hBCATc (red channel) and PDI (green channel) is shown as yellow (merged image) (Fig. 6A, B, respectively). Mander's correlation coefficients (Mx and My) were derived using Volocity (Perkin-Elmer) from an average of 15 individual cell images (6). Mx signifies the correlation of hBCATm/hBCATc co-localizing with PDI, and the My value reflects the correlation of overlap between PDI and hBCAT. This method quantifies the signal intensity and can be measured in a range of 0–1, with 0 indicating no co-localization and 1.0 indicating complete co-localization. The average Mx values for hBCATm with PDI were 0.675 (Fig. 6A) and 0.648 (Supplementary Fig. S3), respectively, validated using antibodies specific for PDI from two independent manufacturers, demonstrating a strong co-localization between hBCATm and PDI; whereas the My value, a measure of PDI co-localizing with hBCATm, was recorded at 0.31. The co-localization of hBCATc with PDI was not as strong with an Mx value of 0.29 recorded (Fig. 6B, merged image). In addition, hBCATm showed co-localization with Mia40 (Mx=0.849, Fig. 6C), a mitochondrial-specific redox protein that also co-localized with PDI with an Mx value of 0.96, confirming that this co-localization is specific to the mitochondria (Fig. 6D). The My correlation for PDI with Mia40 was 0.23. Together, these data indicate that in the mitochondria, the co-localization of hBCATm with PDI is strongly correlated but relative to the whole cell, co-localization of PDI with these mitochondrial proteins (that will account for ER expression of PDI); the signal is weak, illustrating that the co-localization is clearly specific to the mitochondria.
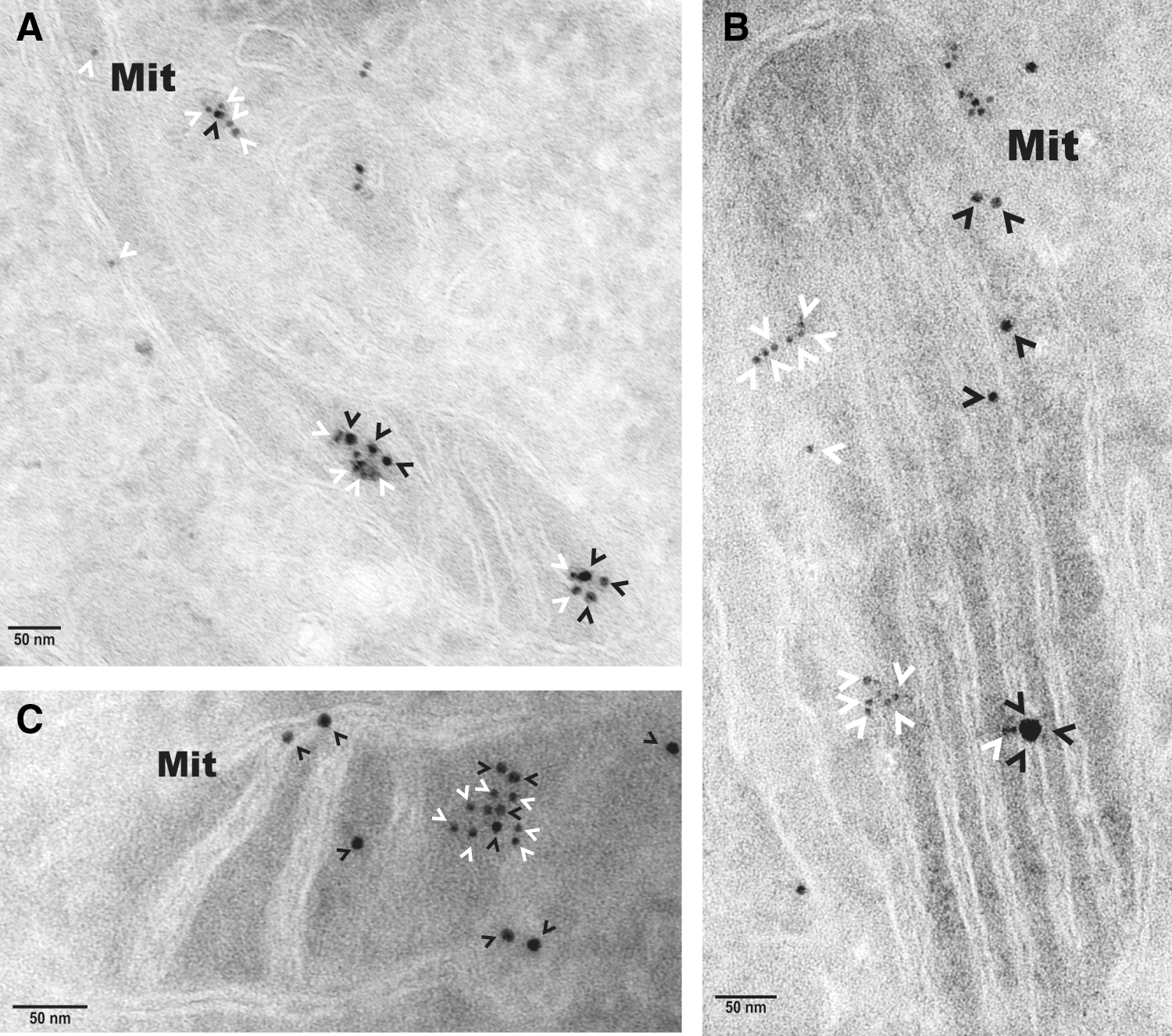
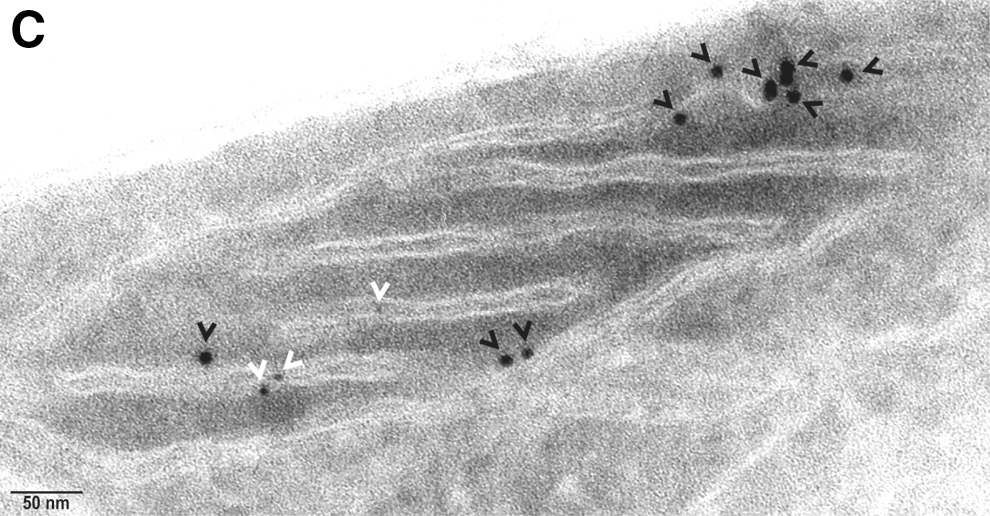
Electron microscopy independently validated that hBCATm can co-localize with PDI in the mitochondria as determined by calculating distances between cellular proteins, where a distance of approximately 50 nm apart is considered co-localization, which factors the distance of the antibodies and gold particles (64). IMR-32 cells showed a predicted distribution of hBCATm and Mia40 within the mitochondria (distance apart=9.46–11.59 nm), suggesting co-localization (Fig. 7A). Validation that PDI was also detected in the mitochondria was determined using Mia40 as a mitochondrial control protein (Fig. 7B). In a separate section, PDI was observed in the mitochondria and in close proximity with hBCATm (distance apart 11.16–15.85 nm), validating our confocal microscopy data that indicated these two proteins were occupying the same space, suggesting an interaction (Fig. 7C). It is also important to note the detection of hBCATm in the intermembrane space (Fig. 7C). These studies were validated using antibodies from several suppliers (Supplementary Fig. S4), demonstrating that PDI is found in the mitochondria of neuronal cell cultures and is co-localizing with hBCATm.
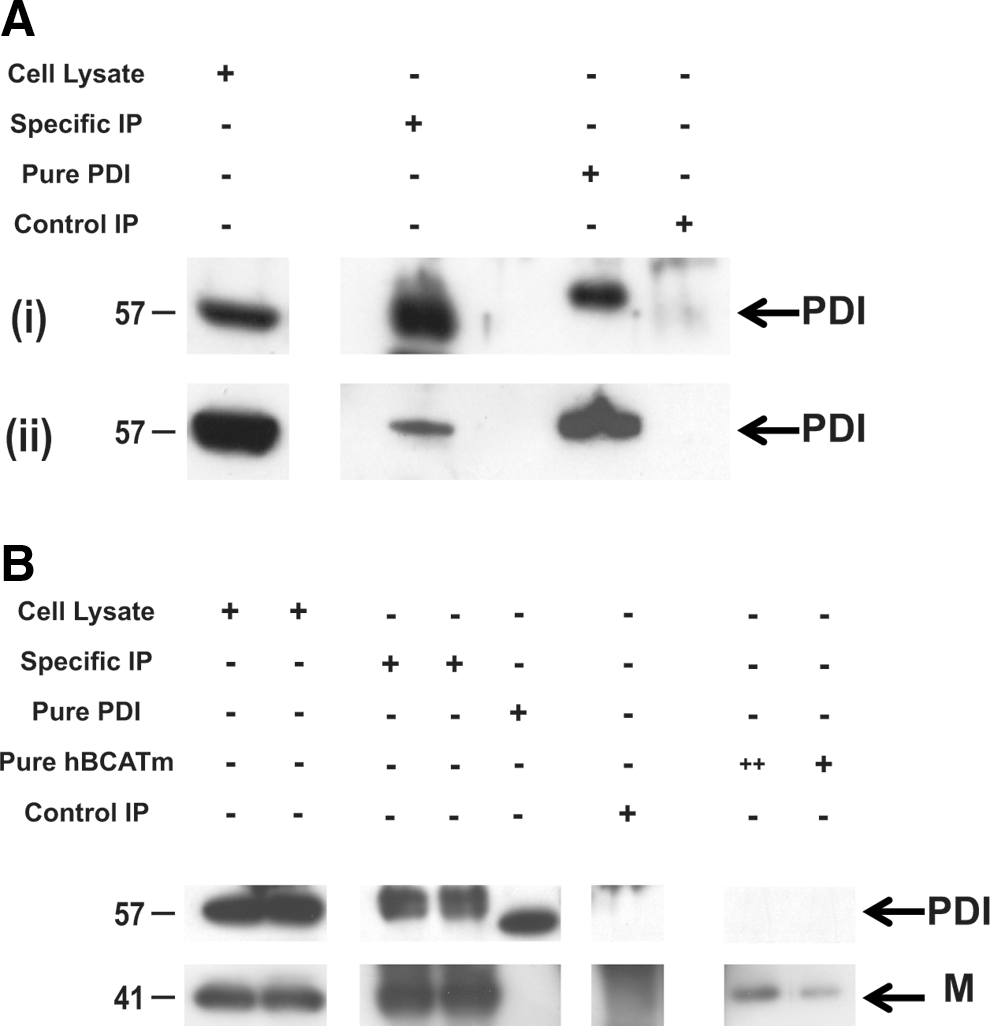
Co-immunoprecipitation of hBCATm and PDI from IMR-32 cells
Direct binding of hBCATm with PDI was demonstrated using immunoprecipitation (IP) analysis, where antibodies specific for hBCATm and HIS-tagged hBCATm isolated PDI from neuronal cell lysates transfected with hBCATm-pDEST™26 Gateway® destination vector (HIS-tagged) and homogenates from human brain (Fig. 8). The expected 57 kDa PDI protein was precipitated with anti-hBCATm and detected using Western blot analysis using anti-PDI (Fig. 8A [i]). This band corresponded to the bands observed in whole cell lysates and pure overexpressed hPDI protein. The absence of a band in the control IP samples confirms that the proteins eluted are specific to anti-hBCATm. Moreover, using the same conditions, PDI was co-immunoprecipitated from human brain samples using anti-hBCATm (Fig. 8A [ii]). To further validate the specificity of this interaction, neuronal cells were transfected with HIS-tagged hBCATm and shown to specifically interact with PDI, where the concentration of hBCATm in the purified complex is ∼31 nM with 16 nM of PDI (as estimated from pure hBCATm and PDI, respectively), indicating a 2:1 ratio of hBCATm to PDI (Fig. 8B). These studies demonstrate that PDI was co-purified with hBCATm, suggesting that hBCATm directly binds to PDI in neuronal cells and, more importantly, in the human brain.
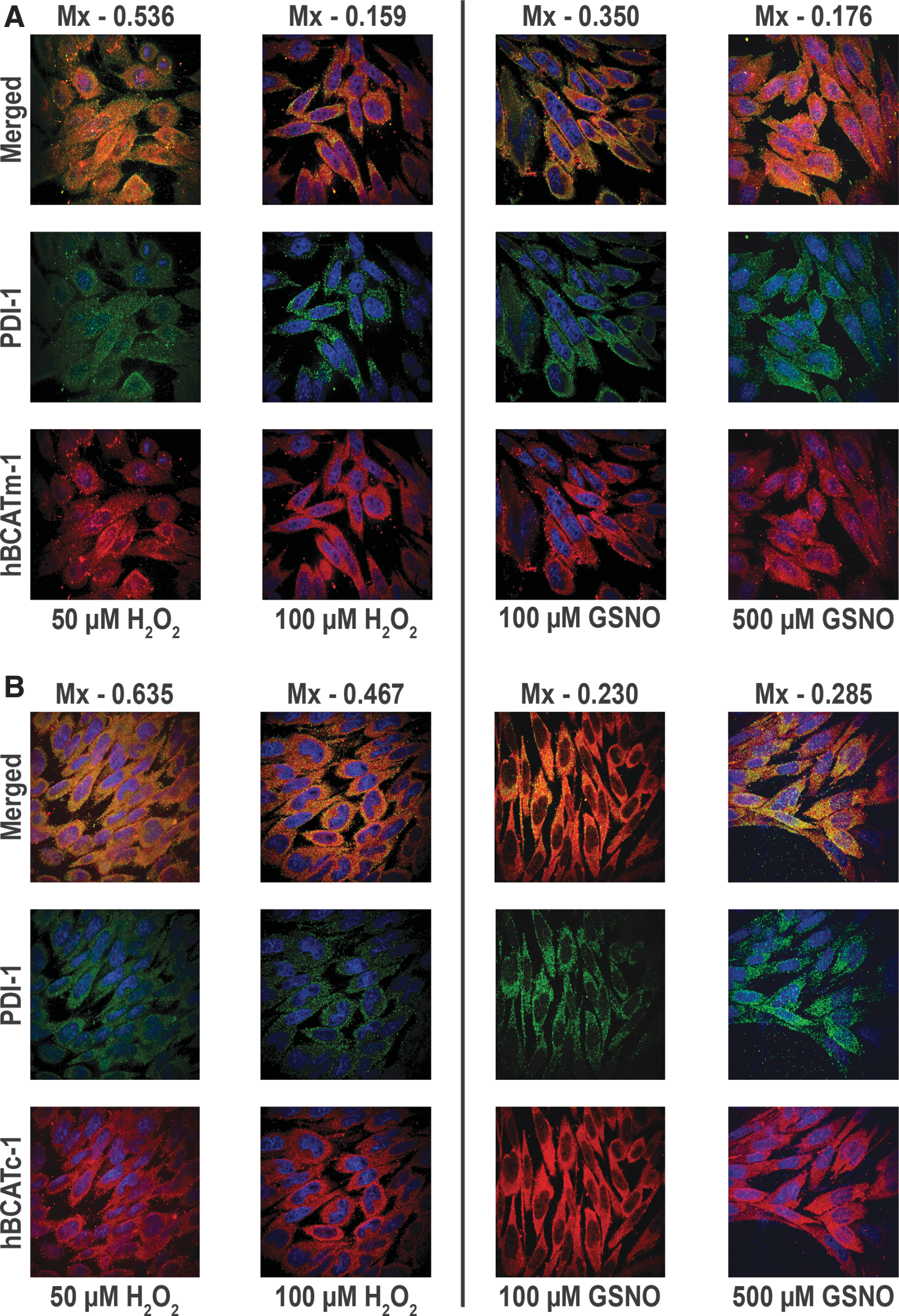
Co-localization of hBCATm with PDI in IMR-32 neuronal cells is reduced under increased oxidative stress
The co-localization of hBCATm with PDI decreased (Mx=0.675 to 0.159) when the cells were incubated with increasing concentrations of hydrogen peroxide or GSNO, respectively (Fig. 9A). Incubation of IMR-32 cells with GSNO did not impact the association between hBCATc and PDI; however, with hydrogen peroxide, the Mx value of hBCATc with PDI increased from 0.29 to 0.635 (Fig. 9B). Electron microscopy also shows that hBCATm is no longer observed in close proximity to PDI when the cells are treated with 100 μM of hydrogen peroxide (Fig. 9C). The increased association of hBCATm with the mitochondrial membrane under these conditions should be noted. These studies taken together provide evidence that hBCAT can associate with other redox-sensitive proteins such as PDI and that these associations are altered when the cells are exposed to increased oxidative stress.
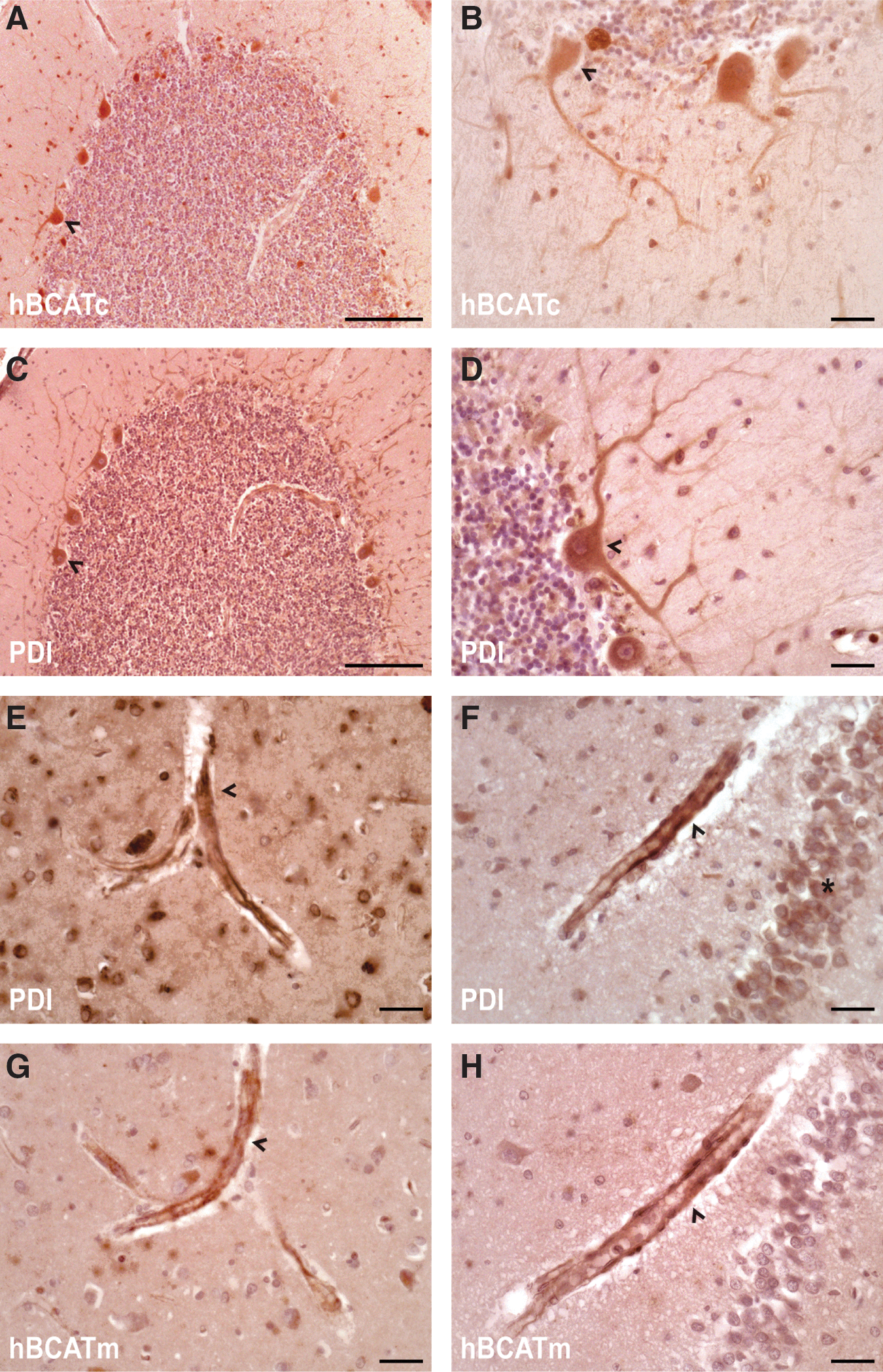
Expression of hBCAT proteins and PDI in the human brain
To ascertain whether the hBCAT proteins were expressed in the same human brain, cells as PDI serial sections from the temporal and cerebellum of human brains were analyzed using antibodies that were specific for each respective protein. In agreement with previous studies (28), hBCATc is neuronal specific (Fig. 10A, B) and here using serial sectioning, we show that PDI is expressed in the same neuronal cells with a similar pattern of distribution (Fig. 10C, D). Interestingly, strong staining of PDI was also indicated in the endothelial cells of the vasculature, where hBCATm is predominantly expressed, in addition to some neuronal staining (Fig. 10E, F and G, H, respectively). These studies confirm that PDI is expressed in the same brain cells as the hBCAT isoforms and along with the co-immunoprecipitation studies, indicate that these proteins have the potential to interact in vivo.
Discussion
Accumulation of mis-folded proteins is the hallmark of several neurodegenerative conditions, including Alzheimer's, Parkinson's, Huntington's, and prion diseases (9, 44, 71). However, the mechanisms that underpin this are not clearly defined. Similar to PDI and other oxidoreductases, the mammalian hBCAT proteins have a conserved CXXC motif, where the thiols of the CXXC motif can recycle between an intra-molecular disulfide bond (inactive) and the reduced dithiol (active) (61, 73). The present study demonstrates that hBCAT proteins catalyze the insertion of disulfide bonds into rdRNase-releasing active RNase, with the active cysteines playing a central role in this mechanism. Several other CXXC redox-sensitive proteins such as thioredoxin and platelet integrin alpha IIbbeta 3 have also been reported to have endogenous thiol disulfide isomerase activity (37, 40, 48). Inactivity reported for the N-terminal mutant proteins can be explained by their inability to form an intermediate with RNase, suggesting that the N-terminal cysteine is essential for the first step in forming a mixed disulfide with RNase. The N-terminal cysteine residues of both isoforms are central to their transaminase activity, as mutation of the reactive cysteine results in a complete loss in hBCAT activity (16). A number of the PDI family members, namely, ERp27, ERp28, PDILT, and TMX2, are missing in the surface exposed to N-terminal cysteine and similar to our N-terminal mutants, lack thiol disulfide isomerase activity, demonstrating the importance of this residue for catalysis (21, 65).
Even though the C-terminal mutant can potentially take part in the first step of the reaction, this intermediate may get trapped, as the reaction cannot subsequently progress to complete oxidation, as evidenced by the inability of these mutants to form active RNase, suggesting that this residue is required for substrate escape. This concept has previously been described for other systems, where stable intermediates result when the C-terminal residue is absent, such as that reported for ERp44, which results in the retention of these proteins in the ER, impacting protein function (3, 4). However, here, when S-glutathionylated, the single cysteine mutants have oxidase activity and form intermediates that are comparable to native proteins. This shows that either cysteine residue when forming a mixed disulfide with GSH can catalyze disulfide bond formation and implies that reactive thiols in other proteins subject to S-glutathionylation may also participate in oxidase activity, even if the CXXC motif is not present. Single active-site thiol disulfide exchange has also been explored and found to have effective activity for single mutants of the bacterial isomerase equivalent DsbA (69).
The increased oxidase activity of hBCATm relative to hBCATc reported as the environment became less reducing is consistent with their measured Em values (−310 and −225 mV, for hBCATc and hBCATm, respectively) (14). This indicates that while both proteins are reducing in nature, hBCATm is the less so of the two, and has a similar Em value to those reported for the human mitochondrial isoforms of the repair enzymes, thioredoxin and glutaredoxin (52, 66, 67). Taken together, we propose that hBCAT oxidoreductase activity is initiated through the S-glutathionylation of the hBCAT protein at the N-terminal cysteine position. Under nucleophilic attack, the reactive thiolate anion of RNase forms an inter-disulfide bond with S-glutathionylated hBCAT. Along with the C-terminal cysteine, the second thiol of RNase forms a disulfide bond, releasing RNase (S-S). Without the thiol at the C-terminal position, the mixed disulfide intermediate cannot be released and remains fixed to the N-terminal residue. Although S-glutathionylation is clearly involved in maximizing oxidase turnover, it is likely that other chaperones are involved in this mechanism in cells. A plausible candidate is human Ero1, a flavin adenine dinucleotide-binding oxidase, which has been proposed to act in part to provide oxidizing equivalents to PDI (24, 54). However, recent studies using knock-out Ero-1 have shown little physiological consequence, leading to debate as to whether Ero-1 is the primary donor of oxidizing equivalents or whether other mechanisms exist that substitute for its absence in these models (74). Lappi and Ruddock have recently revaluated the role of GSH buffers in this system and strongly advocate their physiological role in protein folding, in particular calling for further investigations into the physiological role of GSSG in the ER (38). The pathways for dithiol-disulfide exchange are complex and have not been entirely elucidated. Single thioredoxin-like domains of PDI have redox activity but poor isomerase activity, suggesting that a second CXXC motif is required for maximal dithiol-disulfide isomerase activity (19). This is possibly the case seen here for hBCAT, which shows that although they can undergo cycles of reduction followed by correct alignment of disulfide bonds, their isomerase activity is significantly less than their oxidase activity, and likely to be less significant than its oxidoreductase activity. However, in cells, the direction of the reaction to oxidation or reduction will depend on the overall Eh , the redox state of the CXXC motif of the hBCAT proteins, its microenvironment, and also other chaperone and redox repair systems present, which requires further investigation.
Co-localization of hBCATm with PDI in neuronal cells was curious, as PDI is considered to be largely associated with the ER; however, it has also been localized to mitochondria and the cytosol. Although the cytosolic and mitochondrial proteins are the two key mammalian isoforms, other spliced variants of hBCAT homologous to hBCATm have been described, namely a novel alternative splice variant found in placental tissue and a splice variant that acts as a co-repressor of thyroid hormone nuclear receptors (39, 59). It remains unknown whether another hBCAT isoform could exist and reside in the ER or whether the predominant isoforms participate in their PDI-like activity elsewhere in the cell, as disulfide bond formation can also occur in mitochondria and outside the cell in the blood (1, 36). However, here, using Mia40 as a control mitochondrial protein, PDI was located to the mitochondria using electron microscopy and in close proximity (≤50 nm apart) to hBCATm, suggesting co-localization. These findings along with confocal microscopy confirm that the interaction between PDI and hBCATm is specific to the mitochondria. Co-immunoprecipitation assays validated direct binding, not only in neuronal cell cultures but also from human brain tissue. Moreover, the fact that PDI was identified in neuronal and endothelial cells, where both hBCATs reside, suggests the clear potential for this interaction to be physiologically relevant. We hypothesize that hBCATm and PDI could operate through dithiol-disulfide exchange, and hBCATm could potentially act as a chaperone to assist PDI in facilitating protein folding or in its own capacity act as a dithiol-disulfide oxidoreductase.
The importance of molecular chaperones and PDI is reflected in their role for the correct folding of proteins and the elimination of misfolded or irreversibly damaged proteins from the cell (44). Accumulation of aggregates such as α-synuclein and synphillin-1 in Parkinson's disease and amyloid-β and hyper-phosphorylated tau in AD along with inclusion bodies associated with amylotrophic lateral sclerosis (ALS), and prion diseases are common features of these neurodegenerative conditions (35, 44). Molecular chaperones such as the heat shock proteins (HSP70 and HSP90, in particular) and PDI are up-regulated in response to cellular stress and believed to function as neuroprotectors in these diseases [reviewed in Andreu et al. (2)]. These systems operate as a network with the unfolded protein response, which is directed to the clearance of modified or incorrectly folded proteins. However, although these systems are up-regulated, evidence suggests that oxidative and S-nitrosative stress may impact protein function, thus compromising their neuroprotective role (46, 47, 56, 63). One such example is a study by Uehara et al., which showed that PDI was S-nitrosylated in the brains of patients with Parkinson's disease and AD (63). Modeling of this mechanism in SH-SY5Y cells resulted in the accumulation of protein aggregates, supporting their hypothesis that S-nitrosylated PDI contributes to protein misfolding and has the potential to play a role in the pathogenesis of AD. In a separate study, PDI-SNO was also detected in the spinal cord of ALS patients.
In rat models, up-regulation of hBCATc has been related to cell survival. A report by Kholodilov et al. showed an increase in mRNA expression of BCATc in the substantia nigra of rats after developmental striatal target injury (34). Although this correlated with cell death, the neurons that were immunopositive for BCATc had normal physiology and rarely contained apoptotic chromatin, suggesting that BCATc played a neuroprotective role. Furthermore, administration of brain-derived neurotrophic factor (role in neuronal survival, growth, and differentiation) after a lesion of the visual cortex in newborn rats resulted in the up-regulation of BCATc (10). Increased expression was also observed in the brain of transgenic rats overexpressing BDNF in a time- and region-dependent manner (43). Collectively, these studies support the role of the BCAT proteins in maintaining neuronal survival. Recent studies by our group have demonstrated that both hBCAT isoforms show increased protein expression (up to 140% for hBCATm) in the brain of patients with AD relative to matched control patients (n=30, frontal and temporal region) (Conway, unpublished data). Based on previous studies, we suggest that the expression of hBCAT is increased in response to changes in the cellular redox environment, to assist in neuronal protection. We speculate that since the hBCAT proteins have oxidoreductase activity and can refold proteins, they may play a role in facilitating thiol-disulfide exchange either independently or as a redox chaperone to PDI or Mia40. However, similar to PDI, the hBCAT proteins are regulated through oxidation and S-nitrosylation (13, 15, 16), such that under increased cellular stress, their proposed neuroprotective role, and their association with these proteins would be affected. Therefore, knowing that the function of these proteins is compromised under cellular stress highlights the impact during pathogenic conditions where protein misfolding features. Future studies will investigate and characterize the role of redox-modified hBCAT with potential redox partners and protein folding in cells.
In conclusion, we have identified that hBCAT proteins have novel dithiol-disulfide isomerase activity that plays a role in protein folding. We demonstrate that S-glutathionylation is not only present to protect the function of a redox-sensitive protein, a concept held in high regard by most groups, but it has also shown value mechanistically and was shown here to be integral to the refolding of proteins catalyzed by hBCAT. Moreover, we have demonstrated that hBCATm is localized with other redox-sensitive proteins such as PDI and Mia40 in the mitochondria. These findings may have physiologically relevant implications in the human brain, but more importantly in protein misfolding, a key mechanism that is associated with AD pathology.
Materials and Methods
Materials
Cytidine 2′,3′–cyclic monophosphate (cCMP), RNase, bovine PDI, nonessential amino acids, fetal calf serum, 100× penicillin/streptomycin solution, glutaraldehyde, sodium borohydride, sodium cacodylate, Tween®20, and Triton X-100 were obtained from Sigma-Aldrich (Dorset, UK). RPMI 1640 was obtained from Lonza (Gloucestershire, UK). Human PDI (hPDI) was obtained from Cambridge Bioscience (Cambridge, UK). IMR-32, neuroblastoma cell line (ATCC® CCL-127™) was obtained from LGC Promochem (Middlesex, UK). NuPAGE® LDS sample buffer (4×), NuPAGE Novex® 4%–12% Bis-Tris 1 mm gel, 20× NuPAGE MES SDS running buffer, and 20× NuPAGE Transfer Buffer were obtained from Invitrogen (Paisley, UK). Electron microscopy (EM) grade paraformaldehyde was purchased from BDH (Leicestershire, UK). Acetylated bovine serum albumin (BSA) (10%) was purchased from Aurion (Wageningen, The Netherlands). Peroxidase, substrate 3,3′-diaminobenzidine (DAB), donkey serum, 4′,6-diamino-2-phenylindole (DAPI)-containing mounting medium, biotin-labeled secondary antibody raised to IgG, and avidin (Vectastain ABC kit) were purchased from Vector Labs (Peterborough, UK). Hematoxylin, Clearene, and Clearium were purchased from SurgiPath (Peterborough, UK). BM chemiluminescent horse radish peroxidase (HRP) substrate was purchased from Roche (West Sussex, UK). Hyperfilm was obtained from GE Healthcare (Cardiff, UK). All antibodies used in these studies are summarized in Supplementary Table 1. All other materials were purchased from Fisher Scientific (Loughborough, UK).
Preparation and activity of rdRNase and scRNase
Twenty milligrams of RNase was incubated with 150 mM DTT and 6 M guanidine hydrochloride over 18 h at room temperature (RT) in 0.1 M Tris, pH 8.6, desalted into 0.01 N HCl, and stored under argon at −80°C. To prepare scRNase, rdRNase in 6 M guandine-HCl was adjusted to pH 8.6 with solid Tris, sparged with oxygen for 1 h at RT, and allowed to re-oxidise in the dark for 4 days. The thiol concentration of scRNase was assessed before and after oxidation by titration with DTNB (18). RNase activity was determined by the protein refolding assay (detailed in 41, 42, 49, 50). Briefly, a final concentration of 1.4 μM RNase was added to 0.44 mM cCMP that was prepared in 0.1 M 4-morpholinepropanesulfonic acid, pH 7.0 with the increase in absorbance at 284 nm monitored over 10 min.
Reactivation of rdRNase catalyzed by native hBCAT and mutant proteins
All hBCAT and mutant proteins were exchanged into 0.1 M Tris-HCL, pH 7.4 with 1 mM EDTA before incubation with rdRNase. Typically, for each refolding assay, 30 μM rdRNase was incubated at RT with test protein and aliquots were removed to assess RNase activity at 24 and 48 h. The effect of time (12–48 h) and concentration (4–24 μM) on this reaction was assessed under these conditions. The redox environment was also varied using five different GSH/GSSG ratios (1:0, 3:1, 1:1, 1:3, and 0:1). In a separate experiment, the hBCAT proteins were pretreated with 500 μM GSNO for 30 min at RT before incubation with rdRNase. Control samples for the hBCAT and mutant proteins±rdRNase, rdRNase alone, for each GSH/GSSG ratio, and GSNO were included for each time point and each concentration. Subsequently, using a GSH/GSSG ratio of 3:1, the apparent Vmax for rdRNase and scRNase was determined by holding hBCAT at 10 μM with the substrate (either rdRNase or scRNase) concentration between 5 and 60 μM. A standard curve was generated from the initial rates of a range of RNase concentrations (0–4 μM). Using the slope, the concentration of active RNase formed/μM of protein was calculated. Western blot analysis of these samples was carried out as described in Supplementary Data.
Far Western immunoblotting
Human BCAT (1.0–3.5 μg, Fig. 5A) or hPDI (0.1–0.5 μg, Fig. 5B and constant at 0.4 μg, Fig. 5C) was applied to a nitrocellulose membrane. After blocking with 1× Tris buffer saline with tween (TBST) containing 5% BSA for 1 h at RT, the membrane was incubated with 5 μg/ml of PDI, hBCATc, or hBCATm, respectively, for 90 min at 37°C, with shaking at 100 rpm. After washing, bound hBCAT was detected by incubating with anti-hBCAT (1/1000 dilution) or for the hBCATs, anti-PDI-2 (1/1000) shaking, overnight at 4°C. Bands were visualized using BM chemiluminescent HRP substrate and exposed to HyperFilm.
Immunocytochemistry of hBCAT and PDI in neuronal cells
IMR-32 cells were allowed to reach 70%–80% confluence and treated +/− hydrogen peroxide (50–100 μM) and GSNO (100–500 μM) for 30 min, respectively, at 37°C in a humidified incubator that was adjusted to contain 5% carbon dioxide. After serial washes, the cells were fixed with 0.25% glutaraldehyde and permeabilized with 0.2% Triton X-100 for 20 min at RT in a 0.1 M sodium cacodylate buffer, pH 7.4 adjusted to contain 0.1 M sucrose. The cells were further treated by a succession of washes with sodium borohydride (1 mg/ml in PBS) and blocked in 3% BSA overnight before incubating with primary antibodies, anti-hBCATc, anti-hBCATm-1 (1/500), anti-PDI-1 (1/150 dilution), anti-PDI-2, anti-Mia40 (1/100), and anti-hBCATm-2 (1/250), respectively, for 1 h at RT. After serial washes, the cells were incubated with goat anti-rabbit Alexa Fluor® 568 (hBCATc, hBCATm-1, PDI-2, and Mia40, 1/500 dilution) and goat anti-mouse Alexa Fluor 488 (PDI-1 and hBCATm-2, 1/500 dilution) for 1 h at RT. Slides were washed in cacodylate buffer and mounted in 300 nM DAPI in glycerol. The Mander's correlation coefficients (Mx and My) were derived using Volocity (Perkin-Elmer) from an average of 15 individual cell images.
Electron microscopy of hBCAT and PDI in neuronal cells
Cells that had been previously incubated +/−100 μM of hydrogen peroxide for 30 min were fixed in a final concentration of 2% formaldehyde and 0.2% glutaraldehyde in phosphate buffer and processed according to the Tokuyasu method for cryosectioning (55, 60). Seventy nanometre sections were prepared at −120°C with 1% methylcellulose in 1.2 M sucrose and transferred onto carbon-coated copper mesh grids. The sections were labeled with primary antibodies to anti-hBCATm-1-3, anti-Mia40, anti-PDI-1, and anti-PDI-2 (1/10), respectively. Gold-labeled secondary antibodies, 6 and 10 nm (1/20), respectively, were incubated for 1 h at RT. All antibodies were prepared in 0.1% acetylated BSA. The sections were counterstained with 0.3% uranylacetate in 2% methylcellulose on ice and imaged on a CM10 (Philips) electron microscope.
Mammalian expression of HIS-tagged hBCATm using Gateway™ technology
According to the manufacturer's instructions, the LR recombination reaction contained 50–100 ng of purified hBCATm pENTR221™ entry vector (synthesized and prepared by GeneArt, Life Technologies), 150 ng of pDEST26 Gateway destination vector and TE buffer, pH 8.0 was used to adjust the final volume to 8 μl. Positive controls containing 100 ng pENTR-gus with 150 ng pDEST26 and negative controls containing 150 ng of purified BCATm pENTR221 with 150 ng pDEST26 were also included. The reaction was started with 2 μl of LR Clonase® II enzyme mix (with exception to the negative control). The samples were incubated in a PCR hot block at 25°C for 3 h. To stop the LR reaction, 1 μl of proteinase K solution was added and incubated at 37°C for 10 min. The completed LR reaction (1 μl) was subsequently transformed into Library Efficiency® DH5α™ Cells according to the manufacturer's instructions. Plasmids were purified using the PureLink™ Hi-Pure plasmid mini-prep purification according to the manufacturer's instructions, and restriction analysis was used to confirm correct orientation of the insert. IMR-32 cells were subsequently transfected with hBCATm expression plasmid using jetPRIME® transfection reagent (Polyplus) and following the protocol prescribed by the manufacturers. Controls included transfection reagent alone, cell only, and pcDNA3-EGFP to assess transfection efficiency.
Co-immunoprecipitation of hBCATm with PDI from IMR-32 cell lysates and human brain homogenates
Cell lysates (800 μg) from IMR-32 cells or brain homogenates (800 μg) were incubated with anti-hBCATm-protein G Mag Sepharose beads overnight. Total cell lysates, bound protein, and pure overexpressed hPDI were separated on a 6.5% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel and transferred onto a nitrocellulose membrane for Western blot analysis as described earlier. Primary antibody (1/1000 for hPDI) was prepared in 5% nonfat milk powder in 1× TBST and incubated overnight at 4°C before several washes with 1× TBST. Secondary antibody, mouse anti-rabbit light chain specific (1:2000) was added for 1 h at RT and then washed with 1× TBST followed by a 1 h incubation with anti-mouse HRP (1/2000). The positive bands were visualized using BM chemiluminescent HRP substrate and imaged using hyperfilm.
Immunohistochemistry of the hBCAT isoforms and PDI
The study was approved by the North Somerset and South Bristol Research Ethics Committee. All brain tissue used in this study was from brains donated with consent to the South West Dementia Brain Bank in the University of Bristol, UK. Serial sections (7 μm) were taken from the temporal and cerebellum brain regions, placed in a 60°C oven overnight before immunohistochemical staining, and subsequently dewaxed in clearene (2×5 min) and dehydrated in 100% ethanol (2×3 min). Endogenous peroxidase was quenched in 0.09% hydrogen peroxide/methanol solution for 30 min at RT. The slides were pretreated with citrate buffer (10 mM sodium citrate, 0.05% Tween 20, pH 6.0), then washed (2×3 min) in PBS containing 0.154 M NaCl, 1.86 mM Na2H2PO4.2H2O, 7.48 mM Na2HPO4.12H2O, pH 7.1, and blocked with 10% horse serum in PBS for 20 min at RT. Sections were incubated at RT overnight with primary antibody (1/6000 for both hBCAT isoforms) in PBS. The sections were washed in PBS (2×3 min) and incubated with biotinylated antibody to IgG for 20 min (Vectastain ABC kit). The slides were washed again in PBS (2×3 min) and incubated with the avidin-biotin complex in PBS for 20 min (Vectastain ABC kit). Slides were developed with DAB/H2O2 in distilled water (DAB substrate kit) for 10 min before immersion in copper sulfate solution (16 mM CuSO4.5H2O, 0.123 M NaCl) for 4 min, and counterstained with Harris' hematoxylin (25% Gill haematoxylin). The slides were dehydrated in 100% ethanol (2×5 min), cleared in 100% Clearene (2×3 min), and mounted in Clearium. Sections were viewed and imaged on a Nikon Eclipse 50i or a Leica DMR microscope.
Footnotes
Acknowledgments
The authors would like to sincerely thank Leslie Poole, Wake Forest University, USA, for advice on rdRNase kinetics and the discussion of dithiol-disulfide exchange and also Sarah O'Neill, Royal College of Surgeons, Ireland, for technical assistance. They would also like to acknowledge the help of Paul Verkade and the EM technical staff at the Wolfson Bioimaging Facility, University of Bristol, UK. This research was supported by a Research PhD Fellowship grant awarded by Bristol Research to Alzheimer's and Care of the Elderly (BRACE), Bristol, UK, and an HEFCE-funded bursary from the University of the West of England to Myra E. Conway.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.