
Abstract
Advances in recombinant DNA approaches have resulted in the development of transgene architectures that severely bias their own inheritance, a process commonly referred to as “gene drive.” The rapid pace of development, combined with the complexity of many gene drive approaches, threatens to overwhelm those responsible for ensuring its safe use in the laboratory, as even identifying that a specific transgene is capable of gene drive may not be intuitive. Although currently gene drive experiments have been limited to just a few species (mosquitoes, flies, mice, and yeast), the range of organisms used in gene drive research is expected to increase substantially in the coming years. Here the defining features of different gene drive approaches are discussed. Although this will start with a focus on identifying when gene drive could or could not occur, the emphasis will also be on establishing risk profiles based on anticipated level of invasiveness and persistence of transgenes in the surrounding environment. Attention is also called to the fact that transgenes can be considered invasive without being considered gene drive (and vice versa). This further supports the notion that adequate risk assessment requires information regarding the specific circumstances a given transgene or set of transgenes is capable of invading a corresponding population. Finally, challenges in the review and evaluation of work involving gene drive organisms are discussed.
Introduction: Gene Drive
All students of basic genetics learn about Mendel's law of inheritance; even those who have never taken the subject directly will likely have some familiarity with the concept that they inherited traits from both of their parents relatively equally. Although simple, this concept has strong implications for the accidental release of an organism modified to contain recombinant DNA from a laboratory into the environment. Since the number of accidentally released individuals will likely be much, much smaller than the total number of wild individuals, each generation the number of organisms containing the modified gene will be reduced by half. This fact has likely contributed to the safety record of laboratory research using transgenic organisms over the past 40 years. 1 However, widespread in nature are selfish genetic elements that populate the genomes of animals, plants, and microorganisms. These elements have established methods for ensuring their own preferential inheritance through a variety of mechanisms. By emulating these naturally occurring methods, researchers have devised recombinant DNA approaches for engineering transgenes that also result in the preferential inheritance of modified DNA. Termed gene drive, after the naturally occurring phenomenon known as meiotic drive, such genetic manipulations pose unique challenges from the perspective of laboratory biosafety, both to the Principal Investigator (PI) and to the biosafety communities responsible for oversight. Although the first synthetic gene drives were developed in decades past,2,3 it is in the past 5 years that gene drive technology has become practical from an engineering standpoint across a wide range of organisms (reviewed in Refs.4,5). The rapid pace of development combined with the complexity of many gene drive approaches threatens to overwhelm those responsible for ensuring its safe use in the laboratory. Even identifying that a specific transgene is capable of gene drive is not trivial, further complicating the review and oversight process. This is made more challenging by the use of the term “gene drive” as shorthand to refer to one specific molecular architecture (CRISPR/Cas9 homing-based gene drive). 6 These verbal shortcuts have also led to the use of the term “gene drive” as a noun, rather than a verb. This only sows confusion, as it implies that gene drive is an intrinsic feature of a specific arrangement of recombinant DNA, when this is not the case. As discussed later in this article, even transgene sequences that are not capable of gene drive may present different risk profiles that warrant consideration.
Here I approach the subject of gene drive and other invasive genetic elements from the perspective of performing a biological risk assessment for laboratory-based research. Questions are presented to help in developing risk profiles and ultimately a risk assessment that an Institutional Biosafety Committee (IBC) could build off of during their next protocol review involving gene drive. Although primarily targeted to the biosafety community at large, the identification and classification of invasive genetic elements based on risk profiles and defining features should also be of value to PIs and laboratory staff, who will be in a better position to communicate their work and the salient points to their oversight committees.
Gene Drive Is Not a “Thing,” It Is a Process Encompassing a Broad Category of Things
It cannot be overemphasized that due to the immense variation in the potential of different transgenes to result in gene drive, the phrase “gene drive” itself has little practical meaning in the context of performing a specific risk assessment. Although the biosafety professionals and committees that will be asked to review and approve gene drive research may not be intimately familiar with the term “gene drive” or the issues surrounding it, 6 what is important is that the field of biosafety recognizes that “gene drive” transgenes are not a singularity, but that the process of gene drive can be achieved through an extremely diverse category of genes, aka “agents” (reviewed in Ref. 4 ). To put this in perspective, if during an introductory meeting with a biosafety professional, a PI states that their laboratory plans to work with “microbes,” it is unlikely that the biosafety officer (BSO) would be able to offer much help in setting containment levels until more specific information was provided. For example, were they planning on working with a bacteriophage, a nonpathogenic plant-specific virus, or a hemorrhagic fever virus? The fact that more information is required will be obvious to the readership here, the same is true had the PI provided other noninformative broad encompassing terms, such as “bacteria,” “parasite,” “pathogen,” or “animal.” We intuitively recognize these terms as generic placeholders and use them as shorthand after establishing the agent in question, but these terms in isolation are not enough to perform the risk assessment. The same holds true for the term “gene drive,” which provides just as little information. Thus, rather than use a binary question such as “gene drive or not a gene drive,” any risk assessment must instead ask more nuanced questions, just as they would for pathogens, bacteria, viruses, or similar categories.
Within the category of transgene architectures capable of gene drive, we can anticipate two main parameters concerning the behavior of the respective transgenes that will be important for risk assessment: (1) how fast might the transgene spread if introduced into the surrounding environment and (2) how long might the transgenes last if introduced into the environment? As indicated in Figure 1, there is anticipated to be a wide range of answers to these questions depending on the specific technology used. It is easy to imagine that a gene drive transgene inherited by 99% of all progeny will spread faster than one inherited by 51% of progeny. Both may be considered to have the potential to spread in the environment, but practically speaking the former may warrant a different risk management approach as compared with the latter.
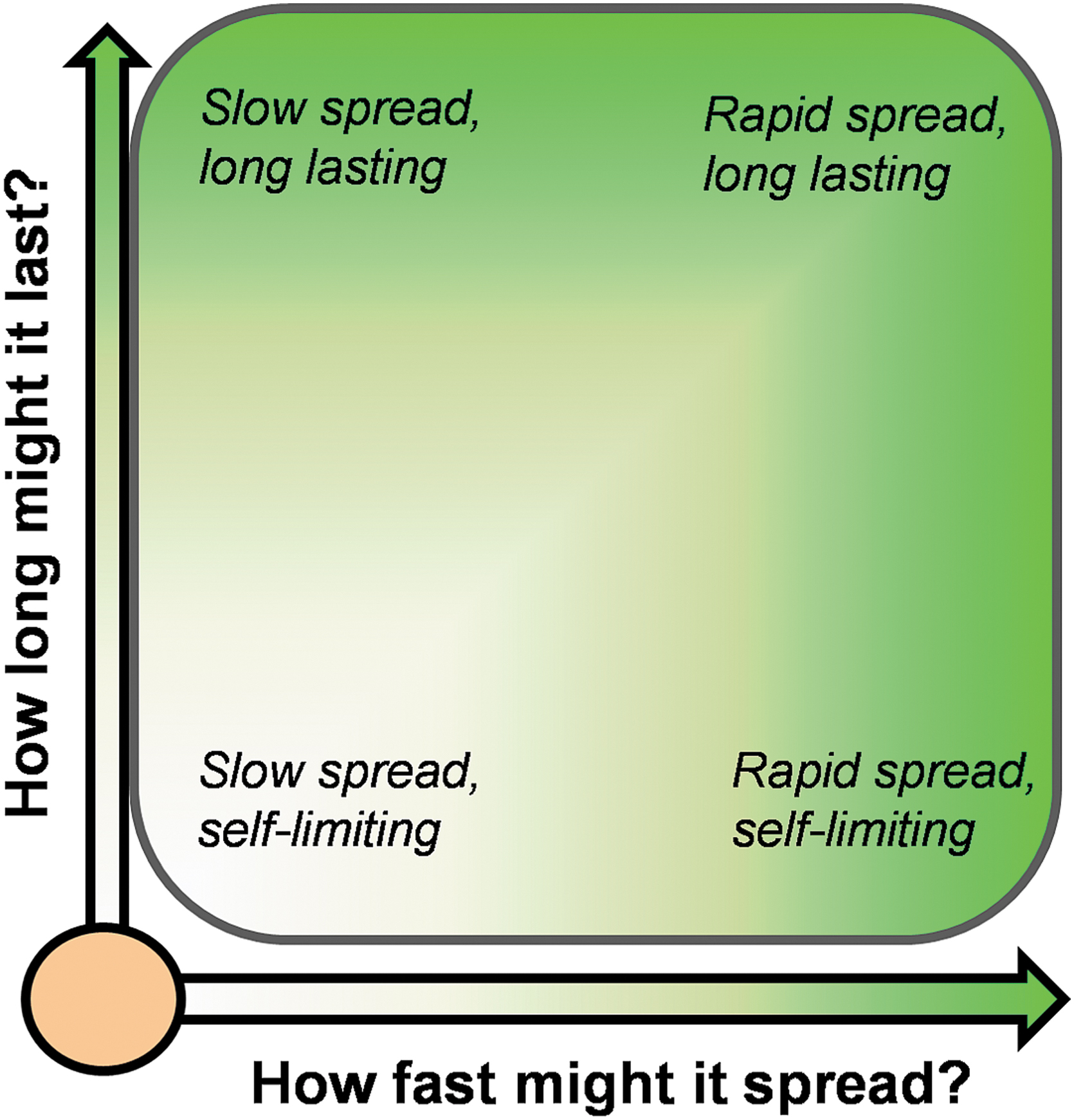
Gene drive is not a single thing, but can be achieved through a broad category of things similar to terms such as “microbe,” “bacteria,” or “virus.” Defining traits of gene drive important for risk assessment are how fast it might spread in the surrounding area (if at all) and how long it might last in the surrounding area if introduced. Although the molecular architectures underlying the process of gene drive are constantly being revised and improved, they will always fall somewhere on this continuum.
Note that other factors besides the arrangement and type of transgenes may influence the likelihood of spread and persistence in the environment. The biology of the organism itself is an important consideration, as the same transgene placed in the genome of a small organism with fast generation times (flies and mosquitoes) would present a completely different risk profile than if it were to be placed in large vertebrates (cow). Although most reports of gene drive have focused on the former, establishing gene drive is possible in any sexually reproducing organism. Thus, the first challenge faced in the oversight of laboratory research is just identifying that the process of gene drive may occur given a specific set of transgenes and a particular host species.
How to Recognize Whether a Recombinant DNA Modification Will Result in Gene Drive
Some laboratories will set out to engineer a transgene or set of transgenes into a target organism for the purpose of evaluating gene drive. In these cases, a PI may state directly on their IBC submission that this is a goal and describe how gene drive will be achieved. In other cases, researchers may be aware of the concept of preferential inheritance but not explicitly use the term gene drive, whereas in still others, the process of gene drive may be an unanticipated consequence of a set of transgenes originally conceived to answer a separate experimental question. Thus, a simple question on an IBC application such as “does your work involve the generation of a gene drive” may not be sufficient, as only those familiar with the term and intentionally pursuing this line of work are in a position to answer completely. More useful might be questions that get at the definition of gene drive, such as “will the transgene or combination of transgenes result in the preferential inheritance of recombinant DNA?” This question can be answered by those familiar with the term gene drive as well as those who are not, and forces the PI to reflect upon their work and recognize gene drive potential if it is not intended. Although the diversity of gene drive approaches being developed continues to grow, these can (at present, at least) be categorized very roughly into just two main groups: homing-based gene drive and selective survival (Box 1).
However, as already mentioned above, the mere fact that some level of preferential inheritance is possible is not sufficient to inform the risk assessment, and thus a follow-up question such as “if so, how, and under what circumstances will preferential inheritance occur?” This makes it clear that gene drive is not an intrinsic feature of a recombinant DNA sequence: the exact same transgene may be capable of inducing the process of gene drive in some circumstances but not in others. Take the case of a homing-based transgene as shown in Figure 2. In a heterozygous individual, the presence of a nuclease (could be Cas9, but does not have to be) triggers a break on the opposite chromosome, and the resulting homology-based repair generates a second copy of the transgene. Notice that in all three cases, there will be preferential inheritance of at least one of the transgenes due to this increase in copy number. However, each has different requirements for how this preferential inheritance occurs, and these substantially alter their corresponding risk profiles.
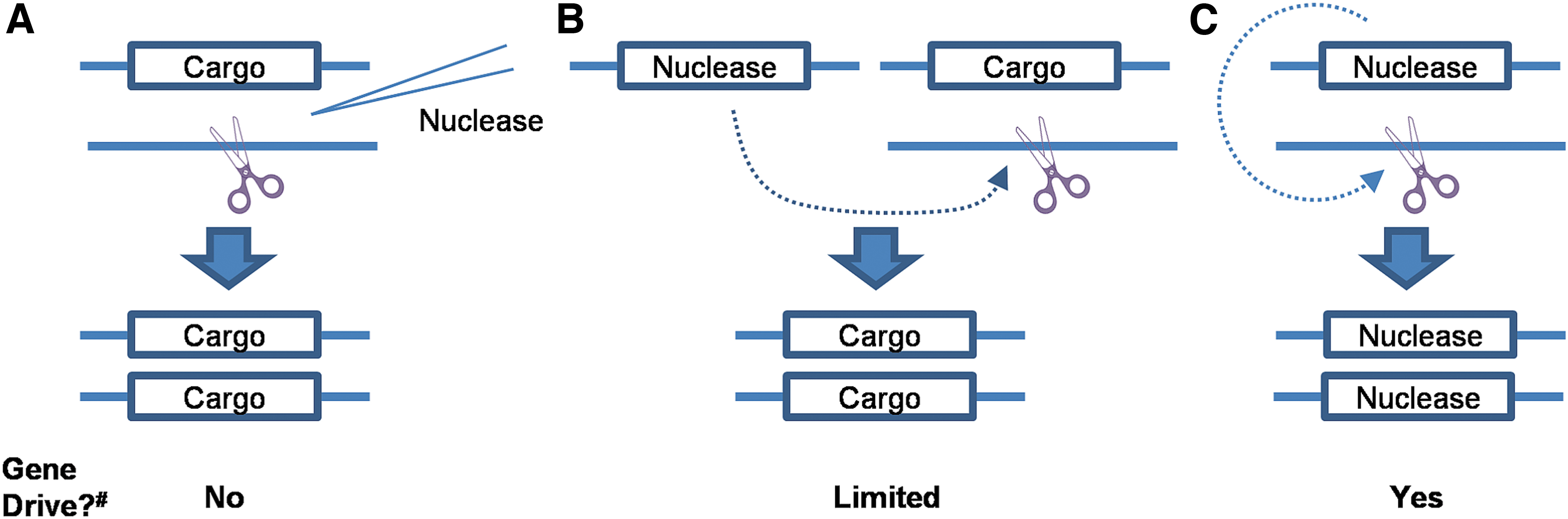
Recognition of homing-based gene drive approaches. In all cases, a nuclease is introduced to make a double-stranded DNA break in a chromosome, resulting in the copying of a target gene.
In the simplest case, the nuclease is provided as messenger RNA or protein and introduced into the organism during the embryonic stages (Figure 2A). This results in preferential inheritance in a single generation only, not sustained gene drive. In the second case, the nuclease is encoded in trans (i.e., it is encoded in a separate location in the genome) and introduced through a genetic cross, a so-called split-drive (Figure 2B). Here, gene drive will occur only in individuals who inherit the nuclease gene, which itself is not subject to gene drive. If these individuals escaped the laboratory, gene drive would continue only as long as the nuclease persisted in the genome; once lost, gene drive could no longer occur. In the final example, the gene encoding the nuclease is placed at the allelic position where it generates a break. In this case, these individuals will be capable of gene drive in every generation. This scenario represents the highest risk profile of the three. For homing-based gene drive, how the nuclease is encoded or delivered is thus a critical factor in when gene drive can occur.
In addition to the requirement for a nuclease, a defining feature of homing-based gene drive is the presence of a target site (DNA sequence recognized by the nuclease) in the genome. Thus, a transgene that might be considered highly invasive when the target site is present could also be completely incapable of gene drive when the target site is absent, with obvious implications for risk assessment. In simple cases, a target site may be an engineered sequence present only in a laboratory population and completely absent from any corresponding wild-type population. Thus, the process of gene drive could only occur in this laboratory population. In other cases, the target site(s) may be present as polymorphisms in some populations but in not others, whereas yet other target sites may be conserved not only in the target population but also in related species capable of hybridization.
The target site may be in an essential gene, or important for one sex only. In the latter case, successful gene drive could be associated with extreme sex bias and potentially population collapse.7,8 In addition, target sites can be repaired through mechanisms other than gene drive resulting in resistance to the nuclease. Thus, any risk assessment for homing-based approaches needs to fully consider both the source of the nuclease and the nature of the target site (Box 2). Readers are advised to keep in mind that the three examples shown in Figure 2A–C are only a subset of possible ways to implement homing-based gene drive, but the defining features (nuclease + target site) will always be present.
Gene Drive Is More Than Just Homing
Homing-based gene drive increases the copy number of a gene and is dependent upon host DNA repair machinery for this to occur. Other forms of gene drive have been described that do not increase transgene copy number and do not depend on DNA repair outcomes, but instead apply strong selection pressure on gametes or developing embryos to ensure the preferential survival of individuals carrying the transgene sequence(s). Although the technical implementations of these selective survival approaches are varied and complex, some basic principles can be distilled and these will help inform the risk assessment process. Selective survival approaches depend on the expression of one or more “toxins” that impose a strong fitness cost on the organism, typically resulting in lethality. Coexpressed with this “toxin” may be one or more “antidotes” that gives the organism the ability to survive in the presence of the toxin. Note that these terms are conceptual shortcuts and do not refer directly to the toxins produced by pathogenic microorganisms more familiar to biosafety professionals. Instead, the term “toxin” is a placeholder for any gene product that causes the loss of an essential activity in a cell/organism. For example, the “toxin” could be a nuclease that disrupts an essential gene, 9 a microRNA (miRNA) that silences the expression of an essential gene, 3 or a protein that inhibits the activity of an essential gene product. The antidote may interact directly with the toxin to prevent its activity(ies), or it may act by providing the missing function in a manner that is resistant to the toxin. To use the same examples, an “antidote” could be a gene that has been recoded to no longer contain the nuclease recognition site or miRNA target site, or whose protein product can no longer be bound to the peptide toxin (Figure 3A). Importantly, on an IBC application, these activities can seem innocuous and unrelated to the process of gene drive, emphasizing the need for a more comprehensive synthesis of how any engineered recombinant DNA sequences will be utilized in combination, rather than a simple list of components. For example, by the definitions used here, some “toxins” may be expressed conditionally solely to understand the role of specific genes during development (Figure 3B), and these may be accompanied by transgenes that can rescue the developmental function in question (Figure 3C). By themselves, these are just basic research questions and do not involve gene drive. Only when the toxin affects individuals that do not encode the transgene, resulting in a selective advantage, do the same components result in gene drive (Figure 3D).
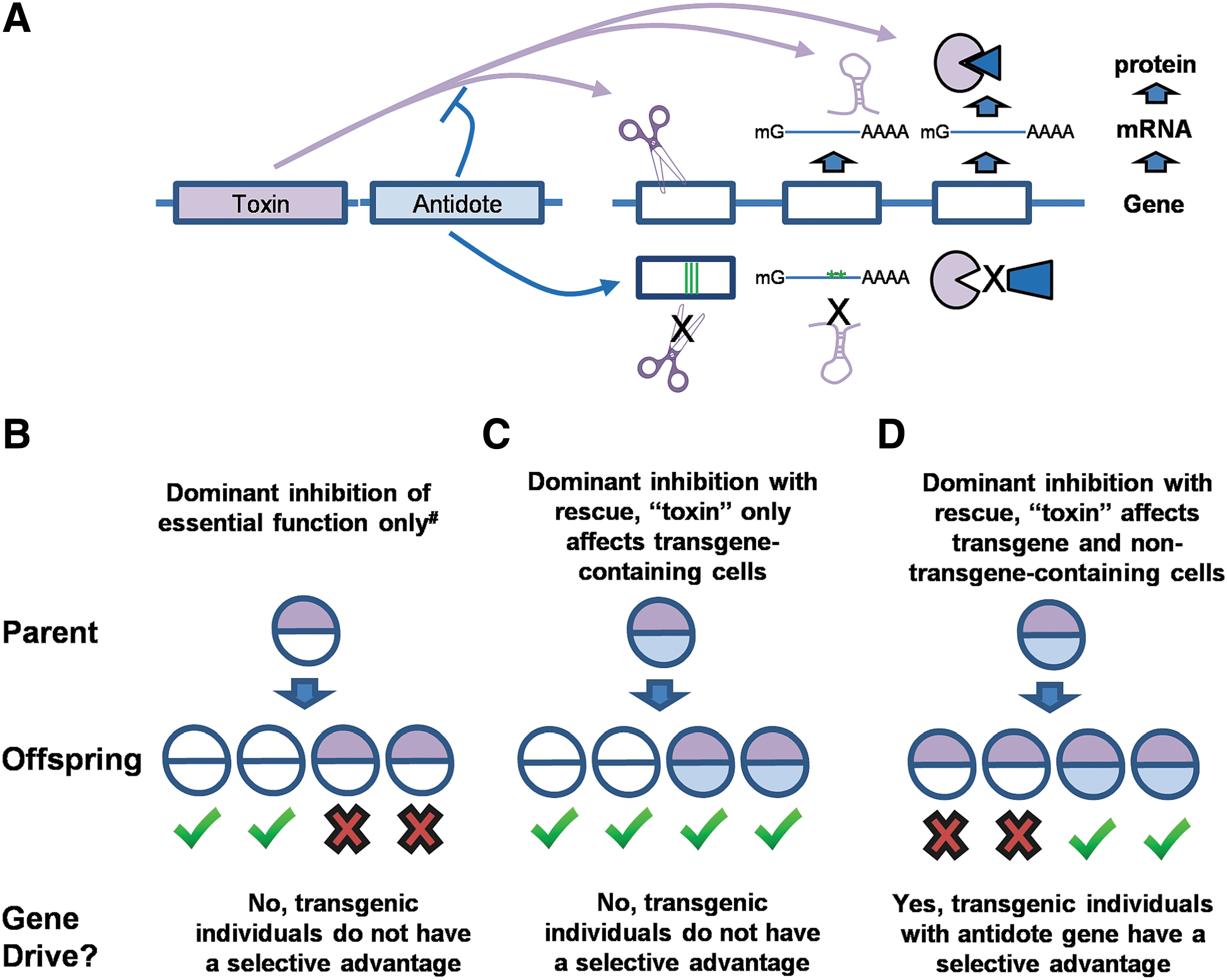
Recognition of selective survival-based gene drive approaches.
Alternatively, a toxin could preferentially target one sex, resulting in extreme sex bias. If these toxins are encoded on an autosome, this sex distortion will not result in gene drive. However, if encoded on a sex chromosome, then this bias can occur in each subsequent generation (Figure 4). For example, the expression of a gene that permanently deletes other genes critical for bloodmeal digestion in female mosquitoes may be completely innocuous to males, as they do not drink blood. If this gene was encoded near the male-determining region, then all daughters produced each generation may die upon bloodfeeding. Some key questions regarding selective survival-based gene drive are listed in Box 3.
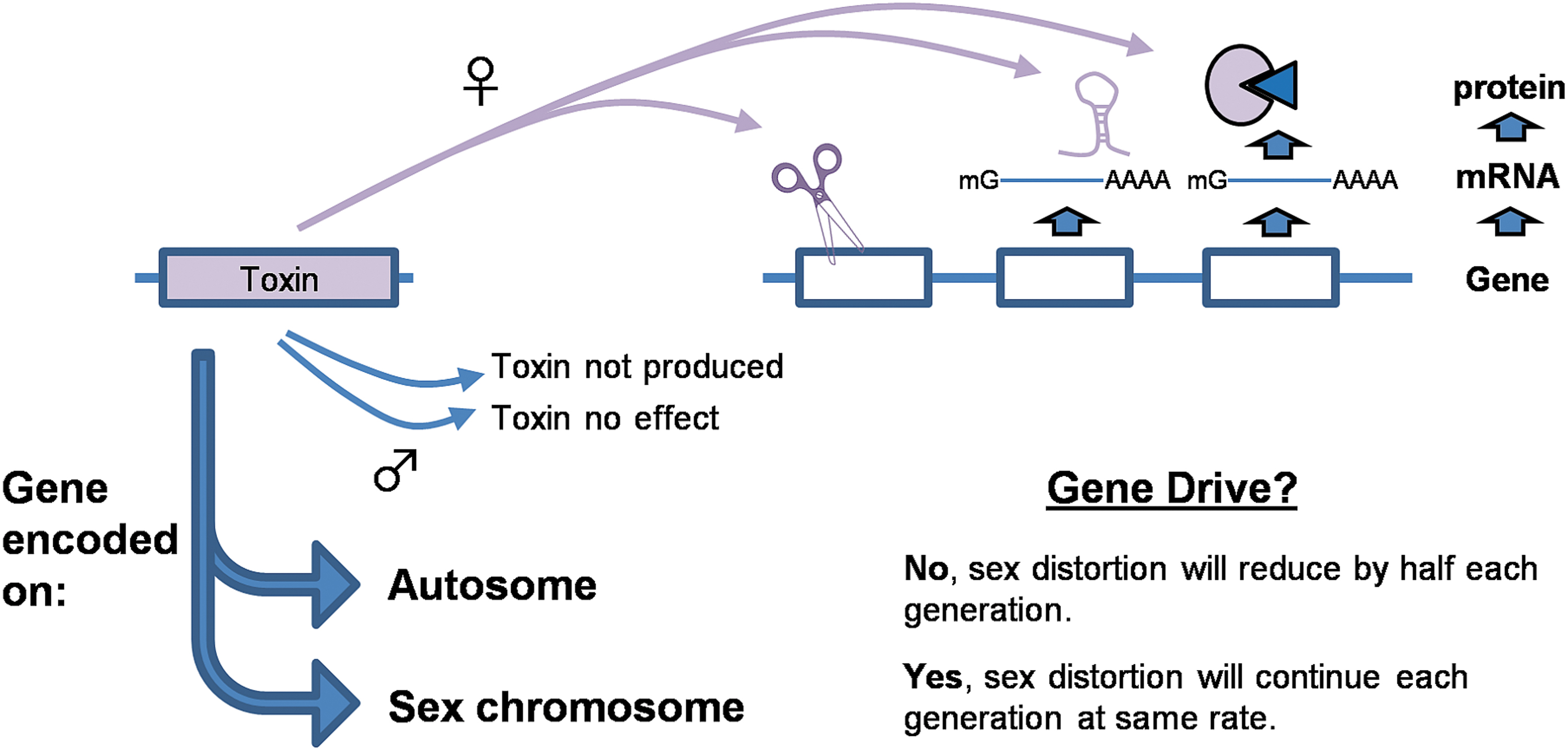
Recognition of sex distortion-based gene drive approaches. Transgene encodes one or more “toxin” genes that only result in the loss of an essential function in one sex, but not the other. This can be achieved by restricting expression of the toxin to a single sex, or by inhibiting a function only important to one sex. Gene drive will only occur if the transgene is placed at or near the region that determines the unaffected sex (e.g., a toxin that does not affect males placed on the Y chromosome, or equivalent region).
Gene Drive Is Part of an Even Broader Category of Invasive Genetic Elements
Given the mentioned information, it should be possible to reliably identify recombinant DNA manipulations that can result in gene drive under a given set of circumstances. However, when the risk assessment process is based on the level of invasiveness of the genetic element, and not on the mere presence of transgene architectures capable of sustaining gene drive in one or more potential settings, a new consideration emerges: engineered transgenes that are not “gene drive” also have the potential to be highly invasive. Transgenes that provide resistance to insecticides or pathogens, which increase lifespan, egg production, mating success, dispersal, or allow the use of new ecological niches, would all have the potential to spread in nature and thus permanently alter wild populations. Such gain-of-function manipulations receive comparatively little attention, even though they may be far simpler to implement since no complicated gene drive strategy is required. For example, a gain-of-function phenotype was recently reported for genetically modified An. gambiae, 10 where transgenic mosquitoes with enhanced immunity were found to have a mating advantage over their wild-type peers. In contrast, some forms of gene drive can be less invasive than a typical neutral transgene. 11 Although counterintuitive, these forms of gene drive only spread when they exceed a critical threshold; below the threshold they are driven in the opposite direction—out of the population. Thus, the mere presence or absence of a transgene or set of transgenes that can or cannot sustain gene drive is also not sufficient for the risk assessment process.
Beginning the Risk Assessment Process
Given that gene drive as a broad group is merely a subset of a larger category of potentially invasive transgenes, the process of risk assessment for any transgene may need to start with a question such as “is there a reasonable expectation that the genetic element (or a combination of genetic elements) can spread and/or persist in the surrounding area” (Figure 5). As already noted, this would be followed with the qualifiers “if so, under what conditions would this be expected” and “what are the anticipated consequences of such spread/persistence?” Since gene drive or other invasive genetic elements are extremely unlikely to pose any direct hazard to the experimenter, the answers to these questions become the driving force in determining containment conditions, work practices, and the need for any additional mitigation strategies. Unlike risk group designations (RG1–4) for eukaryotic pathogens that provide a starting point for risk assessment and are tied to specific containment levels (BSL-1–4 for pathogens, ACL1–4 for arthropods containing pathogens), no such shortcuts are currently available for invasive transgenes.
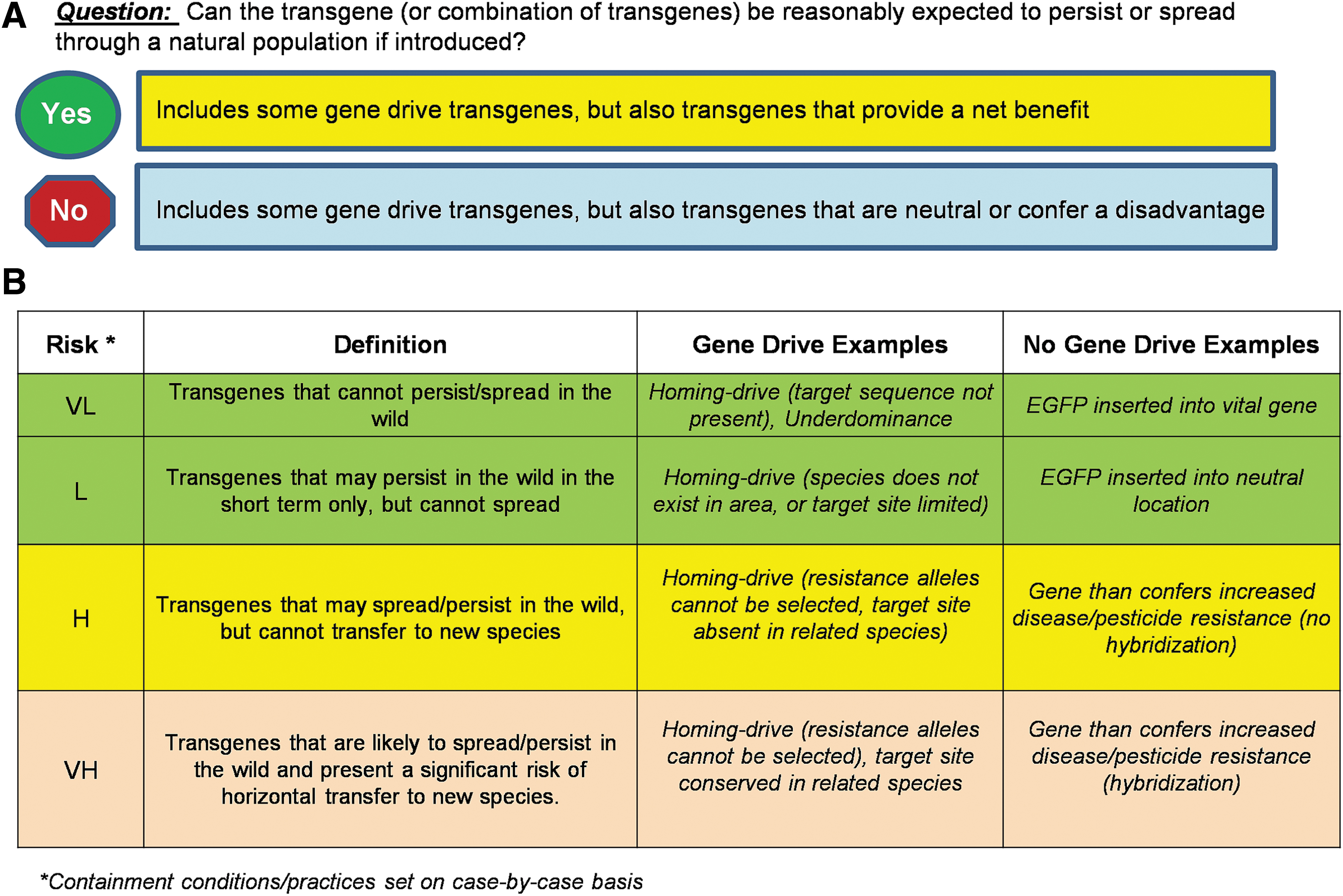
A starting point for risk assessment of laboratory-based transgenic organisms.
Although current gene drive protocols use containment similar to ACL-2 (Ref. 12 ), many also borrow more stringent measures from ACL-3 on an as needed case-by-case basis. 13 As there is also substantial latitude in ACL-2 facility design and operations, establishing risk profiles of different invasive genetic elements may be helpful in determining the adequacy of a particular facility and set of work practices (Figure 5B). For example, a homing-based gene drive transgene that targets an engineered sequence not present in a wild populations would present a very low risk profile, as outside of the laboratory the transgene would not be invasive. In contrast, an invasive genetic element that is likely to not only spread/persist in the target population but that could potentially jump to new species would present a very high risk profile that would likely warrant additional containment practices or a modified facility design.
Setting Containment and Risk Mitigation Approaches
Once the scope of work is clear and the invasive potential for any proposed genetic manipulations articulated, it is the job of the PI to suggest, and the BSO/IBC to set the appropriate containment conditions where the work can be performed safely (or to reject the application if no such conditions exist). Again, in this case, “safety” does not necessarily refer to the health and safety of laboratory workers or the community, who are the traditional focus of biosafety, but to avoiding any potential unauthorized release of the organism in question and any associated potential hazards to the shared environment. Based on the previous experience level of both the PI and the BSO/IBC in handling/evaluating research using invasive genetic elements, the process of setting containment conditions may be associated with various degrees of uncertainty. Depending on the relative experience of each party, different substantial concerns may arise. For established laboratories where research with invasive genetic elements has been ongoing, safeguards against complacency are the major concern. This concern is addressed by regular refresher training and the use of tabletop exercises among laboratory staff to think through possible scenarios. In other cases, the PI, BSO/IBC, or both may or may not be as familiar with appropriate work practices, facility design/upkeep, or the organism in question, and thus care must be taken at the outset to validate protocols and verify facility operations before beginning work with invasive genetic elements. For example, a recently renovated facility intended to house experiments involving gene drive in mosquitoes may be required to rear wild-type mosquitoes only under the same conditions for a period of time to verify that protocols/procedures for handling/rearing are sufficient to maintain containment. A similar trial period can help validate or revise operating procedures regarding entry/exit or waste disposal (see Box 4 for an example of some common requirements).
As the majority of current gene drive research is being performed in arthropods, the arthropod containment guidelines 14 and two recent reports provide detailed guidance on both containment practices 13 and standard operating procedures 15 with regard to arthropods modified to contain some form of invasive genetic elements. Both were written with the biosafety community in mind, and both provide detailed practical advice that should be helpful in this process. For example, Benedict et al. 13 recommend separate rearing/manipulation areas for arthropods with invasive genetic elements as compared with unmodified arthropods to avoid contamination of colonies. In the same regard, care must be taken when shipping material from a laboratory that houses arthropods with invasive genetic elements. Even if the shipment is of a standard colony strain, if contaminated with an invasive element, the receiving facility may be unaware and/or unequipped to deal with it, ultimately resulting in an inadvertent release into the environment. To guard against this, Benedict et al. 13 recommend the development of diagnostic assays that can be used to definitively identify/rule out the presence of the invasive genetic element. As the development of gene drive approaches or gain-of-function transgenes moves into other organisms, the principles of containment and risk management can be adapted as needed.
Challenges for IBC Review
The composition of IBCs is geared toward the review of work with microbial pathogens, either in culture or in combination with animals/plants. This is for good reason, as this type of work presents the greatest hazard to laboratory workers and demands critical and expert oversight. However, restrictions on time and a constant focus on pathogen-based research may present challenges to evaluating what may be unfamiliar potential hazards associated with invasive genetic elements (gene drive or otherwise). IBCs should be proactive in training their members on the identification and risk assessment process for invasive genetic elements.
Despite the lack of immediate hazards to laboratory personnel, IBCs are responsible also for protecting the environment from recombinant and synthetic nucleic acids. Consequences of an inadvertent release of an arthropod or other organism modified with an invasive genetic element may be direct (decrease/increase in population numbers, change in behavior/transmission potential) and/or indirect (effects on predators, microbes, or other species that interact with the modified organism in question). Evaluating these potential consequences and their likelihood may involve expertise not usually found on IBCs, such as ecologists and invasive species biologists. Identifying and involving these individuals early can help direct the risk assessment process and point to key data/information needed to complete it.
Future Outlook
For many decades, engineering invasive genetic elements was limited to the aspirations of a small group of vector biologists. Although major breakthroughs in both selective survival 3 and homing-based gene drive 16 were made a decade or more ago, it was the proof-of-principle experiments with CRISPR/Cas917–19 that have captured the imagination of the wider world. As these initial designs have given way to more powerful approaches to achieving gene drive,8,20–24 we can expect that the number of laboratories aiming to incorporate the development or study of invasive genetic elements will only continue to grow, with organisms such as wasps, 25 snails, 26 mice, 27 and plants 28 on the horizon and likely more to follow. As that happens, it is imperative that the biosafety professionals charged with evaluating work practices and containment facilities keep grounded in the underlying concepts of gene drive and other invasive genetic elements from the perspective of risk assessment.
Footnotes
Acknowledgments
I thank Christine McFarland and Jessica Bourquin of the TAMU Office of Biosafety for their input and comments on a draft of this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.