
Abstract
Eradication of human immunodeficiency virus type 1 (HIV-1) by vaccination with epitopes that produce broadly neutralizing antibodies is the ultimate goal for HIV prevention. However, generating appropriate immune responses has proven difficult. Expression of broadly neutralizing antibodies by vaginal colonizing lactobacilli provides an approach to passively target these antibodies to the mucosa. We tested the feasibility of expressing single-chain and single-domain antibodies (dAbs) in Lactobacillus to be used as a topical microbicide/live biotherapeutic. Lactobacilli provide an excellent platform to express anti-HIV proteins. Broadly neutralizing antibodies have been identified against epitopes on the HIV-1 envelope and have been made into active antibody fragments. We tested single-chain variable fragment m9 and dAb-m36 and its derivative m36.4 as prototype antibodies. We cloned and expressed the antibody fragments m9, m36, and m36.4 in Lactobacillus jensenii-1153 and tested the expression levels and functionality. We made a recombinant L. jensenii 1153-1128 that expresses dAb-m36.4. All antibody fragments m9, m36, and m36.4 were expressed by lactobacilli. However, we noted the smaller m36/m36.4 were expressed to higher levels, ≥3 μg/ml. All L. jensenii-expressed antibody fragments bound to gp120/CD4 complex; Lactobacillus-produced m36.4 inhibited HIV-1BaL in a neutralization assay. Using a TZM-bl assay, we characterized the breadth of neutralization of the m36.4. Delivery of dAbs by Lactobacillus could provide passive transfer of these antibodies to the mucosa and longevity at the site of HIV-1 transmission.
Introduction
N
Thus, new strategies are needed for providing neutralizing antibodies at the mucus surface where HIV is transmitted. One innovative avenue is to use a recombinant commensal Lactobacillus as a vehicle for releasing neutralizing antibodies at the mucosa. Lactobacillus species are common members of the vaginal microbiota in healthy women of childbearing age 7,8 and interact with the host to reduce proinflammatory cytokine levels and serve as the major host inflammatory modulator. 9 Lactobacillus jensenii is one of the more predominant vaginal lactobacilli. The strain L. jensenii 1153 has been proven as a potential platform to deliver sufficient levels of the anti-HIV protein-modified cyanovirin-N to mucosal surfaces to protect macaques from Simian (S)HIV challenge. 10
HIV-1 entry into target cells requires the sequential binding of the viral envelope glycoprotein gp120 epitopes to CD4 and then to the CCR5 chemokine receptor. 11 When CD4 binds to the HIV envelope, a conformation change in gp120 exposes the cryptic epitopes known as the CD4-induced (CD4i) site. 12,13 When a CD4i antibody is present, it recognizes these previously unexposed sites, blocking virus attachment of HIV to the chemokine receptor. 14
Different variants of anti-HIV antibodies have been developed, such as single-chain variable fragments (scFvs), small antibody fragments, and even smaller domain antibodies (dAbs). These alternatives offer advantages over conventional antibodies because of their smaller size, robustness, and their ability to be expressed in bacteria. Moreover, the small size of the dAbs and scFvs overcomes the steric constraints when the virion is bound to CD4; an intact IgG molecule may not be able to access the CD4i bridging sheet. 15 Numerous scFvs and dAb variants and camel-derived antibodies have now been identified that retain the broadly neutralizing anti-HIV activity. 16 –22 Among these broadly neutralizing antibody fragments are ScFv-m9, 23,24 dAb-m36, 25,26 and dAb-36.4 variant 19 that bind CD4i regions and facilitate neutralization of a broad range of HIV isolates.
We selected these antibody fragments that target the highly conserved CD4i epitopes, for expression in Lactobacillus as entry inhibitors, since they are likely to be present across the wide genetic diversity of HIV-1 variants. In addition, their small size may put less strain on the recombinant Lactobacillus allowing for expression levels that are biologically relevant.
In this article, we optimized and compared the expression of scFV-m9, dAb-m36, and dAb-m36.4 by L. jensenii. We evaluated the breadth and potency of dAb-m36.4 against different HIV-1 subtypes. Finally, we constructed a recombinant L. jensenii 1153-1128 producing dAb AP-m36.4.
Materials and Methods
Bacterial strains, culture, plasmid construction, and transformation
L. jensenii strain 1153, a vaginal isolate from a healthy woman, was cultivated as described previously. 27 To express m9 or m36 antibody fragments in L. jensenii 1153, the m9 or m36 genes were cloned into the modular shuttle vector pOSEL175, a modified version of pOSEL144. 27 The expression cassette for secreted scFv-m9 or dAb-m36 contained the Lactococcus promoter p23, the signal sequence from the ribosome binding site to the signal peptidase cleavage site of the L. crispatus S-layer gene (CbsAss) (for protein secretion), and a C-terminal epitope tag DYKDDDDK (FLAG) with a TAA stop codon at the 3′ end.
For purification ease, Profectus Biosciences Inc. modified the cassettes for the m9, m36, and m36.4 genes to include a C-terminal albumin-binding domain (ABD), SA06 for improving its pharmacokinetic properties and also aiding in purification using rat serum albumin (RSA) columns. 28 The original m9 and m36 genes do not bind albumin. The OSEL shuttle plasmids were maintained and propagated in Escherichia coli, purified, and electroporated into L. jensenii 1153 as described before, 27 generating the transformed strains L. jensenii 1153-1126p (expressing m9), L. jensenii 1153-1127p (expressing m36), and L. jensenii 1153-1128p (expressing m36.4). Transformed lactobacilli were routinely propagated using Man Rogosa Sharpe (MRS) or Rogosa media (broth or agar) with 20 μg/ml of erythromycin. L. jensenii 1153-1128 was propagated on MRS or Rogosa broth or agar without antibiotic.
Western blots
Stationary-phase supernatants from transformed (plasmid expressed) L. jensenii 1153-1126p, L. jensenii 1153-1127p, and L. jensenii 1153-1128p were harvested and centrifuged to remove cell debris. Thirty microliters of supernatant was run on a 12% Bis-Tris Gel (Life Technologies) and transferred to polyvinylidene difluoride membranes using an iBlot (Life Technologies). The m9, m36, or AP-m36.4 proteins were detected using monoclonal mouse anti-FLAG antibody (Pierce Thermo) dilutes 1:1,000 and the anti-mouse Western Breeze Chemiluminescent Kit (Life Technologies).
Purification and bioactivity characterizations of Lactobacillus-derived m36.4
The L. jensenii strains expressing m9 and m36 (each containing the ABD) from plasmids were cultured in Rogosa SL broth (Difco) to stationary phase. Cell-free supernatants were centrifuged, filtered, and purified using the (i) albumin Sepharose column (RSA bound to cyanogen bromide [CNBr]-activated Sepharose 4B; GE Healthcare) or (ii) full-length single-chain (FLSC) column (gp120/CD4 complex bound to CNBr-activated Sepharose 4B). In both methods, the columns were equilibrated with phosphate-buffered saline (PBS), the supernatant supplemented with 0.5 M sodium chloride (NaCl) passed slowly through the column to ensure at least 1 min of contact time, column washed with at least 20 bed volumes PBS supplemented with 0.5 M NaCl and m36 eluted with 0.2 N citric acid, pH 3.0.
The eluant was immediately neutralized to pH 6.0–8.0 using 1 M Tris at pH 9.0 and dialyzed against PBS. Control E. coli expressed m36.4 was purified by sequential ion exchange column (Q HP Sepharose Fast Flow columns; GE Healthcare Life Sciences) and size exclusion column (Superdex 75 column, 120 ml bed volume; GE Healthcare). Periplasmic extract from HB2151 expressing m36.4 was dialyzed against 20 mM piperazine, pH 5.8, centrifuged at 14,000 g for 30 min, and loaded onto the Q HP column. The column was washed with 20 mM piperazine, 0.15 M NaCl, pH 5.8 buffer, and the bound protein eluted using gradient salt elution from 0.15 to 0.50 M NaCl. M36 and m36.4 eluted in the peak in the range of 0.2 M NaCl. The collected eluant from this peak was passed through a size exclusion column to eliminate the majority of high-molecular weight contaminants.
Preparations of m9, m36, and m36.4 were quantified by Coomassie blue staining and gp120/CD4 capture enzyme-linked immunosorbent assay (ELISA). FLSC (CD4-gp120) leached from column was removed by an additional passage over an RSA column. The amount of RSA leached from the column was insignificant.
Although LPS levels were not measured following purification from E. coli, similar antibody fragments purified previously had <10 EU/mg of protein.
Gp120/CD4 binding (capture ELISA)
A sheep antibody, D7324 (Aalto Bio Reagents), which binds to a motif (amino acid 497–511) in the envelope gene gp120, was coated on ELISA plates at 2°C–8°C, at a concentration of 2 μg/ml, overnight. Plates were then washed with Tris-buffered saline with Tween (TBST) and blocked with 5% skim milk. FLSC (a gp120/CD4 complex), at a concentration of 0.2 μg/ml in PBS, was captured on D7324 and the plate was washed with TBST. Titrated samples of m9, m36, and m36.4 solution were overlaid onto the FLSC, incubated for 1 h, and washed with TBST. All antibody fragments bound to FLSC and were detected using the horseradish peroxidase (HRP)-conjugated mouse anti-FLAG antibody (clone M2; Sigma).
Peripheral blood mononuclear cell neutralization assay (p24 assay)
Human peripheral blood mononuclear cells (PBMCs) were purchased from SeraCare Life Sciences, Inc. HIV-1BaL virus was initially received from the NIH AIDS Reagents Program and later grown by Profectus Biosciences for laboratory use. PBMCs were infected with HIV-1BaL at a multiplicity of infection of 0.01 in the presence of m36.4 (purified as described above) with desired concentrations. The mixture was incubated at 37°C for 1 week, and the supernatant was lysed using 1% Triton X-100 and quantified for p24 presence by using a p24 capture ELISA developed at Profectus BioSciences, Inc. Plates were coated with 3 μg/ml mouse monoclonal anti-p24 antibody (clone 39/6.14 by ZeptoMetrix) washed with TBST, blocked with 5% casein and 4% rabbit serum, washed with TBST, overlaid with the lysed p24 containing supernatant, incubated for 1 h, and washed with TBST. Bound p24 was detected using the HRP-conjugated rabbit polyclonal anti-p24 antibody (1303-P; ImmunoDX, LLC).
Construction of recombinant L. jensenii 1153-1128
Before insertion of the antibody cassette into the Lactobacillus chromosome, two modifications were made. The two-amino-acid peptide (AP), corresponding to the N terminus of the mature CbsA protein, was inserted downstream of the CbsA signal sequence at the N-terminus of the m36 protein to more closely resemble the native signal peptidase cleavage site of this protein. A point mutation at amino acid 44 glutamine (Q) to glutamic acid (E) Q44E was substituted for the original sequence. 19 This point mutation, also called m36.4, confers greater breadth against HIV-1. The AP-m36.4 gene was cloned into Osel's integration vector.
For stable expression of AP-m36.4 in Lactobacillus, the AP-m36.4 gene expression cassette was integrated in single copy into the L. jensenii 1153 genome by homologous recombination into the minor capsid gene. The recombinant strain was selected by replicate plating into MRS agar with and without erythromycin. During successful integration, resolution of the plasmid occurs, which removes the erythromycin resistance gene. The integrated and resolved strain L. jensenii 1153-1128 contains a single copy of AP-m36.4 gene and requires no antibiotic selection. The recombinant strain L. jensenii 1153-1128 was maintained in the MRS broth.
TZM-bl neutralization assay
The TZM-bl neutralization assay was used to evaluate antibody breadth and potency of m36.4. 29 Neutralization experiments are expressed as IC50 titer of the protein in μg/ml. m9 and m36 were not tested.
Results
Expression of antibody fragments in L. jensenii
We tested the feasibility of expressing anti-HIV-1 antibody fragments using a human vaginal Lactobacillus expression platform to produce a recombinant live biotherapeutic against HIV-1. We examined the scFv-m923,24 and single dAb-m36 (CD4i epitope) 25,26 and its derivative m36.4. 19 We cloned the m9 and m36 genes, which contained an ABD for purification 28 and FLAG tag (for easy detection) into Osel's Lactobacillus expression vector p175 under the p23 promoter and transformed L. jensenii 1153. A diagram of the plasmid construction of pOSELp23-m9 and pOSELp23-m36 is shown (Fig. 1A). Supernatants from L. jensenii secreting m9 or m36 are detected by using an anti-Flag antibody and compared with a purified protein standard produced in E. coli (Fig. 1B). The estimated concentrations of m9 and m36 secreted were ∼660 and 4.0 μg/ml, respectively.
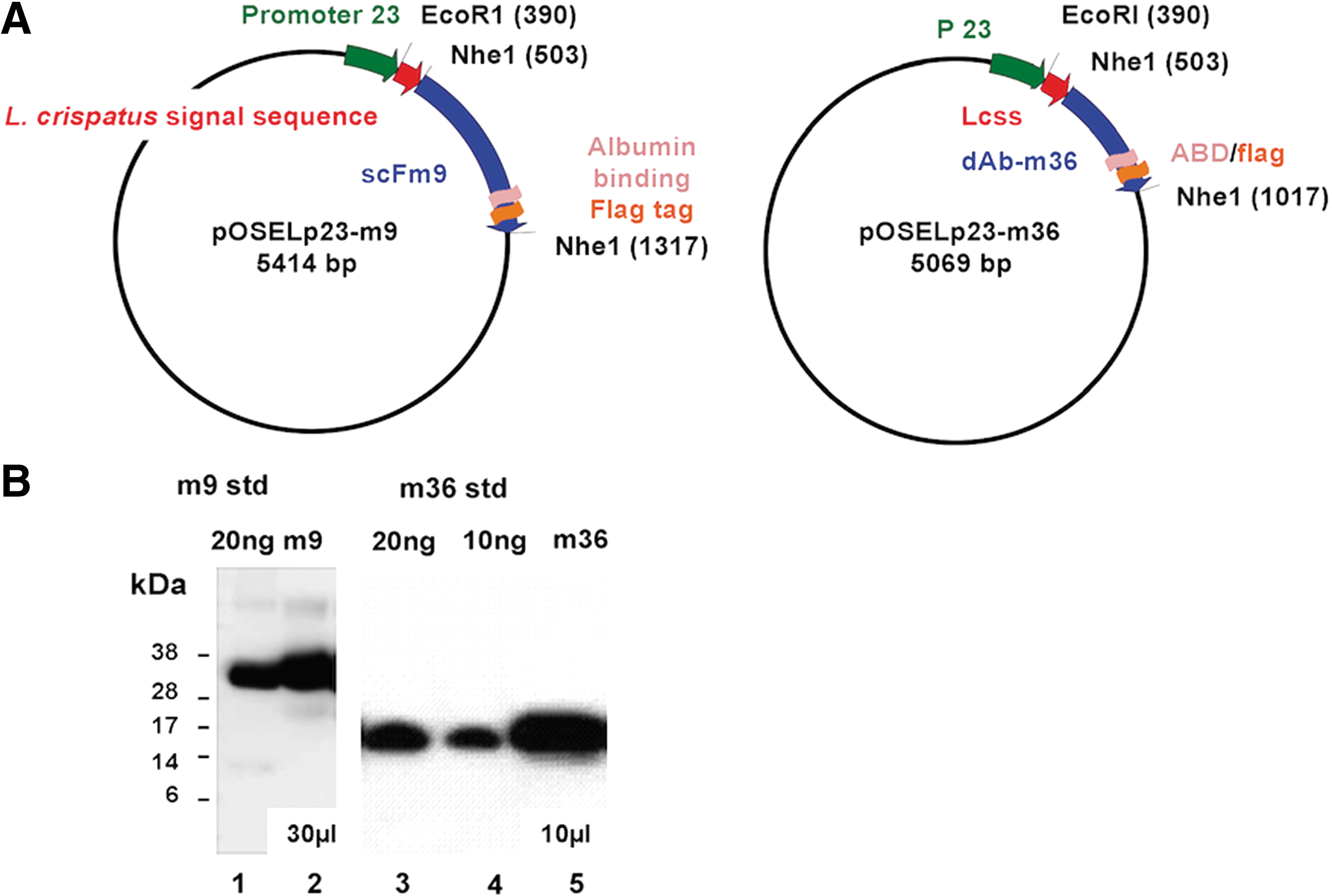
Plasmids used in this study and protein expression.
Binding of antibody fragments to FLSC
Supernatants of scFv-m9 and m36 expressed from plasmids transformed into L. jensenii 1153 were purified and tested for functionality by determining binding to FLSC (Fig. 2A, B). The scFv-m9 plasmid expressed by L. jensenii or scFv-m9 expressed by E. coli bound to the gp120-CD4 complex in ELISA, while the control supernatant from L. jensenii did not (data not shown); however, scFv-m9 expressed by L. jensenii bound with lower affinity (Fig. 2A). Due to lower expression levels and lower affinity, scFv-m9 was not pursued further.
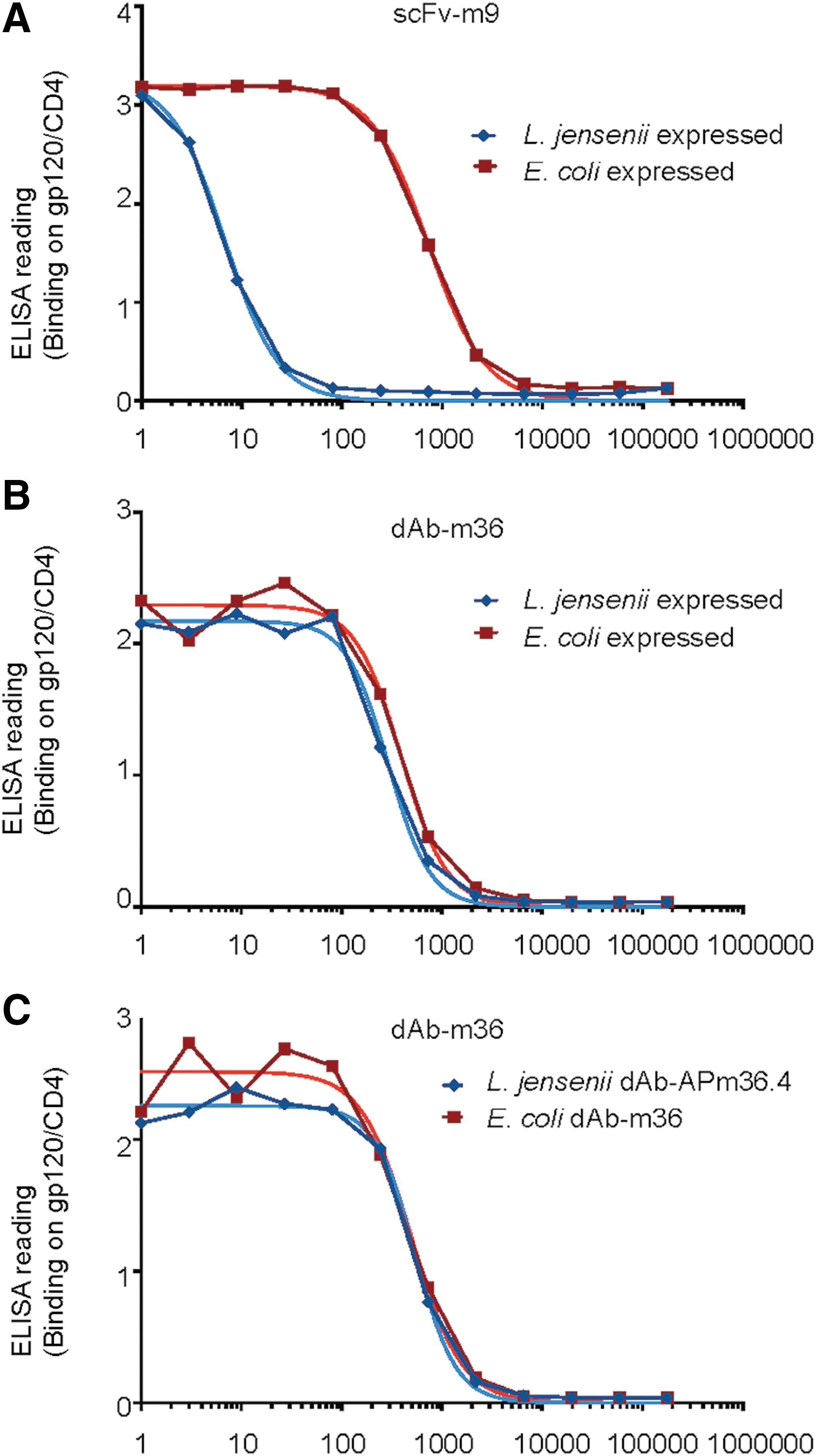
ELISA readout of binding of antibody fragments to FLSC (gp120/CD4) and purity of m36.4 and AP-m36.4. Comparison of binding properties from antibody fragments produced by L. jensenii (blue) to Escherichia coli produced standards (red).
L. jensenii plasmid-expressed m36 bound to FLSC to the same level as the standard (Fig. 2B). The recombinant L. jensenii 1153-1128 expressed a modified version of m36.4, with the addition of amino acids alanine (A) and proline (P) at the amino terminus. Addition of the AP-sequence was previously shown to improve the secretion of full-length protein from L. jensenii. 30 Both E. coli-expressed m36.4 and recombinant L. jensenii 1153-1128-expressed AP-m36.4 were tested to confirm that the addition of AP to the amino terminus of m36.4 did not influence binding to FLSC (Fig. 2C). The EC50s were comparable: E. coli produced dAb-m36.4 12.7 ng/ml and recombinant L. jensenii 1153-1128 produced AP-m36.4 11.9 ng/ml (no statistical difference).
Antiviral activity of m36.4
L. jensenii-expressed m36.4 was purified using two techniques; purity of the protein using the different strategies is shown in Figure 3A. Purified m36.4 was tested for anti-HIV activity using a p24-PBMC-based assay (Fig. 3B). Both RSA-purified m36.4 and FLSC-purified m36.4 showed similar reduction of p24, and thus, the Lactobacillus-produced m36.4 protein was active against HIVBaL with an IC50 of ∼1 μg/ml.
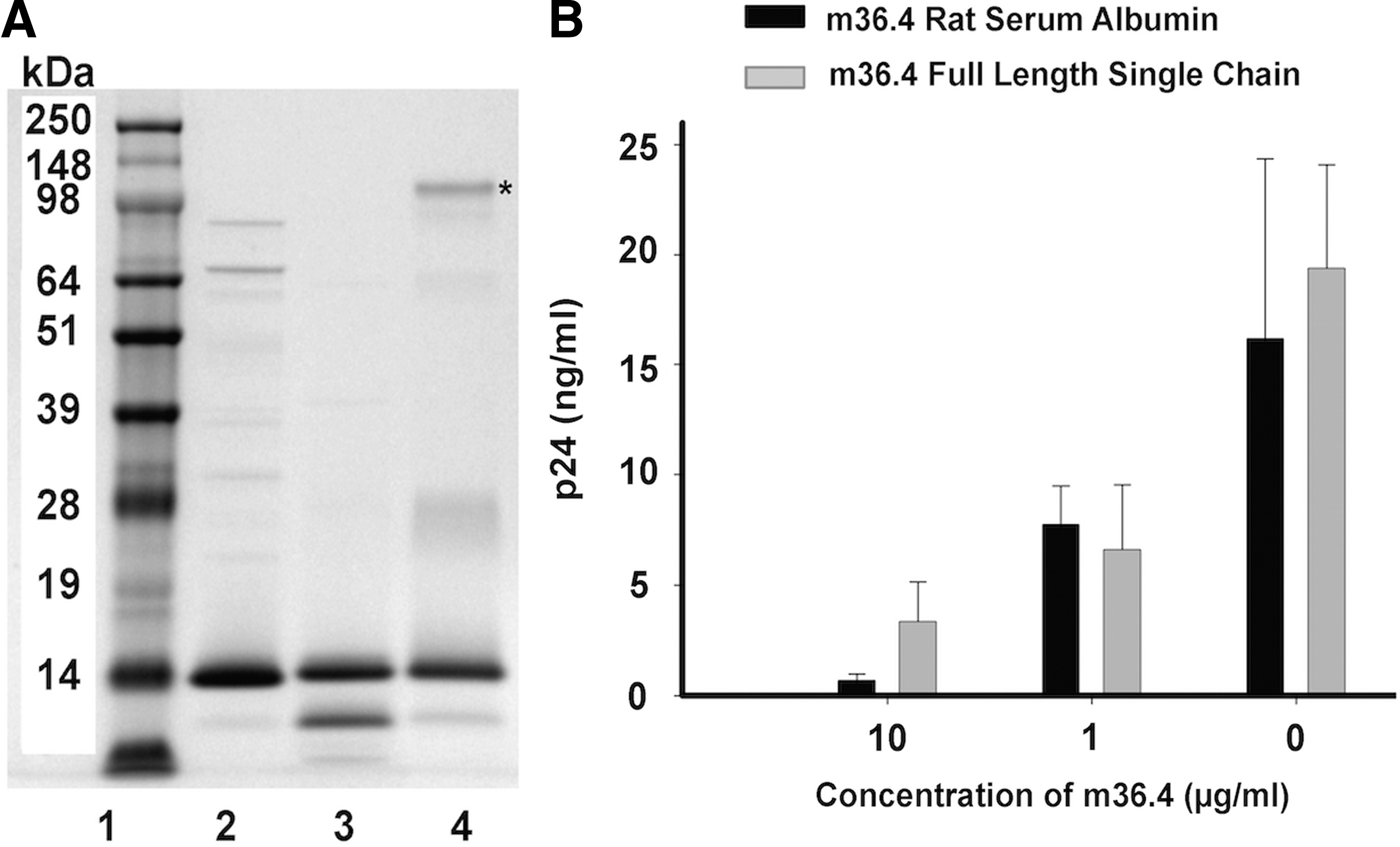
Purification of AP-m36.4 protein used for HIV inhibition assay.
Neutralization breadth of m36.4
Table 1 details the breadth of neutralization of the m36.4 protein. m36.4 was purified from E. coli and tested for activity against a panel of 68 pseudoviruses from each of the major genetic subtypes representing clades A, B, C, D, G, including transmitted founders (T/F) and circulating recombinant forms CRF01 AE and CRF02 AG. Table 1 shows the broadly neutralizing activity of m36.4 against clades B, C, and D; most IC50 values were in the ≤5 μg/ml. Less or no activity was noted against clades A, G, and CRFs. Previously, we tested a partially purified supernatant from wild-type L. jensenii in pseudovirus assays with HIV-1HxB2 envelope and found no inhibition. 27 In addition, m36.4 did not inhibit MuLV control in the TZM-bl assay.
T/F, transmitted founders.
Construct integration vectors and integrated single domain expression cassettes into the L. jensenii 1153 chromosome and tested phenotypic stability
We constructed a recombinant L. jensenii 1153 by integrating an optimized AP-m36.4 antibody expression cassette into the minor capsid gene of L. jensenii 1153 through homologous recombination (Fig. 4A). The strains generated were evaluated for complete resolution of the plasmid and antibody production (not shown). A diagram of the two sets of internal and external primers, which were used to amplify the integration region within the minor capsid gene, is shown (Fig. 4B).
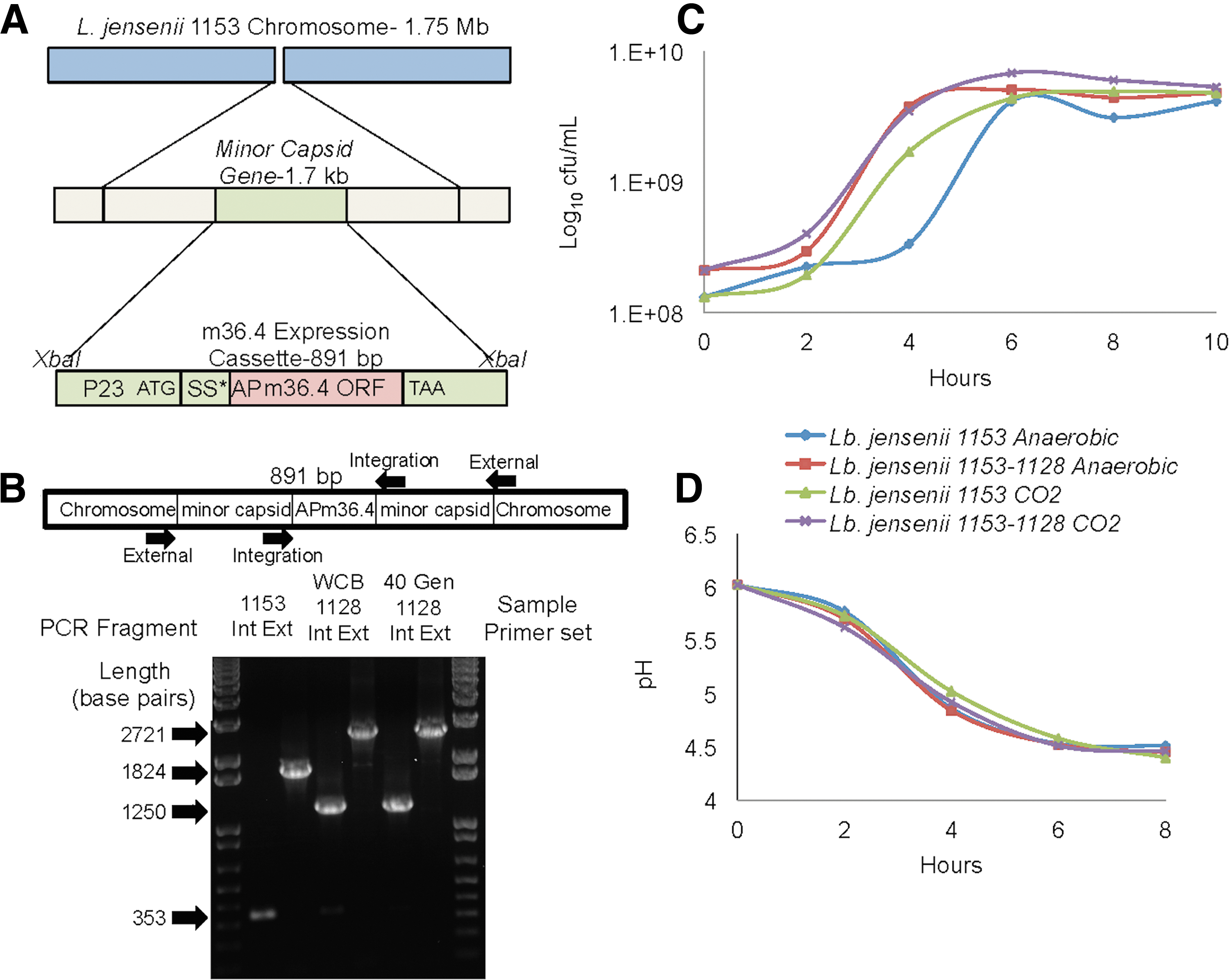
Construction and characterization of recombinant Lactobacillus.
Polymerase chain reaction (PCR)-amplified genomic DNA from the parental L. jensenii 1153 and recombinant strain designated L. jensenii 1153-1128 were run on a 2% agarose gel. L. jensenii 1153 shows a 353 bp band corresponding to a region of the minor capsid gene and a 1,824 bp band of the entire minor capsid gene (Fig. 4B). L. jensenii 1153-1128 shows a 1,250 bp band corresponding to a region of the minor capsid gene with the 891 bp band (+6 bp restriction site) cassette insertion. The external primers amplify a 2,721 bp band (1,824 + 891 + 6 bp).
The strain was passaged 40 generations, genomic DNA was extracted, and the cassette was sequenced (not shown). The 40-generation sample was also analyzed by PCR for stability. No changes were noted in the size of the internal and external primer bands 891 and 2,721, respectively, and thus, the L. jensenii 1153-1128 recombinant is genotypically stable. We examined phenotypic stability by performing a carbohydrate utilization profile of parental strain L. jensenii 1153. Both L. jensenii 1153 and L. jensenii 1153-1128 fermented glucose,
We tested growth of the 1153-1128 strain and lactic acid production (pH) and compared them to parental strain L. jensenii 1153. L. jensenii 1153-1128 grew to the same level as the parental strain and reduced pH of the media under aerobic (5% CO2) and anaerobic growth conditions (Fig. 4C, D).
Discussion
There is a tremendous need for approaches to combat HIV-1 transmission in women of reproductive age. Lactobacilli (including L. jensenii) are important members of the vaginal microbiome, lowering vaginal pH, 31,32 modulating cervicovaginal inflammation, 9 and providing resistance to HIV infection. 33 We established the feasibility of expressing bioactive anti-HIV-1 antibody fragments from a vaginal strain of L. jensenii and show that these fragments are functional and can be stably expressed from a recombinant L. jensenii. In addition, we confirmed that the recombinant L. jensenii 1153-1128 expressing m36.4 could colonize the rhesus macaque vagina (data not shown).
ScFv-m9 was expressed at lower levels and had lower affinity for FLSC, so it was not pursued. However, the smaller dAbs m36 and m36.4 could be expressed at high levels and thus were more viable for this approach. It is noteworthy to mention that the small dAb fragments m36 and m36.4 can bind the CD4i epitope even without the conformational change that occurs with CD4 binding to gp120.
Previously, several groups have used other commensals to express functional antibody fragments. Early work used the commensal Streptococcus gordonii engineered to produce scFv H6 and an anti-idiotypic scFv with anti-Candida activity. 34,35 S. gordonii was also used to produce Guy's 13 scFv against the caries-producing bacterium Streptococcus mutans. 36
Marcotte et al. expressed scFv on the surface of Lactobacillus paracasei against Porphyromonas gingivalis, 37 and Kruger et al. used Lactobacillus casei expressing an scFv against S. mutans, 38 both approaches reduced dental caries. L. paracasei was used to express an scFv to protect against anthrax edema toxin caused by the bacterium Bacillus anthracis. 39 L. casei, L. paracasei, and Lactobacillus rhamnosus GG were engineered to express llama VHH antibody fragments against rotavirus. 40,41 Most recently, Andersen et al. described the use of single dAbs to neutralize Clostridium difficile toxin B. 42
Another noteworthy approach is the use of adeno-associated virus (AAV) to deliver broadly neutralizing antibodies. 43 While the approach appears to be safe, and the AAV are long lived in the tissue and can secrete antibody persistently, one caveat with this approach is the inability to remove the AAV vector easily, should the need arise. Commensal bacteria (expressing antibodies) can be removed by treatment with an appropriate antibiotic.
Although the idea of expressing antibody fragments in commensals is not new, with the increasing development of antibiotic and antiviral resistance and therefore fewer treatment options for both bacterial and viral infections, new approaches are needed. With a better appreciation of the health benefits provided by commensals as part of the microbiome, perhaps this is an approach whose time has come.
For prevention of HIV, the benefit to using human colonizing vaginal strains would allow for delivery of antibodies mucosally at the site where the virus first enters the body. Development of broadly neutralizing antibodies in infected individuals, in the blood or at mucosal sites, takes months to years. Current vaccine strategies do not uniformly generate these antibodies. Our results indicate that stable expression of anti-HIV-1 antibody fragments is possible using a common Lactobacillus in the vaginal microbiota as a recombinant live biotherapeutic. m36.4 was used as a prototype antibody fragment, but other newly discovered broadly neutralizing antibodies might also be candidates to test in this approach.
Footnotes
Acknowledgments
We thank the Michael Seamen Laboratory at Harvard Medical School for performing the TZM-bl pseudovirus assays. This work was funded, in part, by the Bill and Melinda Gates Foundation GCE Phase 1 and 2 OPP1035271 and NIH grant 5R44AI100753-03. This work was supported by the National Institutes of Health, SBIR grant 1R43AI100753-01.
Author Disclosure Statement
L.A.L., P.P.L., and T.P.P. work for Osel, a biotherapeutics company that is developing L. jensenii as a delivery system for anti-HIV therapeutics, and are shareholders. A.M. works for Osel. X.L. and L.J. previously worked for Osel. X.L. is a shareholder of Osel, Inc. A.S.D. and W.Z. declare no conflicts. T.R.F. is a founder and shareholder of Profectus Biosciences, Inc.