
Abstract
In literary texts, the visual arts and the popular imagination, ‘madness’ and ‘chaos’ are indelibly intertwined. For psychiatric neuroscientists, the cognitive disorganization of schizophrenia and the unstable fluctuations of mood in bipolar psychoses intuitively appeal to the concept that a chaos may underlie brain dysfunction in the major psychoses. Modern time series techniques allow empirical testing of this intuition and a number of studies have appeared in the scientific literature. In this paper, the progress of scientific research in the study of nonlinear dynamics (‘chaos theory’) and schizophrenia is reviewed and an outline of a ‘nonlinear theory of schizophrenia’ is proposed.
Several considerations motivate a study of nonlinear brain dynamics in schizophrenia. First, because of the nature of ‘excitable membranes’, the basic dynamical properties of neural systems are nonlinear [1]. One of these – the relationship between cell membrane potential and its rate of firing – is depicted in Fig. 1. Any abnormality in neural architecture and physiology associated with schizophrenia – and many have been proposed – may impact directly or indirectly on the basic nonlinear properties of brain systems. Changes in neuropil density or glutamate receptor properties, for example, affect the nonlinear response of local neural population firing rates to afferent excitation. Computer models of such processes have been shown to lead to abnormal dynamics and simulation of ‘hallucinations’ [2]. Hence, nonlinear considerations should be among the primary ‘armaments’ of any theorists of brain dysfunction. Second, schizophrenia symptoms such as formal thought disorder – where the sequence or flow of thinking is disrupted – are dynamic in nature, suggesting a disruption in nonlinear processes such as state transitions in cortical systems [3]. Third, the irregular fluctuation of symptom severity during acute psychosis suggests a dysregulation of the nonlinear dynamics of neuronal systems [4]. Finally, if schizophrenia is viewed as a dysfunction in the dynamical regulation of the self [5], then its psychosis may be conceptualized as a failure of the stability, self-regulation and hierarchical ordering of nonlinear brain systems.
The nonlinear relationship between the average membrane potential and the mean rate of firing in a small neural ensemble obeys a nonlinear sigmoid relationship. This is because in a population of heterogeneous neurons, the threshold potential at which individual neurons fire is (Normally) distributed. According to a variety of factors, this nonlinearity may be weak (a.) or strong (b.) – which may in turn have a profound impact on the behaviour of the neural population. The dashed line shows a linear relationship: In some circumstances mathematical modeling may employ a linear (approximate) approach.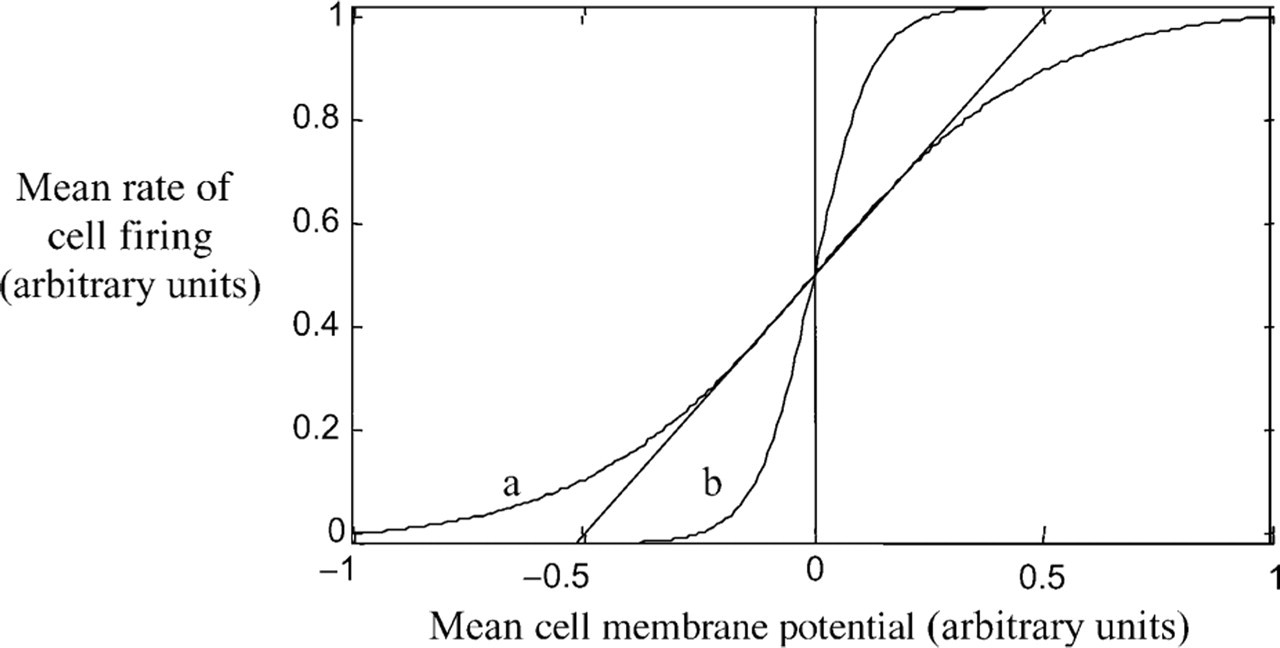
Basic neuronal systems and large-scale brain processes, however, differ from each other by several orders of magnitude. In addition, the relationship between neural activity and mental phenomena is a far-from resolved problem. Hence, the role of chaos theory in understanding the psychoses is an empirical question, which requires careful scientific research. This research is reviewed in this paper, which is structured as follows. In the next section, the development of the conceptual and experimental tools required for the nonlinear analysis of experimental data is presented. More technical summaries can be found in [6, 7]. In section ‘Initial studies of schizophrenia using nonlinear techniques’, the application of thesemethods to time series data from healthy and schizophrenia patients is reviewed. Section ‘Flaws in the initial application of nonlinear methods to human EEG’ discusses the major limitations of these algorithms. The major advance in this regard has been the development of ‘surrogate data’ techniques, which permit the statistical re-appraisal of nonlinearity in brain dynamics. This is discussed in section ‘Statistical hypothesis testing in nonlinear studies’. The study of ‘brain chaos’ in the spatial domain is examined in section ‘Brain chaos: spatial aspects’. The paper concludes with an appraisal of the overall impact of nonlinear methods in current neuroscience and psychiatry research.
Principles of measuring nonlinearity in experimental data
Dynamical systems theory has a long and successful history, underpinning diverse sciences such as celestial mechanics, fluid dynamics and quantum physics. Isaac Newton's Time series and phase space representations of a model neural system consisting of three dynamical variables: V – pyramidal cell membrane potential, Z – inhibitory cell potential, and W – potassium ion conductance (from [9]). Top row shows a brief time series (a) and corresponding phase space orbit (b) of the system. The arrow depicts the direction of flow on the orbit starting from a random initial condition (diamond). The bottom row depicts a longer time series (c) and phase space attractor (d) of the system in a chaotic state. The attractor corresponds to the long-term evolution of the system's orbits.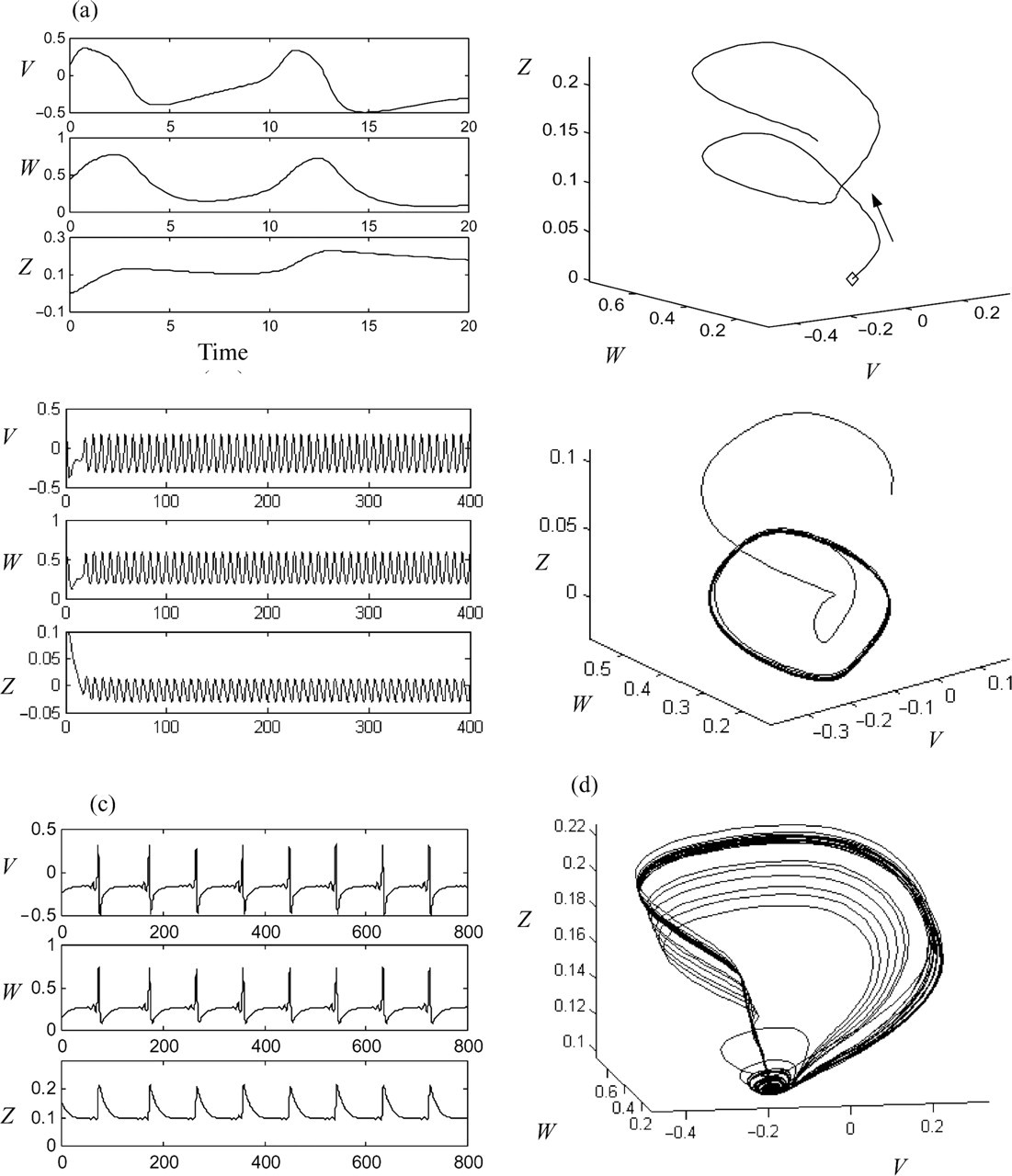
Two important quantities are essential to a proper understanding of attractors. They are termed
‘Exponents’ embody the nature of a system's dynamics. ‘Dimensions’ capture the degrees of freedom of such dynamics. Although these properties both contribute to the behaviour of a system, exponents are most important to the stability and predictability of the system, whereas the system's dimension reflect the minimum number of variables required to capture the system's evolution.
The Lyapunov exponents capture the nature of a dynamical system. Here, two orbits beginning at points 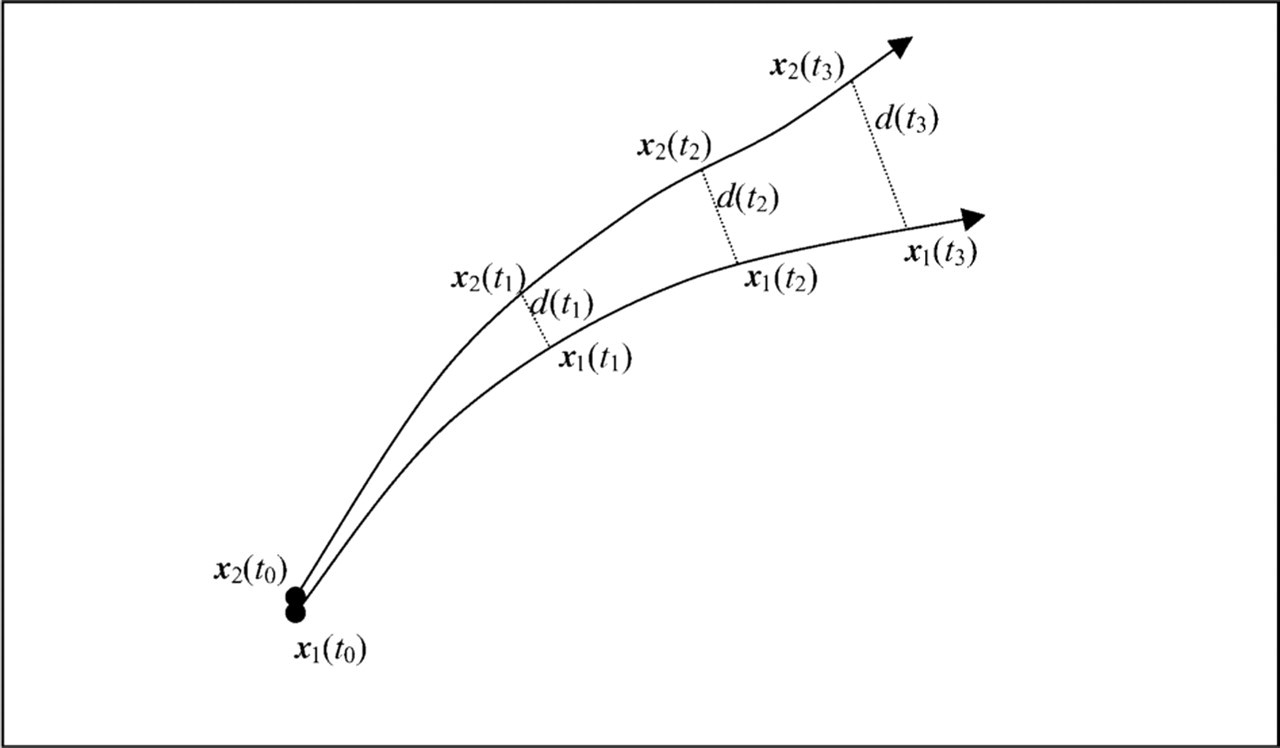
In an experimental setting, a dynamical system such as the brain yields (a) Through experimental measurement of a system of interest (here a graphically illustrated brain region) a time series data set (b) is captured (in this case a Petit Mal seizure). (c) Time delay-embedding provides a reconstruction of the complete attractor, which can then be subject to nonlinear analysis, yielding ‘exponents’ and ‘dimensions’.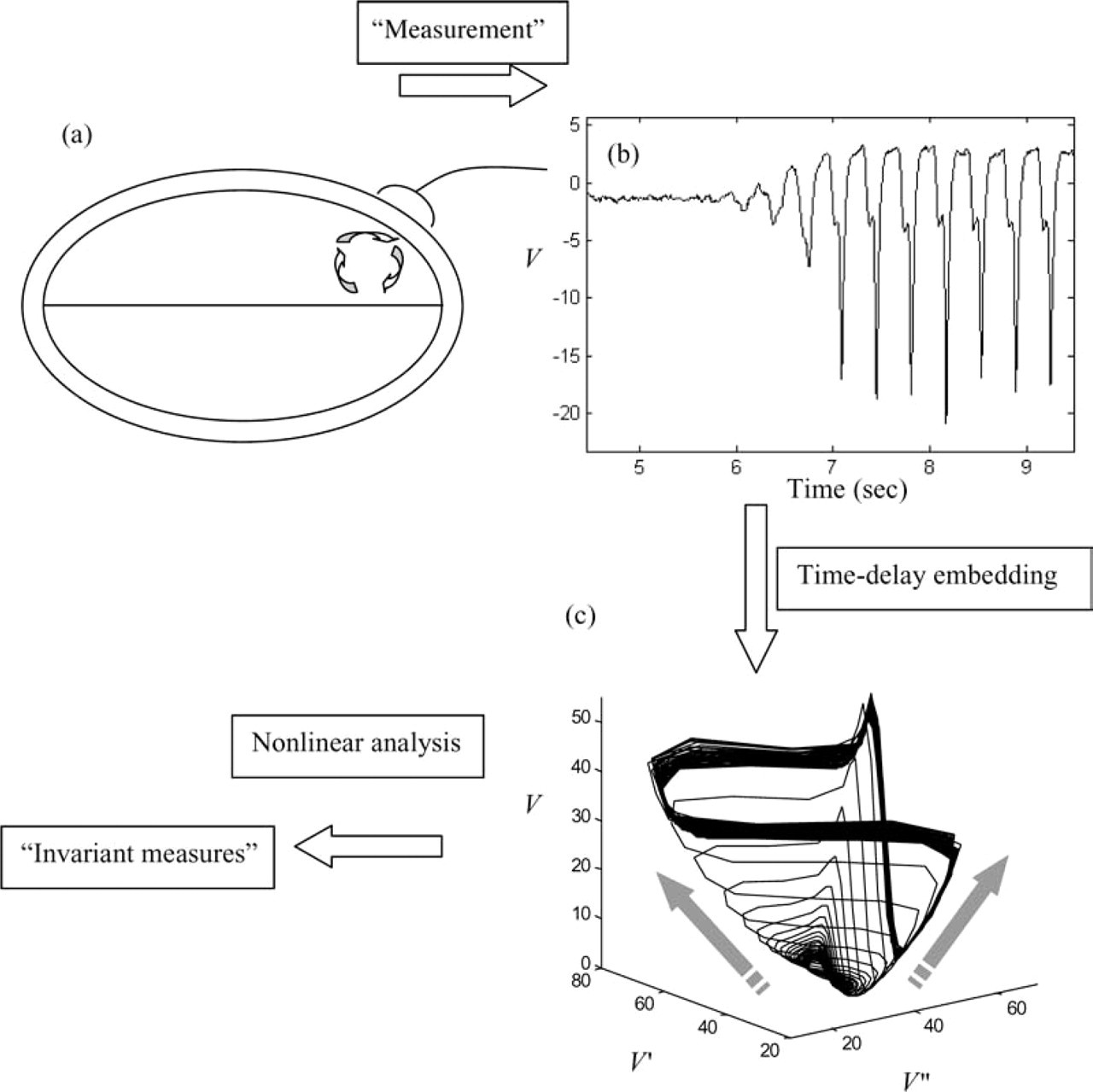
Thus,
Initial studies of schizophrenia using nonlinear techniques
Application of nonlinear methods to EEG data
Until the 1980s, linear methods of time series analysis prevailed in neuroscience. For example, the analysis of sleep EEG relied on spectral and coherence measures, both linear techniques [19]. Advances in nonlinear theory initiated the widespread use of nonlinear methods in the 1980s for the first time. Their application to the EEG was motivated by a re-appraisal of the assumption that the irregular nature of EEG data must be because of a stochastic process. The aim was to determine whether an orbit reconstructed from EEG data bore the characteristic fingerprints of a strange attractor as outlined above – a non-integer correlation dimension and a positive principal Lyapunov exponent. There soon appeared a series of reports of positive findings. The normal human EEG was reported to exhibit chaotic dynamics in waking states [20–24] and in the alpha rhythm [25–27]. The correlation dimension was shown to change with different sleep stages [28–30] and during the performance of various cognitive tasks [31–33]. For example, the performance of mental arithmeticwas shown to correlate with an increase in D2 calculated from parieto-occipital EEG recordings [34]. Chaotic activity during an epileptic seizure was also reported, with a D2 significantly lower than during normal brain activity [35].
The proposal that the EEG contained evidence of lowdimensional chaos generated widespread interest and gave impetus to conjectures about the computational significance for cognitive processes [36, 37]. Chaotic systems act as sources of information and complexity: This was conjectured to underlie the dynamical state changes in different stages of awareness and with changes in cognition [34].
Nonlinear analysis of EEG differences in schizophrenia
The enthusiastic embrace of this new dynamical paradigm quickly lead to speculation on the significance of brain chaos for psychiatric diseases [38] and motivated the application of the Grassberger–Procaccia and other algorithms to EEG data collected from patientswith schizophrenia.1 Elbert
The disturbances of the normal sleep EEG architecture associated with schizophrenia were also investigated from a nonlinear perspective. Using the Grassberger–Procaccia algorithm with 13 patients with schizophrenia (neuroleptic free) and 13 matched healthy controls, Roschke and Aldenhoff [44] found a decreased D2 during stage II and REM sleep in parietal EEG of the clinical patients. Roschke
The impaired cognitive performance of patients with schizophrenia has also been studied using nonlinear methods. Kirsch
Two experiments reported evidence for abnormal nonlinear dynamics in behavioural observations of patients with schizophrenia. Paulus
In summary, the initial application of nonlinear methods to EEG and behavioural data from patients with schizophrenia showed abnormal patterns of complexity and irregularity in comparison to healthy patients. Overall, both the behaviours and electrophysiology in the clinical patients exhibited a loss of complexity and, most importantly, a loss of the normal flexibility. Taken literally, this could be interpreted as evidence that in schizophrenia, abnormal neural dynamics compromise a balance between flexibility and stability that may be critical to normal brain function. This, in turn, could result from a variety of pathological processes impacting on basic neurophysiological function.
Flaws in the initial application of nonlinear methods to human EEG
Unfortunately, themethodology used to study nonlinear structure in human EEG contained several major flaws. Some researchers now view the whole paradigm with considerable scepticism. Although caution is well justified, recent research has addressed many of the difficulties, facilitating further progress in this field. The main theoretical, methodological and conceptual flaws are now discussed.
Theoretical flaws
First, it is erroneous to attribute a non-integer correlation dimension directly to chaotic behaviour. Many of the studies reviewed above do make this mistaken assumption. Chaos is defined in terms of sensitive dependant dynamics and, therefore, a positive Lyapunov exponent. It is true that
Second, there is a problem with interpreting the significance of a nonlinear index in the absence of a welldeveloped nonlinear model of the brain at the scale of experimental measurement. By a model, a set of equations together with a relation between the solutions of this model (e.g. neural firing rates) and the experimental measurements (e.g. scalp potential) is implied. Although large-scale neural models have also recently been elucidated [51–58], in general, they have not been used in conjunction with nonlinear experimental studies. Moreover, because of their complexity, most of the analysis of these models has been restricted to a linear approximation of their behaviour. The interpretation of nonlinear measures of EEG waveform in the context of a nonlinear brain model would strengthen their significance.
Methodological flaws
The methodological flaws allow for spurious detection of nonlinearity in time series data generated by linear, stochastic processes. Stochastic noise, by definition, has an infinite dimension, but it can mimic chaotic behaviour through the ‘looking glass’ of dimensions and exponents derived from finite-length time series.
Ruelle [59] showed that the length of the analysed time series, whatever its nature, puts an upper bound on the correlation dimension that can be inferred. This is important because itwas previously assumed that the difference between an irregular nonlinear signal and a (linear) noisy signal was that the correlation dimension would plateau for the former, but not for the latter. Ruelle showed this was not true. Thus, a small non-integer D2 can be derived from an infinite dimensional stochastic process sampled for a finite length of time [60]. This flaw has possibly contributed to erroneous detection of low-dimensional chaos in papers well received at their time of publication [61] and to the wide variety of dimension estimates for the EEG that have been reported in the literature. Almost invariably such studies used data of a variable and inadequate sample length and do question the validity of the reports of lower D2 in schizophrenia.
Osborne and Provencale [62] showed that
Estimates of Lyapunov exponents face similar problems. Specifically, it has been shown that Lyapunov exponents calculated from stochastic signals are generally positive and take a value that depends upon computational parameters used in the reconstruction algorithm, not the nature of the signal [66]. In other words, a positive principal exponent is not, by itself, evidence of chaos.
Conceptual flaws
Three conceptual limitations are inherent in the above research. First, critical aspects of neural connectivity are not reflected by the design of the analysis. The brain is characterized by local dense interconnectivity and sparse long-range excitatory projections. This permits both local (specialized) and global (integrative) network function [67, 68]. For example, cortical macrocolumns are to some degree functionally specific, but they must operate in synchrony with others to achieve integrated processing of complex perceptual objects. Nonlinear algorithms are derived from the analysis of
Second, mathematical derivations of the invariant measures rely on the properties of attractors. These are reached by systems with static state parameters after initial transients have died away. However, the dichotomy between (static) state parameters and (rapidly evolving) independent variables seems unsuited to a study of the brain, where different physiological systems evolve and interact across a wide spectrum of temporal scales. Thus, it may be more appropriate to conceptualize multiple dynamical subsystems interacting across different spatial and temporal scales [71]. Such systems are typically observed to switch erratically between different types of behaviour. ‘Transient chaos’, ‘attractor ruins’ and chaotic ‘itinerancy’, rather than long-term chaotic or periodic dynamics, are seen [72, 73]; as the names suggest, each of these terms capture transient behaviours. These occur either because state parameters are changing such that the ‘attractor landscape’ of the system is constantly in flux or the landscape consists of several competing potential attractors; the system switches unpredictably between each attractor. Theorems concerning invariant measures are not valid in such situations. Alternative techniques such as the study of ‘dynamical archetypes’ [74] may be more appropriate.
Third, the relationship between observed data and the systems from which they arose is often given insufficient thought. Scalp and surface cortical EEG data nicely capture the fine timescales of many neural processes, but their actual neural origins and the influence of electrical propagation from surrounding tissues (‘volume conduction’) remains poorly understood [75]. Until the science of such ‘observation’ processes is better elucidated, the conceptual relationship between the data and the brain must be understood cautiously. As with the theoretical flaws, models of large-scale neural activity may provide clarification of this area [76].
Statistical hypothesis testing in nonlinear studies
The recognition of these flaws by the scientific community had a significant impact on research in this field. Although the initial studies led to a growth of interest in the nonlinear properties of the brain, they failed to adequately generate explicit hypotheses that could potentially be rejected by statistical testing. The ‘verification’ of chaos in EEG data is quite different from refuting alternative null (linear) hypotheses. This problem occurred in other areas where chaos theory had been applied, including the analysis of financial trends, animal populations and weather patterns; nonlinear science lacked a suitable statistical framework.
Surrogate data: the development of statistical hypothesis testing
A major advance in this regard was achieved with the development of the method of ‘surrogate data’ [77, 78]. This technique addresses the preceding methodological flaws by controlling for the imperfect nature of the experimental data. The principle is to construct an ‘ensemble’ of datasets which have the same linear properties as the original data, but have had any nonlinear structure destroyed. The surrogate data represent the null hypothesis that the data have been produced by a stochastic process. The null hypothesis can be tested by a statistical comparison of the nonlinear ‘invariant’ calculated from the original data and the distribution of values calculated from the surrogate data. Rejecting the null hypothesis that the time series is linear supports the alternative hypothesis – that it has nonlinear structure – in a more robust manner than previous confirmatory studies could provide. The most widely used surrogate technique relies on randomizing the phase of the time series in discrete frequency bins [78].3 Improvements to this process, optimizing the linear match between the original data and the surrogates [79, 80], or permitting analysis of multivariate datasets [70, 81], data with an asymmetric amplitude distribution [82] or strong periodic components [83] have recently been introduced.
Estimates of dimension and exponents have generally been superseded by other measures sensitive to nonlinearity such as nonlinear forecasting [84–86] and the detection of temporal asymmetry [83]. The focus of research has shifted from the explication of chaos to the more modest aims of showing the presence of nonlinearity, which is necessary for chaos but does not guarantee that it occurs. In a spatially extended system, such as the brain, chaos may only emerge when there is large-scale synchronization of neurons so that the number of degrees of freedom are sufficiently constrained to a low-dimensional manifold (such as the attractor of Fig. 2d).
Re-appraisal of previous findings using surrogate data testing
The development of this technique permits a thorough statistical analysis of nonlinearity in human EEG and a re-examination of previous findings. Rombouts
The contribution of nonlinear dynamics to the generation of grossly abnormal EEG rhythms in neurological illness has also been studied using surrogate data comparison. The periodic complexes associated with cerebral infarctions, herpes encephalitis, seizures and Creutzfeldt–Jakob disease have nonlinear contributions [94, 95]. Whereas some forms of pathological delta rhythms (frontal intermittent rhythmic delta activity) are consistent with nonlinearity, others (polymorphic delta activity) could not be distinguished from their (linear) surrogate counterparts [96]. It was concluded that the nonlinear complexes resulted from disturbed neuropil architecture, steepening the sigmoid membrane-potential/ cell firing relationship. Nonlinearity has also been detected in the EEG of patients with Alzheimer's dementia [97–99]. Interestingly, the combination of linear and nonlinear analysis obtained a more accurate differentiation between normal and dementia EEG than either method alone [100].
Statistical testing of the nonlinear schizophrenia hypotheses
The combined use of nonlinear methods and surrogatebased hypothesis testing has thus shown the potential to speak to brain dysfunction in a variety of diseases. However, to our knowledge,4 there has appeared very few studies of nonlinearity in the EEG of schizophrenia using the surrogate data technique. Lee
At least two studies have used surrogate data techniques to investigate the dynamics of the
Nonlinearity in the symptoms of psychosis and mood disorders supports the hypothesis for abnormal nonlinear brain dynamics in these disorders [4], but is not direct evidence. The timescales for psychiatric symptoms (hours to days) and brain dynamics as assessed by EEG and fMRI (milliseconds to minutes) are different by several orders of magnitude. It is possible that these fluctuating symptoms reflect nonlinear dynamics in subcortical structures operating over the appropriate timescale.
In summary, the use of surrogate data is essential in establishing the validity of nonlinear measures of brain dynamics. Studies using this technique have been used to re-examine the evidence for nonlinear dynamics in the EEG. They have consistently illustrated strong evidence for weak and fluctuating nonlinearity, but not low-dimensional chaos. There are, however, pitfalls associated with this technique, which demand that caution is still required when interpreting their significance. Nonlinearity has been illustrated in the abnormal EEG rhythms of severe neurological illness, the symptoms of schizophrenia and bipolar disorder. One study has reported evidence of nonlinearity in schizophrenia EEG. The
Brain chaos: spatial aspects
The focus of much nonlinear research has been the analysis of EEG recordings because these provide the high temporal resolution that is required for most nonlinear algorithms. Unfortunately, the spatial resolution of EEG is poor and affected by technical problems such as volume conduction. Much current research on brain dysfunction in schizophrenia uses functional brain-imaging technologies, such as fMRI, which provide both spatial and temporal information. What is the relevance of nonlinear theory to this research?
Spatial chaos: theoretical considerations
There are many complex steps between neural firing, regional blood oxygen levels (BOLD) and the computerized reconstruction of fMRI images. In fact, the relationship between neural activity, evoked haemodynamic response and the BOLD signal involves several nonlinear processes [105, 106]. This is a potential barrier to disentangling the significance of nonlinear analysis of fMRI. However, preliminary work has been successful at placing functional brain imaging in the context of nonlinear dynamical theory. Friston
Coupled chaos: a new direction in nonlinear neuroscience
An alternative method of studying spatial aspects of brain chaos is afforded by modelling the brain as an ensemble of coupled nonlinear neural systems [6, 108]. Each ‘system’ represents a large-scale cortical region with nonlinear properties, which are hierarchically coupled together by long-range pyramidal cell connections as depicted schematically in Fig. 5. This principle captures the important ‘modularity’ of the cerebral cortex, which has been widely used as a conceptual basis for neuroanatomy [109, 110]. This approach can be tested by examining EEG data for evidence of nonlinear interdependencies (a) Some modern theories of brain dynamics model the cerebral cortex as a system of coupled nonlinear oscillators. Local network activity is facilitated by dense short range neuronal connections between excitatory and inhibitory neurons. Long-range intra- and inter-hemispheric connections between such local subsystems are purely excitatory. (b) Analysis then proceeds by investigating interdependences between the time series of the local oscillators.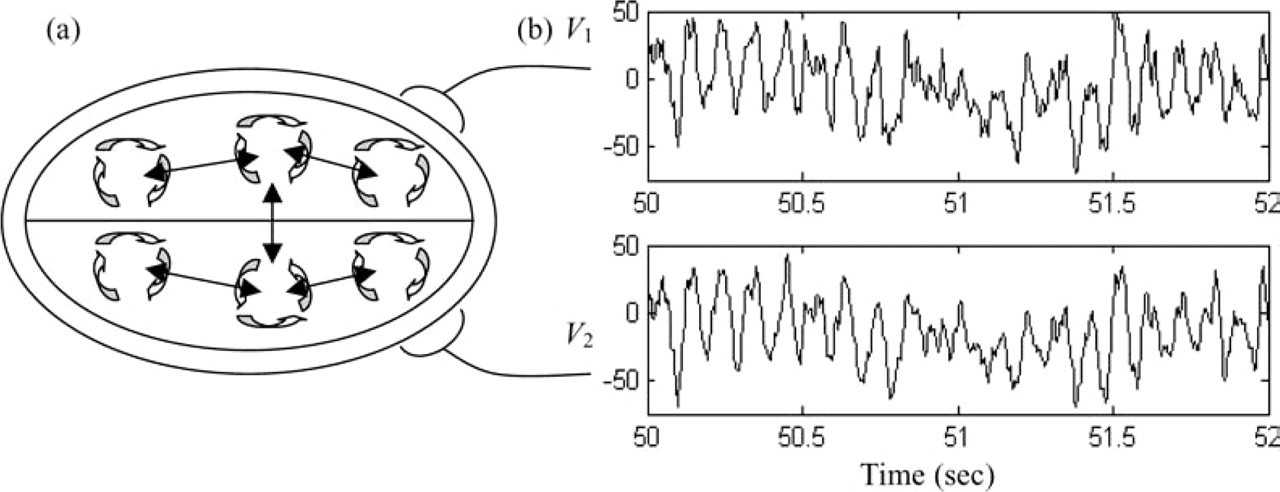
There have recently been several important advances in the study of coupled chaotic attractors that facilitate such an approach. The first advance was the recognition that two or more chaotic attractors can quickly synchronize when permitted to interact [111]. Given the sensitive dependence and irregularity of chaos, this is somewhat surprising. Depending on the type of coupling, chaotic systems are capable of different types of synchronization. With
The existence and type of chaotic synchrony depends upon a balance between local nonlinear behaviour and the type of intersystem coupling [116]. These considerations are strongly plausible for an understanding of large-scale brain dynamicswhere local neural nonlinearity and largescale excitatory coupling are each modulated by separate neuropharmacological mechanisms [117]. Monoaminergic neurons, for example, modulate effective long-range synaptic strength through regulation of receptor density. Quite distinct mechanisms, such as excitatory feedforward, modify local neural response properties. Hence, the study of nonlinear interactions in EEG appeals as a suitable adjunct to the study of nonlinear structure in single time series and multivariate approaches grounded in linear theory such as principle components analysis.
Experimental evidence of nonlinear cortical interdependence
Two preliminary investigations [118, 119] using intracranial EEG recordings in human patients with epilepsy have tested for nonlinear interdependence using multivariate surrogate data. Both reported strong evidence of nonlinear coupling. The interdependencewas strongest between the focus of epileptic activity and other brain regions, and increased for up to 30 seconds before a seizure. In EEG data from healthy patients, it appears that a strong nonlinear contribution to EEG coherence is only occasionally present [120, 121]. Nonetheless, it is possible that large-scale nonlinear cortical interdependence plays a crucial role in the adaptive brain, such as during phase transitions [122], the perception of novel sensory stimuli [123] or the construction of a global ‘cognitive workspace’ [124].
The tendency of antipsychotic medication to have affinity formonoamine (serotonin and dopamine) neuroreceptors motivates the hypothesis that schizophrenia may be associated with disturbed patterns of cortical interdependence [125–127]. This is because the monoamine systems project diffusively throughout the cortex and modulate the effective synaptic connectivity of corticocortical projections. Peled [128] has formulated this hypothesis as a disorganization in the way distributed cortical regions constrain and mutually shape their dynamic behaviours. This ‘disconnection hypothesis’ of schizophrenia was recently tested in a study of EEG data from patients with a recent diagnosis of schizophrenia [129]. This required analysing how often nonlinear interdependence was expressed between cortical regions – as reflected in the overlying EEG data – and how this was organized across an array of scalp sites. The expression of nonlinear interdependence between any two areas of cortex was not observed to be different from matched healthy patients. However, the topographic organization of the nonlinear interdependence was found to be different at high levels of significance. Specifically, in the clinical group, the expression of nonlinear interdependence between any two brain regions was less likely to occur as an isolated event, and more likely to be accompanied by the appearance of nonlinear interdependence between multiple other brain regions. Hence, the findings do not support a simple ‘disconnection’ of cortical interactions as implied by the disconnection hypothesis of schizophrenia. Instead, they suggest a loss of the fine-grained organization of cortical interactions and hence can be cautiously interpreted as evidence of an impoverishment of flexibility across hierarchical brain regions in schizophrenia.
Conclusion
Chaos theory in the neurosciences: from ideological to scientific revolution
The qualitative essence of chaotic dynamics – complex behaviour arising from a relatively simple deterministic system – has broad aesthetic and intuitive appeal. The application of nonlinear methods to neuroscientific and psychiatric data can be seen to have occurred in two stages. In the first stage, the concept of the brain as a nonlinear system and the development of practical nonlinear algorithms led to a series of studies, which reported evidence of brain chaos. Concordantly, abnormal patterns of behavioural and EEG chaos were found in the data of patients with schizophrenia. In other words, an interesting and plausible hypothesis was verified in experimental studies. However, this stage was concluded by the recognition of fundamental flaws in the basic nonlinear algorithms.
The most central of these flaws is the lack of explicit hypotheses for statistical testing, allowing false detection of low-dimensional chaos in purely stochastic data. The development of surrogate data techniques represents a major advance in this regard and permits a re-appraisal of previous studies. It appears that there is strong statistical evidence for weak and fluctuating nonlinearity, but not low-dimensional chaos, in brain time series data. The symptoms of schizophrenia and bipolar disorder also appear to be nonlinear in some circumstances. The presence of abnormal patterns of nonlinearity in schizophrenia EEG has not been thoroughly tested using surrogate data techniques and is thus not yet properly established.
It is interesting to compare these two stages from a historical perspective. The first stage is characterized by novel hypotheses that countered the prevailing use of linear methods in neuroscience. Most of the research
In addition to advances in nonlinear theory, the project to understand changes in brain dynamics in schizophrenia needs to keep abreast of advances in schizophrenia. For example, it may be prudent to examine patterns of nonlinear brain dysfunction in clinical ‘subgroups’ of schizophrenia defined using multivariate analysis of symptom profiles [133]. This technique has been used to enhance the detection of abnormal patterns of blood flow in schizophrenia [134] and the detection of abnormal brain activity in schizophrenia as detected by linear techniques [135, 136].
A nonlinear theory of schizophrenia
Given the current state of the theory and evidence reviewed in this article, it is clearly premature to propose a nonlinear theory of schizophrenia. Nonetheless, it may be possible to indicate the essential requirements and form of such a theory. These include: (i) a physiologically derived mathematical model of brain activity; (ii) appropriate dynamical and statistical techniques for measuring neural activity and making inferences on such measures; (iii) an empirically validated method of relating such measures of neural dynamics to cognitive activity; (iv) a sufficiently sophisticated understanding of the phenomenology of schizophrenia; and (v) a suitable experimental design which permits interrogation of the appropriate cognitive activity and neural substrates.
The disconnection hypothesis of schizophrenia lends itself to such a framework, where the term ‘disconnection’ denotes both spatial disorganization of the dynamic behaviours of cortical regions (cognitive incongruency) and temporal disorganization of sequentially expressed dynamical states (formal thought disorder). Heteromodal brain regions, such as prefrontal cortex, appear preferentially affected (in neuroimaging studies) not necessarily because they carry a greater incipient pathology, but rather because in a hierarchical system, distributed disturbances are amplified towards the apex (heteromodal cortex) in comparison to the base (unimodal sensory cortex).
To understand the disturbances in belief and perception that occur in schizophrenia, it may also be necessary to understand how the brain makes causal inferences about the world based upon its sensory inputs. Although this sounds rather ambitious, preliminary work has already suggested that nonlinear interactions across a hierarchically organized system play a crucial role in this regards [137, 138]. With appropriate experimental designs, such a framework shows great potential in meeting the aforementioned criteria for a nonlinear theory of schizophrenia.
Footnotes
Acknowledgements
The author thank E. Gordon, L. Williams and P. Robinson for the excellent supervision during this research. Thanks also to D. Burke for comments on the paper and J. Roberts for assistance with preparation of the manuscript. The author was supported by a Sydney University SESQUI post-doctoral fellow (2002), an Institute of Psychiatry Research Fellowship (2002), Pfizer Neuroscience Research Grant (2003) and Young Investigator Award from the American Association of Psychiatry (2004) during research in this field.
1All patients with schizophrenia in this review were reported as satisfying the DSM-III-R or DSM-IV criteria, unless otherwise stated.
2Twenty-two patients with Research Diagnostic Criteria for schizophrenia compared to 16 healthy patients.
4Cross-checked with Medline and ISI World of Science electronic databases.