
Abstract
This study investigated vertical jumps from single support for two trans-tibial amputees from a standing position. The mechanisms used to achieve flight and the compensatory mechanisms used in the production of force in the absence of plantarflexors are detailed. Two participants completed countermovement maximum vertical jumps from the prosthetic and the sound limbs. The jumps were recorded by a 7-camera 512 VICON motion analysis system integrated with a Kistler forceplate. Flight height was 5 cm jumping from the prosthetic side and 18–19 cm from the sound side. The countermovement was shallower and its duration was less on the prosthetic side compared to the sound side. The reduced and passive range of motion at the prosthesis resulted in an asymmetrical countermovement for both participants with the knee and ankle joints most affected. The duration of the push-off phase was not consistently affected. At take-off the joints on the sound side reached close to full extension while on the prosthetic side they remained more flexed. Joint extension velocity in the push-off phase was similar for both participants on the sound side, though the timing for participant 2 illustrated earlier peaks. The pattern of joint extension velocity was not a smooth proximal to distal sequence on the prosthetic side. The magnitude and timing of the inter-segment extensor moments were asymmetrical for both subjects. The power pattern was asymmetrical in both the countermovement and push-off phases; the lack of power generation at the ankle affected that produced at the remaining joints.
Introduction
The enhanced profile of the Para-Olympics and the consequent development of elite disabled sport have not been fully addressed in sport sciences and specifically by the field of biomechanics. For non-elite athletes there is no evidence of research for recreational sporting activities. Vertical jumping is an activity that is fundamental to many sports, but notably volleyball and basketball. It also offers some insight into the fundamental mechanisms for movement as a result of its clear process and objective, namely to achieve maximum height. An understanding of the biomechanics of amputee jumping and the identification of the compensatory mechanisms employed by amputees to achieve controlled flight is justified. Further, research into the design of prostheses is appropriate, in order to enhance their use in activities which are more vigorous and involve larger forces than walking and running, which have been well researched (Winter and Sienko 1988; Colborne et al. 1992).
The purpose of this research was to redress this deficiency in the biomechanical analysis of amputees. For this, a one-footed countermovement jump from a standing position was chosen to isolate the prosthetic side and to annul any compensatory mechanisms, which may have been recruited by the sound-side, if a two-footed jump had been chosen. While it is acknowledged that this is a somewhat unnatural movement, in that an amputee is unlikely to perform a one-footed jump from a standing position from the prosthesis, it makes it possible to analyse in detail the mechanism chosen by the amputees to achieve flight. This leads to an understanding of the production of the movement in the absence of a key joint (the ankle) and the role of the prosthesis in the jumping action. It will also serve as a useful baseline analysis for other jumps when extra momentum is included, through an approach, or when the sound-limb is also involved in a two-footed jump.
The vertical jump can be broken into four main phases: the countermovement; the push-off; the flight and the landing phases. In order to attain maximum flight height, the velocity of the centre of mass at take-off must be maximised, a process which is achieved in the first two phases. This paper therefore focuses on those two phases. Harman et al. (1990) defined a countermovement as ‘ … a quick bend of the knees during which the body's centre of mass drops somewhat before being propelled upwards'. A countermovement is an action that appears in many activities which require a high end point velocity (e.g. throwing, batting etc.). It is clear that the height gained from the countermovement jump is greater than that of a static jump from the same starting point joint angles (Challis 1998; Bobbert et al. 1996). In the case of amputee jumping, it is possible that the restricted range of passive motion at the prosthetic ankle could have an effect on the depth to which the amputee can lower his body in the countermovement. Consequently the energy stored in the remaining muscles during this stretch-shortening cycle will be reduced. In the prosthesis itself, it is probable that the prosthesis will not store sufficient energy during this phase. Non-amputees have recorded power returns at the ankle of over 1794W in push-off for one-legged jumps (Van Soest et al. 1985). This may result in compensations at the remaining musculature and subsequent asymmetrical loading of the joints.
During the push-off phase of the jump, the ability of the limbs to produce the optimal impulse (force × time) is vital in maximising the velocity at take-off and therefore the height of the jump. This is dependent on an appropriate countermovement, followed by the production of sufficient force through the concentric contraction of the musculature over a useful period of time. Coordination, determined by timing, sequencing and amplitude of muscle activation is an important aspect of this push-off. Kinematic studies have shown that typically in sporting applications, a sequential pattern of movement from proximal to distal joint peak angular velocities is seen in movements where velocity is key to the skill or where there is the involvement of a heavy object. Both of these conditions are present in jumping. Bobbert and van Ingen Schenau (1988) identified the sequence of muscular activation in jumping to be from the upper body, upper legs, lower legs to the feet. The mechanical energy released by the muscles is transported via the biarticular rectus femoris and the gastrocnemius. In the case of amputee jumping, the loss of the ankle joint and its replacement by a prosthesis could have an effect on the coordination of the movement as the most distal joint in the sequence is effectively removed. The push-off could also be affected by the loss of the biarticularity or inadequate tethering of the gastrocnemius.
In order to develop the force to produce the vertical movement a generation of power is required at the joints of the lower limbs. This power also illustrates a proximal to distal sequence (Bobbert and van Ingen Schenau 1988; Aragon-Vargas and Gross 1997) reaching average values of 650W at the hip, 800W at the knee and 1700W at the ankle (Van Soest et al. 1985). These values are significantly higher than those reported for walking. The value at the ankle is probably greater than any prosthesis can return from strain energy stored in the countermovement phase. Compensations will therefore be required to produce the jump. It can be expected that these compensations would be in the remaining limbs and they would not be sufficient to allow the amputee to achieve a similar flight height as non-pathological subjects.
The aim of the study was to assess the biomechanical techniques trans-tibial amputees use to reach maximum flight height in a 1-footed counter movement jump from the sound and also from the prosthetic sides. Through this assessment, alterations to jump biomechanics could be identified and compensatory mechanisms detailed. To achieve this aim a number of variables were analysed: the range of joint movements in the countermovement phase, to examine if the prosthesis and socket had an effect on the depth of the countermovement; the magnitude and timing of joint angular velocities in the push-off phase, to assess the effect the prosthesis had on the coordination of the joints; the kinetic variables of intersegment moments and powers in both phases, to gain greater understanding of the biomechanics of movement production.
Methods
Subjects
Three male trans-tibial amputees volunteered to perform maximal one-legged counter-movement vertical jumps from their right and left limbs. Of the three, only two were comfortable completing the task and achieving height from the prosthetic side. Therefore, one participant withdrew from the research. Both remaining participants were left-sided amputees (LTT). Both participants were active and healthy and were not experiencing any musculoskeletal injuries. Participant one had a mass of 69kg and height of 1.78m. The amputation had taken place 8 years prior to testing as a result of an accident. Participant two had a mass of 81kg and height of 1.77m. The amputation had taken place 12 years prior to testing also as a result of an accident. Both participants wore an Iceross PTB socket with a Mercury R pylon. Participant 1 wore a Seattle-lite foot while participant 2 wore a Mercury Multi-axis foot. The stump condition for both participants was good. The research was submitted to and approved by the Roehampton University Ethics Committee. Both participants completed an informed consent form. Their prosthetist was present during data collection in case of any injury to the participant or damage to the prosthesis.
Data collection
For the motion capture, a 7 camera VICON 512 retro-reflective motion analysis system was synchronised with a Kistler forceplate. The cameras acquired data at a frequency of 120Hz and the forceplate at 600Hz. Thirty-three skin-surface retro-reflective markers were placed with tape on the head, trunk, arms and legs. The markers of the lower-limbs were placed according to the protocol of Davis et al. (1991). For the amputee the markers on the prosthetic side were placed correspondingly to that of the intact limb. For the upper body the markers were placed according to the VICON protocol and were as follows: 4 markers were attached to a headband, placed around the hat line with 2 above the temples and 2 diametrically opposite the temple markers; 6 markers defined the thorax segments, one placed on C7, 1 on T10, 1 on the xiphoid process, 1 on the clavicle notch and one on each acronium process. The upper arms, forearms and hands were defined bilaterally by placing markers on the acromium process, the olecranon process, each styloid process of the radius and ulna and over each knuckle of the middle finger.
Data processing
Kinematic data were filtered using Woltring's cross-validatory quintic spline routine. VICON Plug-in Gait software was used for calculating the three-dimensional angular displacements, moments and powers. The contact time not normalised as the timing of the movement was seen as an important variable for analysis. Kinetic data was normalised to body mass. A single-subject research design for each amputee was selected as it was necessary to gain and apply an understanding of the individual response of each participant and to assess if a common kinematic and kinetic biomechanical pattern is evident. In this type of research it is important to establish whether the participants follow the same mechanics for producing the movement before referring the information to a general population of amputees. The limitation of this design approach in terms of applicability of results to the population is recognised, however, this limitation does not alter the scope and aims of the present study.
The countermovement vertical jumps analysed were (i) one-footed sound limb only; (ii) one-footed prosthetic limb only. The instruction to the participants was simply to jump as high as possible from a standing position. No restraints were placed on the amputees as the research aimed to analyse, initially, how amputees jump.
Both participants warmed up for about ten minutes prior to data collection and they were allowed sufficient time to practice the jumps. At least 3 practice attempts were taken. Each participant then performed 3 jumps of each type and the best jump, defined by maximum height of the Centre of Mass (CoM) (calculated from the kinematic model) was selected for further analysis.
Definition of terms
The start of the countermovement was defined by the beginning of the descent of the CoM; take-off was determined by unloading of the forceplate; flight height was defined as maximum height less take-off height of the CoM; depth of the countermovement was taken as standing height less maximum downward displacement of CoM. Flight height and depth of countermovement were defined in this way to allow comparisons between the jumpers, to negate the effect due to the height of the participants and arm movement. These measurements were calculated from the kinematic data of the CoM.
The flight height, depth of the countermovement, duration of the countermovement and push-off phases, maximum joint flexion angles in the countermovement phase and joint extension angle at take-off, joint angular velocity magnitude and sequence of reaching maximum value and moments and powers in the countermovement and push-off phases for the different jumps were analysed.
Results
The results for both participants illustrate the process and outcome of the one-footed countermovement jump.
The primary outcome measure for the vertical jump is flight height. The participants were evenly matched in terms of flight height in both conditions, though the jump from the prosthetic limb was substantially lower than that from the sound limb for both participants (Table I). On both the sound and the prosthetic side the height was lower than for trained non-amputee athletes who achieved 31 cm (Van Soest et al. 1985). Both amputees only achieved 5cm of flight when jumping from the prosthetic limb. The countermovement was much shallower from the single-prosthetic limb for both participants (Table I). This may be due to poor balance as well as from joint immobility. The duration of the countermovement phase was similar for the sound and prosthetic sides for participant 1, even though the depth was much shallower. For participant 2, the duration of the countermovement on the prosthetic side was much shorter and the duration of the prosthetic-side countermovement for participant 2 was substantially less than for participant 1 even though the depth was similar. The duration of the push-off phase, which has implications for the impulse, was not consistently affected, participant 1 increased the time of push-off compared to the sound side, even though the depth from which to push was not as great. Participant 2 reduced the time of push-off compared to the sound side. It had been anticipated that the time of the push-off would have been increased to compensate for the reduced force-production capacity of the prosthetic limb, but given the limited range of motion the extent of the increased duration for participant 1 was unexpected.
The knee and the ankle joints were the joints most affected in the countermovement phases with substantially less flexion at these joints on the prosthetic side compared to the sound side (Table II). The prosthetic side knee was 21° more extended for participant 1 and 30° more extended for participant 2 compared to the sound side. The lack of mobility at the prosthetic ankle was obvious with 16° less maximum dorsiflexion for participant 1 and 40° less for participant 2 compared to the sound side results. The prosthesis dorsiflexed by only 2° less for participant 2 compared to participant 1. The hip was more symmetrical for participant 1 with 8° less flexion on the prosthetic side. For participant 2 the hip was 168 less flexed.
Outcome results for vertical countermovement jump.
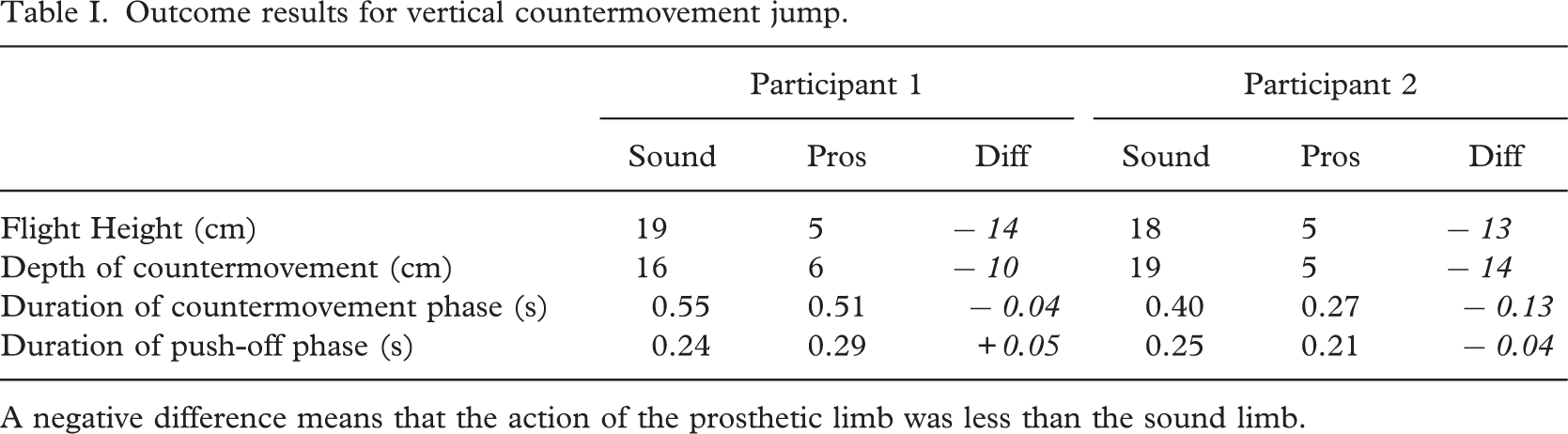
A negative difference means that the action of the prosthetic limb was less than the sound limb.
Kinematic results for the countermovement (cm) and take-off phases.
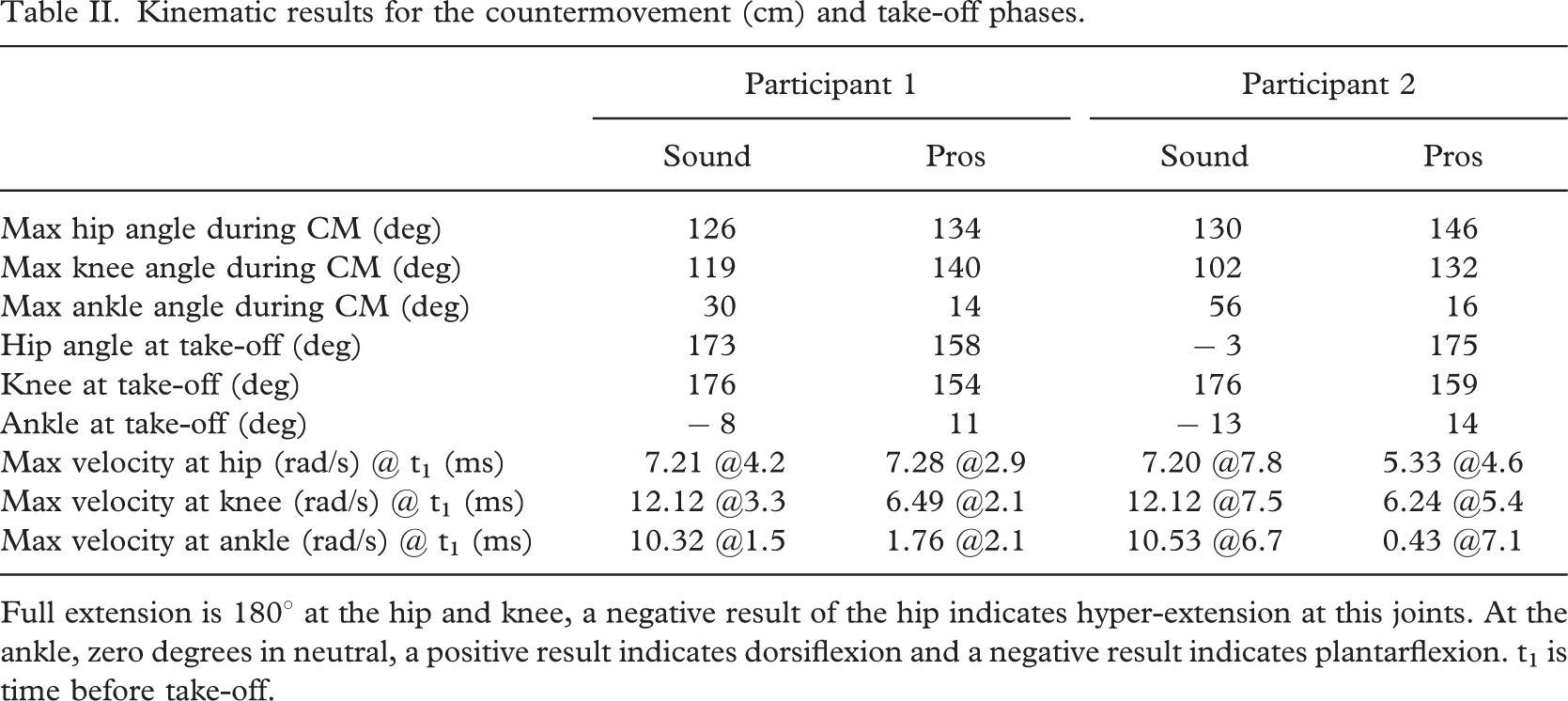
Full extension is 1808 at the hip and knee, a negative result of the hip indicates hyper-extension at this joints. At the ankle, zero degrees in neutral, a positive result indicates dorsiflexion and a negative result indicates plantarflexion. t1 is time before take-off.
During the countermovement from the sound limb, participant 2 flexed his knee and dorsiflexed his ankle more than participant 1 but flexed his hip less. This resulted in marginally different kinematics in the production of the countermovement. On the prosthetic side, participant 2 flexed his hip more and flexed his knee and ankle slightly more.
For both amputees at take-off, it is clear that the joints on the sound side went through a full range of motion reaching close to full extension while on the prosthetic side the extension was not as great at the ankle and knee and hip, though participant 2 reached near to full extension at the hip.
When considering joint velocity data in the push-off phase, both the pattern and magnitude are important. The pattern of joint extension velocity was very similar on the sound side for both participants. A proximal to distal sequence of extension maximum velocity for both participants prior to take-off was evident. This pattern is also found in the literature (Van Soest et al. 1985). A feature of the results presented here is that participant 2 values reached a peak earlier than participant 1. The magnitude compared well with other results for one-footed jumps (Van Soest et al. (1985) report values of: hip 8 rads-71, knee, 12 rads-71 and ankle, 16 rads-71).
By contrast, on the prosthetic side for participant 1 the joint angular velocity data was of smaller magnitude for the knee and ankle though similar for the hip. The reduced angular velocity on the prosthetic side illustrates the affected kinetics in the action-phase, discussed below. The pattern of peak velocity was similar to that for the sound side except that the knee and ankle reached their peak velocity simultaneously for participant 1. Further, the peaks were reached closer to take-off on the sound side. For participant 2 the pattern of peak joint velocity followed the sequence ankle, knee and finally hip. The magnitudes at all joints were less than that for the sound side. The peak velocity at the ankle was less than for participant 1. These results indicated a poor return from deformation of the prosthetic ankle as well as poor timing in the return–it reached the maximum velocity first rather than last in the sequence.
The inter-segment moments are presented in Figure 1 to illustrate how the amputees produce the movement. The graphs illustrate the data from the initiation of counter-movement to take-off for both jumps.
Clearly for participant 1 (Figure 1) there was a different intersegmental moment pattern for the sound compared to the prosthetic limb. This was obvious for both the countermovement and the push-off phases. In the countermovement phase, the ankle showed an asymmetrical pattern, tending towards a plantarflexor moment for the sound limb but it remained quite passive on the prosthetic side.
In the push-off phase, the magnitude and timing of the extensor moment about the joints was different between the limbs. On the sound side, the sequence for peak joint moment was hip–335Nm, ankle–322Nm then knee–133Nm. For the prosthetic limb, there was a more prolonged moment about the joints probably to compensate for the ankle joint. The sequence was hip–192Nm, ankle–178Nm and knee–124Nm. The knee moment was similar for both jumps (sound, prosthetic).
For participant 2 (Figure 1) there was also a different intersegmental moment pattern for the sound compared to the prosthetic limb. However, apart from the duration, this difference was not as noticeable as for participant 1.
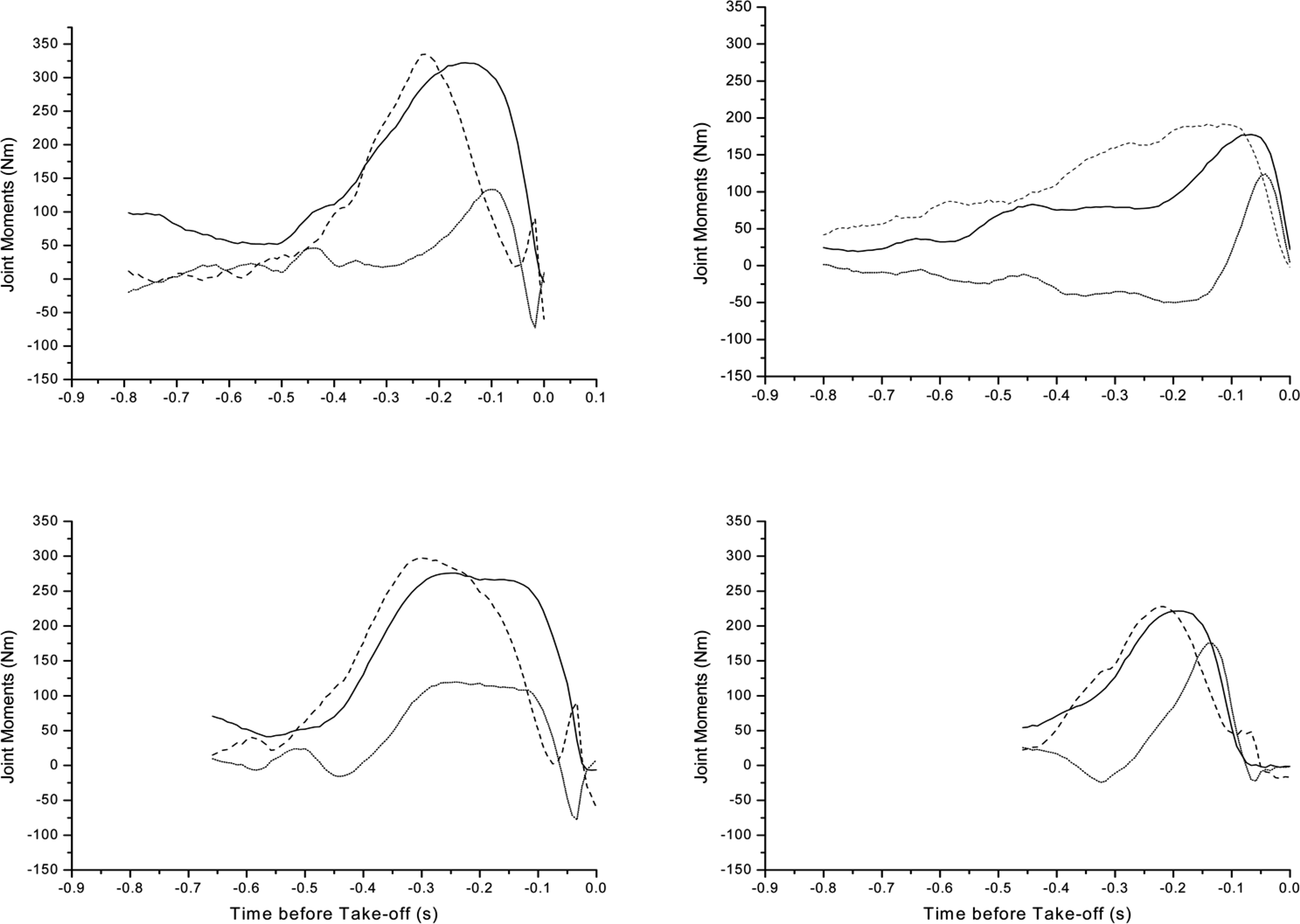
Joint Moment patterns for participant 1 (top) and participant 2 (bottom). Sound side movements are indicated on the left and prosthetic side on the right. Take-off is at time-zero. The different data length indicates the time difference between the participants and the jumps from initiation of the countermovement to take-off. The jump from the prosthetic side took the shortest time for participant 2 who also took the longest time from the sound side. Data are presented for the ankle (____), knee (……) and hip (____).
In the push-off phase, the magnitude of the extensor moment about the joint axes was different between the limbs. On the sound side, the intersegmental moments reached the maximum in a sequence of hip, ankle and knee. The pattern was not repeated on the prosthetic side, where more peaked action was obvious. The sequence was hip, ankle then knee. The hip intersegmental moment was lower on the prosthetic side compared to the sound side (sound 297Nm, prosthetic 228Nm). The knee (sound 120Nm, prosthetic 175Nm) increased the contribution on the prosthetic side. The reduced ankle intersegmental moment (sound 276Nm, prosthetic 221Nm) illustrated that the prosthesis is contributing asymmetrically but the magnitude illustrates that the prosthesis is contributing more than for participant 1.
The joint moments for one-legged standing countermovement jumps in the literature were similar for the sound limb. Van Soest et al. (1985) reported hip joint moments for a similar jump type, though for trained athletes as: hip 280Nm, knee 180Nm and ankle 230Nm. For both the amputee participants, the values for the hip and ankle on the sound side were marginally higher, and that for the knee was marginally lower.
Figure 2 illustrates the power patterns at the ankle, knee and hip, demonstrating how the participants produced the movement in the countermovement and push-off phases. Power patterns at the joints are analysed as they lead to an understanding of the role of the muscle groups about the joint as they shorten and lengthen under tension. Positive power indicates a concentric contraction of the muscles about the joint and negative power an eccentric contraction.
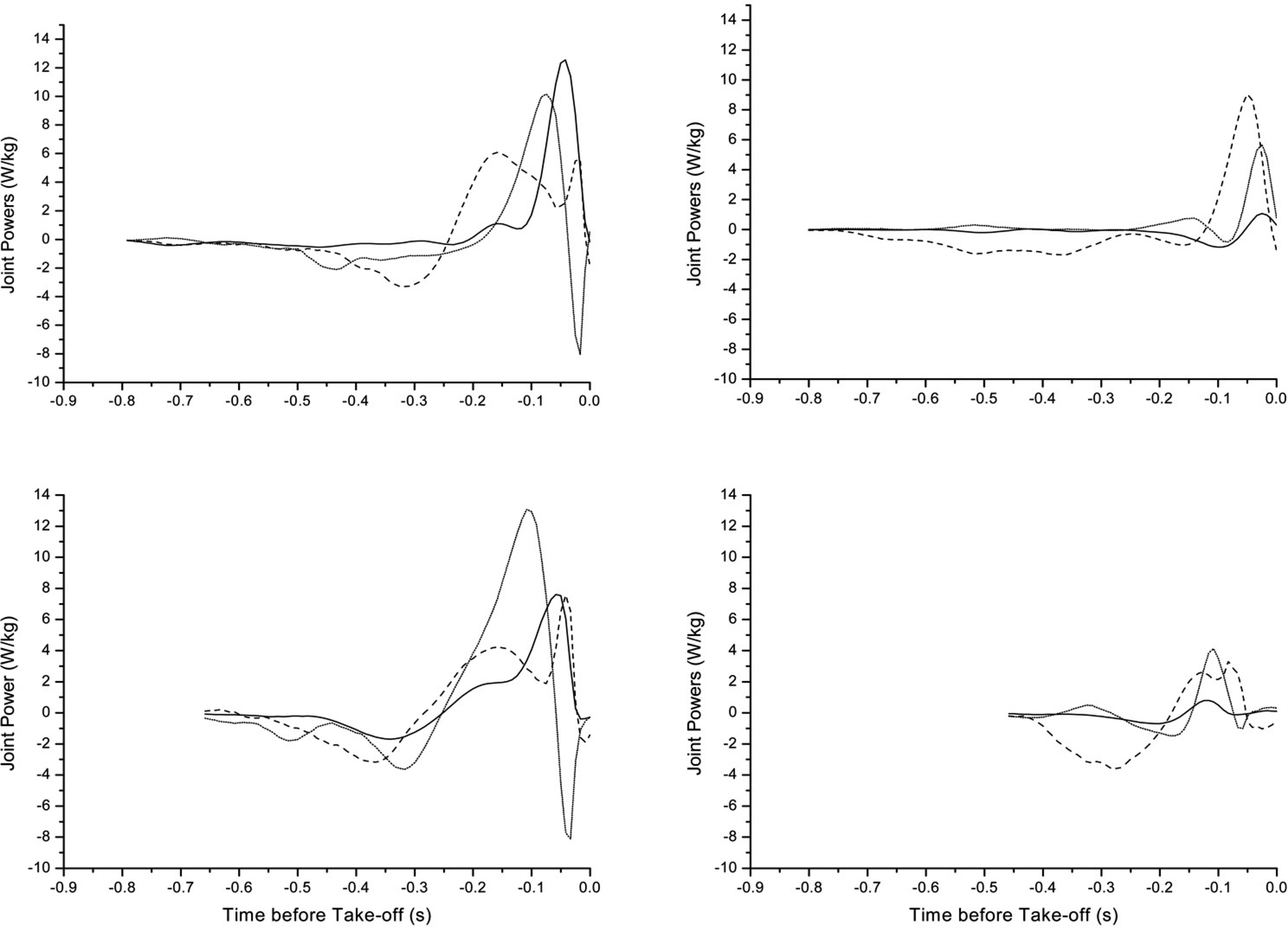
Joint Power patterns for participant 1 (top) and participant 2 (bottom). Sound side movements are indicated on the left and prosthetic side on the right. Take-off is at time zero. Data are presented for the ankle (___), knee (….) and hip (——). The different data length indicates the time difference between the participants and the jumps from initiation of the countermovement to take-off.
The close to zero power pattern in the initial phase of the countermovement reflected the lack of muscular contraction as the body descended under the action of gravity. This was followed by an absorption of power during the eccentric contraction, close to the bottom of countermovement. This eccentric contraction halts the descent and is immediately followed by a concentric contraction about all the joints in action phase.
For participant 1 (Figure 2), at the later part of the countermovement phase on the sound side, there was an absorption of power at the knee and the hip which controlled the lowering of the body under the eccentric contraction. During power production this changed to positive power in order to propel the body into flight. The hip peak power (at sound side 6W/kg compared to prosthetic side 9W/kg) was greater on the prosthetic side. The knee, as a result of its reduced range of motion produced less power on the prosthetic side at 6W/kg compared to 10W/kg on the sound side. There was some return of power from the prosthesis in the push-off phase (1W/kg). This was substantially smaller compared to the sound side at 13W/kg, as a result of the passive return from the prosthesis.
On the sound side, in the countermovement phase, for participant 2 more power was absorbed than for participant 1. The hip, knee and ankle were involved in this phase. In the push-off phase, this participant illustrated a proximal to distal power generation. The knee was the main source of power for this participant, generating 13W/kg of peak power. The second peak at the hip at 8W/kg is greater than the first at 4W/kg and was later than that of the ankle (8W/kg). Similar to participant 1, it coincided with the absorption of power at the knee prior to take-off.
For participant 2 (Figure 2) on the prosthetic side, in the countermovement phase the hip was the main power absorber. Absorption begins immediately and indicated that the amputee did not initially descend with gravity. The power pattern at the knee indicated the balance difficulties experienced by the participant in this phase. Unlike participant 1, less power was generated at all the prosthetic side joints in this phase. At the hip there were two power peaks about the joint reaching lower peaks than for the sound side both at 3W/kg. The knee power was obviously asymmetrical, the prosthetic side being one third of that at the sound side at 4W/kg. Again, at the ankle the asymmetry was most obvious with the sound side being 8 times that of the prosthetic side, an indication of the limitations of the prosthetic ankle. The prosthetic side peaked at only 1W/kg. The pattern is closer to simultaneous than proximal to distal.
Discussion
A two case-subject research design was adopted to describe the kinematic and kinetic biomechanical pattern for the two participants and the results were used to compare the two participants. This determined if similar biomechanics were used to produce this controlled jump. In research of this type, it is important to establish whether the participants are following the same method for producing the movement before the data can be grouped. In the experiment reported here, for the two participants, the flight height achieved was remarkably similar. This indicates that, at least for outcome measurements, the amputees were both jumping in a similar manner. When the underlying biomechanics were analysed, the results indicate that on the sound side, the biomechanics were similar to those of non-amputee subjects. On the prosthetic side, the amputees show slightly differing techniques and the biomechanics were somewhat different in comparison to non-amputee subjects. For a larger sample size, grouping of the data should be treated with caution, depending on the variance between the subjects.
So-called ‘dynamic” prostheses (e.g. the flex-foot series) have been developed to enhance the active lifestyle of lower-limb amputees. At the elite level of sport the prostheses appear to be extremely effective, with world record achievements for amputees appearing close to those of non-amputees. These prostheses allow participation in events to a similar standard as non-amputees, often alongside non-disabled athletes. However, these prostheses are often not prescribed for non-elite athletes due to their cost. There are some prostheses which do not have the same high specifications but which still have some dynamic properties. These prostheses are regularly prescribed to active amputees and facilitate low-impact activity participation such as power walking, jogging and dancing.
All “dynamic” prostheses function on the same basic principle. Under loading, energy is stored as elastic energy in the structure. This energy is then returned when the prosthesis is unloaded. The mechanics of the spring are often altered depending on the weight of the user and/or their activity level. If the spring is correctly adjusted and optimally tense then the energy returned should facilitate participation in the activity and complement the remaining musculature. As little research has been carried out for amputees in jumping, it is difficult to determine the spring set-up and adjust it accordingly. However, before any further development can take place in the design of the prostheses, it is necessary to understand the biomechanical limitations experienced by amputees which could affect their ability to jump.
While it was expected that the loss of the lower limb would have an effect on the jump, the extent of the effect was greater than anticipated. The participants were barely able to achieve flight in jumping, indicating that the prosthetic side was extremely inhibited by the amputation. This implies that the jumping mechanics were substantially affected by the lack of the ankle joint and its associated musculature. This resulted in an altered process by which flight was achieved.
In any jump, the countermovement phase is an important part of the jump for two reasons. Firstly, it increases the range of motion over which the joints can later contract concentrically in the push-off phase, and secondly, it enables the storage of elastic energy in the muscles in the eccentric phase. For both amputees the depth of the countermovement was greatly reduced as a result of the reduced range of motion at the ankle. Van Soest et al. (1985) found that in the countermovement phase the maximum joint flexion at the hip was 968, at the knee was 1048 and that the ankle dorsiflexed to 658. The results are different to those in the literature for both limbs. On the sound side, the hip is more extended for both participants, the knee is more extended for participant 1 but similar for participant 2 and the ankle is less dorsiflexed for participant 1 and similar for participant 2. This is possibly as a result of the untrained status of the amputees and their consequent lack of flexibility and strength. On the prosthetic side, the hip and knee are more extended and the ankle less dorsiflexed for both participants. It could be that this reduced range of motion was a safety issue or that the proprioceptive feedback from the prosthesis was not sufficient to reach a full range of motion. This kinematic pattern resulted in less countermovement which lasted for a shorter duration. The reduced moment at the prosthetic ankle was expected due to the passive nature of the joint reacting under loading. The absorption of power in the phase is both individual and asymmetrical. Participant 1 absorbs power over a longer duration throughout the countermovement and participant 2 begins to absorb power from the initiation of the movement, over a shorter time period than participant 1. For the remaining joints on the prosthetic side the less stable moment patterns and the more poorly timed power absorption sequence, illustrated the problem that the participant had in maintaining his balance. The maintenance of balance, a factor on which both amputees commented, is a complication facing amputees jumping from one leg. Prior to beginning the counter-movement phase, jumping from one limb requires that the athlete be balanced and comfortable on that limb. It has previously been shown that bipedal standing is altered in lower limb amputees compared to control subjects due to the presence of a prosthesis (Isakov et al. 1992). The participants were concentrating more on maintaining their balance than on descending with gravity in the countermovement. Previous literature which compares one-legged versus two-legged jumps (Vint and Hinrichs 1996; Challis 1998) has not looked at the issue of balance and its effect on the jump, probably because for non-injured athletes it is not deemed to be a problem. Further research in both amputees and controls is warranted.
The duration of the push-off phase was not consistently affected with participant 1 following the expected result of increasing the duration of this phase, while participant 2 reduced the phase duration. The extent of the increase from participant 1 was greater than anticipated considering the small range over which the movement was conducted. It had been anticipated that the length of time of push-off would have been increased to compensate for the reduced force-production capacity of the prosthetic limb. The reduced force-production is evident through the less well coordinated pattern of joint extension velocity, the lower joint moments and the lower joint power results for the prosthetic side compared to the sound side for both amputees.
In the push-off phase on the prosthetic side, the main producer of power for participant 1 was the hip, which reached a single peak, followed by the knee and then the ankle. For participant 2, the knee was the main producer of power, closely followed by the hip, which retained the double hump seen on the sound side.
On the sound side, participant 2 used his knee as the main power source. He also illustrated an extra burst of absorption at the knee in the transition from the countermovement to the push-off phase. Participant 1 used the ankle as the main power source.
Van Soest et al. (1985) present data which has not been normalised by body mass for non-amputee subjects. Joint powers are reported as hip 650W, knee 800W and ankle 1700W. Converting the results presented here from normalised results to non-normalised results, participant 1 produced a sound side hip power of 414W and prosthetic side hip power of 621W, sound side knee power of 690W and prosthetic side knee power of 414W and sound side ankle power of 870W and 69W on the prosthetic side. These results were lower for the amputee than for the non-amputee subjects, and indicate why the amputee had a reduced flight height compared to the 31cm presented by Van Soest et al., even from the sound side.
Participant 2's non-normalised power results were also asymmetrical and different from the literature with the sound side hip power of 648W, knee power of 1053W and ankle power of 648W while on the prosthetic side the hip peak power was 243W, the knee 324W and the ankle 81W.
The reduced power at the joints is a consequence of the reduced range of motion over which the muscles could produce force. The mechanical properties of skeletal muscle, including muscle length, affect the performance of any movement. Specifically, isometric studies on the force-length relationship of skeletal muscle have suggested that maximal muscular force depends on its length, which is determined by the angle of the joint(s) that the muscle is crossing. For the prosthetic side one-footed jump, the reduced range of motion resulted in the muscular force about all the joints at the lower limb being reduced. The result was that the amputee did not produce force at an optimal muscle length. Also as a result of the reduced countermovement, less energy could be stored as elastic energy in the muscles about the knee and hip joints. At the prosthetic ankle, insufficient energy was stored in the countermovement phase to be effective in compensating for the loss of the ankle plantarflexors. This was indicated by the reduced power profile in both the countermovement and the push-off phases for that joint.
A further reason for the reduced height of the jump was the coordination of the movement. Given the biomechanically complex nature of jumping appropriate neuromuscular control is vital in achieving the task. The proximal to distal muscle recruitment was interrupted. For non-pathological trained athletes, a link-chain proximal-to-distal sequence of muscle activation (i.e. muscles spanning the hip are activated first, then the knee and finally those at the ankle) has been shown (Bobbert and van Ingen Shenau 1988; Pandy and Zajac 1991). This sequence requires that joint moments are produced at high velocities, especially at the ankle (Van Soest et al. 1995). If this sequence is disrupted, it is likely that jump height will be affected. While the participants in this study illustrated a pattern of recruitment on the prosthetic side, the lack of power generation at the ankle had an obvious effect on the remaining joints. The result was that the order of peak powers was not as fluent as that for the sound limb, or that which is seen in the literature. This different timing of the joint maxima indicates a different motor pattern.
On the sound side, there was a clear proximal to distal pattern of power generation at the joints. The hip illustrated a double peak, the second peak coincided with a negative power burst at the knee immediately prior to toe-off. This negative peak slowed the extending knee down and prevented extension at this point. Comparing the two participants, the method of absorbing power in the countermovement phase and generating the power in the push-off phase was somewhat different on both the sound and prosthetic sides. This difference was more marked on the prosthetic side. In the countermovement phase on the prosthetic side, participant 1 illustrated an extended phase over which the hip continued to absorb power, it was the only absorber of power in this phase, the ankle and knee were neutral, probably to ensure balance. Participant 2 illustrated a very short phase with a larger absorption at the hip which was evident from the initiation of the movement.
Conclusion
A maximal-effort vertical jump from standing was the control and asking the participants to alter the limb used, while maintaining the maximal propulsive effort, modified the task. The research aimed to assess the variation of the movement for the amputees using their sound and prosthetic limbs. Flight height was substantially reduced when jumping from the prosthetic limb, implying that the loss of a single joint, the ankle, can have a profound effect on the ability of the remaining musculature to produce the movement. The effect was clear in the countermovement and the push-off phases. The depth of the countermovement was substantially reduced for both participants when jumping from the prosthetic limb. This resulted in a reduced effectiveness of this phase due to (i) the reduced range of motion over which the force could be produced; (ii) the probable reduction of energy stored in the prosthesis compared to the sound ankle and (iii) the probable reduction of strain energy in the remaining muscles due to the reduced stretch-shortening cycle. The reduced range of motion was most obvious at the knee and ankle joints.
As a result of the shallower countermovement-phase, the push-off phase was also affected. The usual proximal to distal joint moments, powers and angular velocities were disrupted and the magnitudes of the variables reduced. This reduction resulted in a lower overall push-off impulse and as a consequence the jumpers did not achieve the same flight.
There are four possible explanations for the reduced jump height: firstly, the prosthesis was unable to compensate sufficiently for the lost limb; secondly, as a consequence of the amputation the strength and muscle mechanics of the amputees affected the biomechanics of the jump; thirdly, the amputation affected the proprioception of the amputees who could not then coordinate the jump effectively and finally, the impaired balance resulting from the amputation resulted in an inability to prepare adequately and recruit the muscles effectively for the jump. Further work is required on each of these theories to determine to what extent each has an influence on jump height.
Acknowledgement
The researchers would like to thank John Ross at Chas A Blatchford and Son for his advice throughout the project and his assistance in completing the testing procedure.