
Abstract
End plates serve as the interface between rigid vertebral bodies and pliant intervertebral disks. Because the lumbar spine carries significant forces and disks don't have a dedicated blood supply, end plates must balance conflicting requirements of being strong to prevent vertebral fracture and porous to facilitate transport between disk cells and vertebral capillaries. Consequently, end plates are particularly susceptible to damage, which can increase communication between proinflammatory disk constituents and vascularized vertebral bone marrow. Damaged end plate regions can be sites of reactive bone marrow lesions that include proliferating nerves, which are susceptible to chemical sensitization and mechanical stimulation. Although several lines of evidence indicate that innervated end plate damage can be a source of chronic low back pain, its role in patients is likely underappreciated because innervated damage is poorly visualized with diagnostic imaging. This literature review summarizes end plate biophysical function and aspects of pathologic degeneration that can lead to vertebrogenic pain. Areas of future research are identified in the context of unmet clinical needs for patients with chronic low back pain.
Chronic low back pain remains a difficult clinical problem, both to diagnose and to treat. Despite significant investments in basic and clinical research, the rates of disability and associated costs continue to escalate. 1 Although the prevailing view is that axial back pain arises from sensitized nociceptors within the annulus fibrosus of degenerating disks (annulogenic pain), there is growing evidence that the end plates are richly innervated and that innervated end plate damage may represent a common painful pathology (vertebrogenic pain). 2 , 3 Properly identifying the pain generator is requisite for optimal treatment, so distinguishing between these forms of pain will likely be important for improving patient outcomes. The goal of this review is to summarize data regarding normal end plate anatomy, physiologic age-related end plate changes, and evidence for the role of pathologic changes as a source of chronic low back pain. In an effort to cover these topics within a clinical context, we have focused our summary on the end plates in the human spine. We refer the reader to the literature for a detailed comparison of end plate anatomy and biochemistry between humans and animals. 4 , 5 Related, we acknowledge that it remains open for debate whether the end plate belongs to the vertebral body or to the intervertebral disk. Rather than presenting a specific viewpoint, we consider topics that are relevant to both its bony and cartilaginous components. 6
Structure
The end plate is a bilayer of cartilage and bone that separates the intervertebral disks from the adjacent vertebrae (Fig. 1A to C). During prenatal development, the future vertebra starts as a cartilage anlagen that arises from chondrification centers of the sclerotomes during the sixth embryonic week (Fig. 2). 7 The anlagen begins ossification at its centrum around invading blood vessels. 8 This trabecular centrum is separated from the forming disk by an epiphyseal plate of columnar cartilage that progressively thins as the vertebra lengthens. Peripheral to the epiphyseal plate is a ring apophysis that doesn't participate in longitudinal growth, but is rather a traction apophysis by virtue of annular fiber insertion. 9 Yet, the ends of the vertebrae are completely covered by the same end plate cartilage. By age 18, the epiphyseal cartilage has thinned and a subchondral bone plate has formed, thus creating the adult end plate bilayer. Simultaneously, the ring apophysis fuses to the vertebral body.
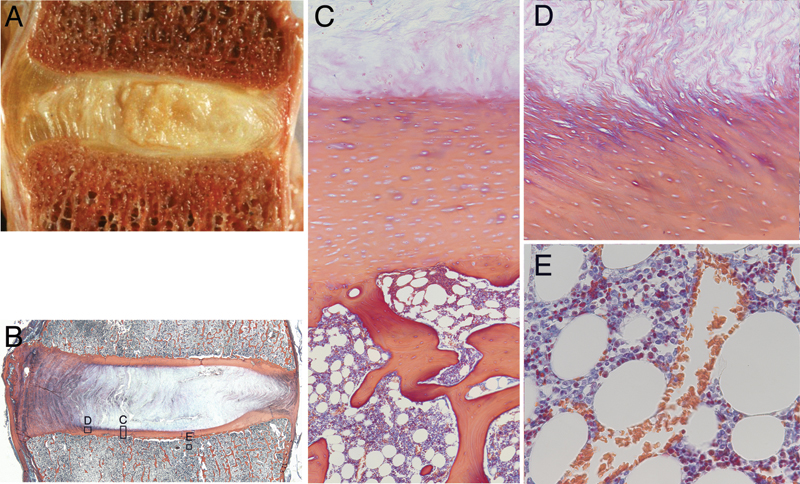
(A) Gross morphology of the lumbar intervertebral joint. (B) Histology section showing regions of interest for panels C, D, and E. (C) End plate detail showing cartilaginous and bony components with hematopoietic marrow elements. (D) Insertion of annular fibers into the end plate cartilage at the inner annulus junction. (E) Vascular sinusoids in the marrow space adjacent to the end plate. Note for panels A and B, left side is anterior.
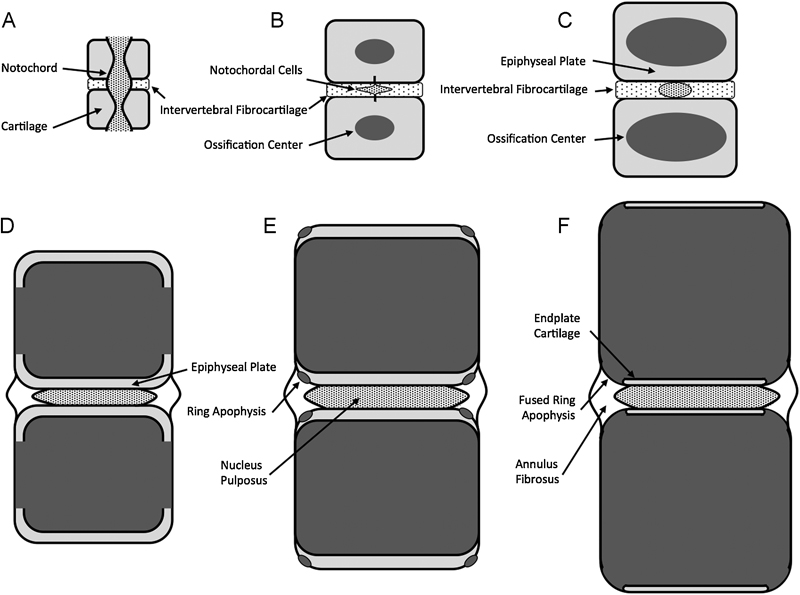
Schematic representation of vertebral end plate development. (A) At embryonic week 6, the sclerotome begins to segment around the notochord to form periodic cartilaginous and fibrocartilaginous precursors to the vertebra and disks, respectively. (B) By embryonic week 15, the notochord atrophies within the vertebra, and ossification begins at the vertebral centers. (C) At embryonic week 25, the ossification centers expand as the vertebrae lengthen. Columnar cartilage develops at the vertebral ends to form the epiphyseal plates. (D) By age 5 years, the ossified portions of the vertebra extend to the lateral margins and the epiphyseal cartilage begins to thin. (E) By age 13 years, peripheral ossification centers outside the epiphyseal plate form the ring apophysis. (F) By age 18 years, the ring apophysis begins to fuse to the osseous mass of the vertebral body.
Like articular cartilage, the end plate cartilage consists of chondrocytes interspersed throughout an extracellular matrix of proteoglycans, collagen (types I and II), and water (Fig. 1C). However, the end plate cartilage differs from articular cartilage in its collagen fiber organization. Although healthy articular cartilage has zones of differing collagen orientation, end plate cartilage has collagen fibers aligned horizontally (parallel to the ends of the vertebrae). 10 In the young disk, the end plate cartilage proteoglycan content is ~300 µg/mg, with water and type I collagen contents being 78% and 0.9 ng/mg, respectively. 11 The cartilage end plate is typically between 0.1 and 2.0 mm thick 12 , 13 ; however, its thickness is known to vary with position and level, being thinner centrally and in the upper levels of the spine than peripherally and in the lower levels of the spine. 13 The nature of the structural integration between the end plate and the surrounding tissues also varies with position. Peripherally, the collagen fibers in the lamellae of the annulus fibrosus are continuous with the collagen fibers in the end plate (Fig. 1D), whereas centrally, the integration between collagen fibers in the nucleus pulposus and the end plate is more convoluted. 13 , 14 The collagen fibers of the cartilaginous and bony components of the end plate are completely separate. 13
The bony component of the end plate has a structure not unlike that of the vertebral cortex and resembles a thickened, porous layer of fused trabecular bone with osteocytes entombed within saucer-shaped lamellar packets. 15 Like the end plate cartilage, the bony end plate thickness varies depending on spinal level and location and is generally between 0.2 and 0.8 mm thick. 16 , 17 , 18 , 19 At a given lumbar level, the bony end plates are thinner centrally than peripherally; also, the end plate cranial to a particular disk is thicker and has higher bone mineral density than does the end plate caudal to it. 18 , 20 In some individuals, a second, dense layer of bone exists below the superficial layer. 16 , 19
The bone marrow compartment adjacent to the bony end plate consists of hematopoietic cells, fat cells, sinusoids (thin-walled capillaries), and nerves. The vertebral capillaries and nerves enter via the basivertebral foramen at the posterior vertebral cortex and small pores in the cortical shell, form an “arterial grid” at the vertebral centrum, then branch and terminate just adjacent to the cartilage end plates. 21 , 22 , 23 These sinusoids and nerves provide a continuous bed across the bone–disk interface (Fig. 1E). 21 Importantly for the disk, there is an intimate relationship between the effective perfusion of these sinusoids and marrow cellularity type. For example, perfusion decreases as thicker-walled capillaries replace the sinusoids, which can occur when the hematopoietic marrow converts to fat. 24 , 25 The cause of this marrow conversion is unclear, but the increase in bone marrow adiposity may be related to declining bone mass, 26 ,27, 28 , 29 , 30 vascularity, 29 , 31 temperature, 25 or reduced bone loading. 32
End plate subchondral bone is innervated by basivertebral nerve, the fibers of which reach the bone marrow along with nutrient arteries that enter the vertebra through the posterior basivertebral foramen. 21 , 33 , 34 , 35 End plate innervation is comparable to that of the peripheral annulus, 2 , 3 , 36 and it is increased in areas of bone damage (Figs. 3 and 4). 37

Distribution of protein gene product 9.5 (PGP 9.5)-positive nerve fibers across the end plates (63-year-old woman, L5–S1). Compared with the density of nerves in normal end plate regions, nerve density is higher in end plate regions with damage. Nerve fibers in this disk were observed in the inferoposterior outer annulus. Note: left side is anterior.
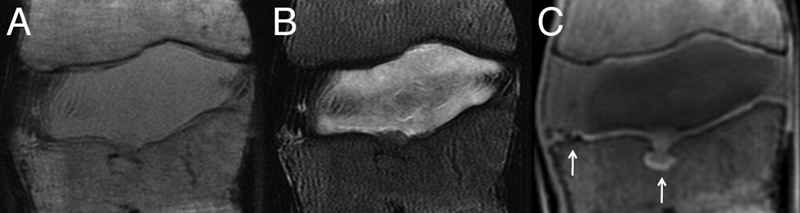
Midsagittal T1-weighted (A) and T2-weighted (B) magnetic resonance (MR) images of an L1–L2 motion segment with poor end plate signal. (C) Corresponding ultrashort time-to-echo (UTE) MR image showing enhanced end plate signal. Arrows indicate end plate defects shown in Fig. 5A and 5B. (UTE imaging courtesy of Drs. Roland Krug and Misung Han, Department of Radiology, University of California, San Francisco.)
Biophysical Function
The structure of the end plate facilitates important biomechanical and nutritional functions. Biomechanically, the end plate is subjected to significant loads during activities of daily living as the trunk muscles contract to stabilize posture. Lumbar compression forces can be in the range of 800 N while standing upright to over 3,000 N during active lifting. 38 The nucleus becomes pressurized in response to these forces, the values of which have been measured to vary from 0.4 MPa while lying, to 1.5 MPa while standing and sitting, to 2.3 MPa while lifting. 39 , 40 , 41 The end plate distributes these intradiscal pressures onto the adjacent vertebrae and prevents the pressurized disk nucleus from bulging into the underlying trabecular bone. 42 , 43 , 44
During spinal compression, the pressurized nucleus causes the end plate to be stretched, like a drumhead. 45 Consequently, the end plate is most prone to fail in tension. Ultimately, thickness, porosity, and curvature are important structural determinants of end plate biomechanical function: thick, dense end plates with a high degree of curvature are stronger than thin, porous, and flat end plates. 18 , 46 ,47, 48 , 49
Nutritionally, the end plate is the primary pathway for transport between vertebral capillaries and cells within the disk nucleus. 50 , 51 Blood vessels and marrow spaces abut the cartilage layer (Fig. 1C, E) and provide channels for glucose and oxygen to enter the disk and for waste products to exit the disk. Permeability across the cartilage end plate correlates with the amount of direct contact between the end plate and vertebral marrow or vascular buds. 50 The typical marrow contact area (or effective exchange area) is between 10 and 40%, 52 , 53 , 54 with the central end plate (adjacent the nucleus) being more permeable than at the periphery. 50 The density of these vascular channels is higher adjacent to the disk nucleus than the annulus. 13 , 50 , 55
Once nutrients reach the end plate, movement of small solutes (glucose, lactate, and oxygen) pass through disk matrix primarily by diffusion. 56 , 57 Larger solutes may also be influenced by convective fluid flow created by mechanical disk compression and recovery. Diffusion into the disk is driven by the concentration gradient between the blood plasma and tissue matrix and represents a balance between supply (capillary density) and demand (disk cell density and metabolic rate).
Consequently, the end plate must balance conflicting biophysical demands. It must be strong to resist mechanical failure but must also be porous to facilitate chemical transport. Thin, porous end plates may favor disk health and thick, impermeable end plates favor vertebral integrity. 18 , 58 Recent data indicate that double-layer end plates may provide a more optimal balance between end plate strength and porosity, 15 thereby protecting against damage while supporting improved transport to and from adjacent disks.
Physiologic Degeneration
During aging, the cartilage end plate experiences changes in proteoglycan and collagen, resulting in gradual thinning and calcification. 11 , 13 , 59 , 60 Proteoglycan content decreases from 300 µg/mg at age 2 to 150 µm/mg by age 80. Simultaneously, water and type I collagen decrease from 78 to 67% and from 0.9 ng/mg to 0.25 ng/mg, respectively. 11 Although the specific mechanisms responsible for the compositional deterioration are unclear, these age-related changes coincide with degeneration in the adjacent disk and are generally consistent with markers of chondrocyte hypertrophy (e.g., elevated expression of type X collagen). 11 , 61 Hence, it may be that factors such as diminished hydrostatic pressure play a role in end plate deterioration because hydrostatic pressure is a potent regulator of chondrocyte function. 62 , 63
When the spine is compressed, the bony end plate is subjected to high tensile strains as it deforms into the underlying trabecular bone. 42 , 43 , 44 , 45 , 64 Several factors influence the susceptibility of the end plate to damage, including the nature of the mechanical loading, the local morphology of the end plate structure, the tissue material properties, and the condition of the intervertebral disk. End plates at the cranial vertebral aspect may be more susceptible to damage than caudal end plates because they are thinner and supported by less dense trabecular bone. 18 Likewise, damage often occurs to the central end plate, the thinnest and weakest region. 18 , 65 Accumulation of end plate damage can cause focal weak points that progress to circumferential fissures. 44 , 66 , 67 This potential is exacerbated with age as the central region of the bony end plate becomes more porous (~60%) and consequently less stiff and weaker as the adjacent disks degenerate. 12 , 19 , 68 , 69 These deleterious structural changes may be the result of adaptive remodeling to decreased disk proteoglycan content and pressure. 70 , 71 However, disk degeneration also diverts a greater proportion of the compressive load to the end plate periphery and vertebral rim, 72 , 73 thereby reducing tensile and shear strains in the central end plate. 45 , 74
End plate disruptions upset the uniformity of disk stress distributions. 75 , 76 This, in turn, is thought to precipitate alterations in disk structure and matrix composition that typify disk degeneration because abnormal pressures can inhibit disk cell metabolism and accelerate matrix degradation. 77 , 78 , 79 , 80 End plate disruption may also impede nutrient transport to the cells within the nucleus of the disk or incite inflammatory responses in the disk or vertebra. 50 , 81 , 82 , 83 , 84
Pathologic Degeneration
A theoretical requirement for discogenic pain is pathologic innervation. 85 , 86 In the normal disk, innervation is restricted to the outer layers of the annulus. 87 By contrast, vertebrae are well innervated: the periosteum is the most densely innervated bone component, but when the total tissue volume is considered, the bone marrow receives the greatest number of sensory fibers. 36 , 88 This extensive network of nerves may modulate hemopoiesis and bone metabolism. 89 Marrow sensory and sympathetic fibers are frequently associated with blood vessels and consist of both fast myelinated fibers (group III or A-delta fibers with diameters ranging from 1 to 5 μm) that transmit sharp pain and slow unmyelinated fibers (group IV or C-fibers with diameters ranging from of 0.5 to 2 μm) that transmit dull or aching pain. Nearly all marrow pain fibers express calcitonin gene-related peptide (CGRP) and coexpress TrkA and p74 receptors that are sensitized by nerve growth factor (NGF). 90 These marrow fibers are the first to encounter and presumably become excited by pathologic processes occurring in the bone space. Consequently, patients can experience bone pain from elevated interosseous pressures even when the pathology is confined to the marrow, 23 and this pain can be ameliorated when bone innervation is ablated, such as after vertebroplasty. 91
Provocation diskography (PD) is considered by many as the gold standard for diagnosing discogenic pain. 92 The procedure consists of injecting a contrast agent into disks of a lightly sedated patient while monitoring the injected volume, pressure, contrast distribution pattern, and patient's pain response. 93 A positive test is based on pain intensity, concordance (similarity to pain before the procedure), degree of annular disruption, and presence of a negative adjacent control disk. 94 This test can reveal internal disk disruption and also identify which disks are painful and may be appropriate for treatment. Although not without controversy regarding its usefulness and safety, 95 , 96 , 97 PD results can be quite accurate (specificity of 0.94 and a false-positive rate of 6%) if performed using low-pressure technique. 93
The theoretical basis for PD-provoked pain is mechanical stimulation of chemically sensitized nociceptors. 98 Sensitized nociceptors within the outer annulus of the disk can be stretched by nucleus pressurization if the annulus is weakened by fissures. Nociceptors in the end plate may be similarly perturbed if the end plate is weakened by damage. 99 For example, the end plates can deflect comparably to the annulus during diskography (0.3 mm versus 0.5 mm, respectively, at 75 to 100 psi), 99 , 100 and end plate deflection can increase in the presence of bone microdamage. 101 In support of this concept are the observations that increased vertebral interosseous pressures: (1) occur during PD as pressures are transmitted to adjacent disks; 102 (2) can cause pain; 23 , 103 and (3) are elevated in chronic low back pain (CLBP) patients. 104 Further, end plates removed from patients with chronic back pain show proliferation of blood vessels and CGRP-positive nerve fibers in the subchondral bone that predominates in areas of end plate damage and are sensitive to direct mechanical stimulation. 37 , 105 , 106
Perhaps the best evidence for the role of end plates in CLBP is the association between PD-confirmed discogenic pain and vertebral bone marrow abnormalities. Three types of vertebral bone marrow lesions (BML) noted on magnetic resonance imaging (MRI) were first described by Modic et al in 1988. 107 Type I changes (fibrovascular replacement) show decreased signal intensity on T1-weighted images and increased signal intensity on T2-weighted images. Histopathology of type I changes shows an active inflammatory stage that coincides with disruption and fissuring of the end plate and vascularized granulation tissue within the marrow. Type II changes show increased signal intensity on T1-weighted images and an iso- or slightly hyperintense signal on T2-weighted images. Type II changes correlate with fatty marrow replacement. Both types of Modic changes are dynamic in that type I changes can convert to type II or back to normal marrow, and similarly, type II can convert back to type I. 108 , 109 , 110 Type III changes are represented by decreased signal intensity on both T1- and T2-weighted images that correlate with dense woven bone (sclerosis).
Data from multiple independent studies suggest that Modic type I and II changes adjacent to the end plate are among the most specific of all MRI observations for predicting concordant PD-induced pain (Table 1). In one prospective study, moderate to severe Modic type I or II end plate abnormalities correlated 100% of the time with positive concordant pain at the adjacent disk. 116 Recently, increased end plate innervation has also been reported for end plates with Modic changes. 117 , 118 However, the presence of Modic changes are not very sensitive (15 to 65%) to PD-confirmed disk pain. 111 , 112 , 113 , 114 , 115 Poor sensitivity may reflect the categorical and subjective techniques used to classify BML, 119 rather than ones that are quantitative and objective. It may also be that innervated end plate damage is not well visualized using standard MRI techniques, because the end plate has a short T2 that shows little signal with pulse sequences that have long echo times. Newer imaging sequences with ultrashort echo times may therefore help discriminate between patients with and without end plate pathologies (Fig. 4).
Summary of incidence and diagnostic values of Modic changes for diskography-concordant CLBP
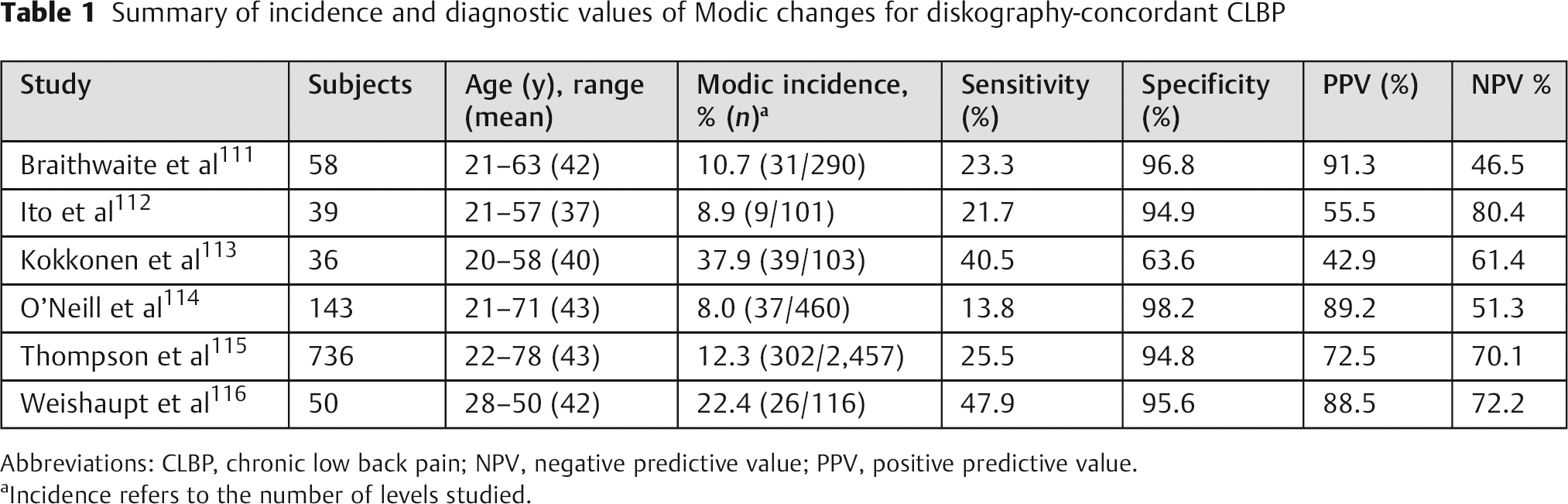
Abbreviations: CLBP, chronic low back pain; NPV, negative predictive value; PPV, positive predictive value.
Incidence refers to the number of levels studied.
The precise etiology of BML is not well understood, but it appears to involve autoimmune and inflammatory responses to chemicals produced by disk cells. Crock first proposed that vertebral BML result from inflammatory constituents that diffuse from the adjacent disks, 120 because disk tissue can trigger an autoimmune response due to secretion of proinflammatory and neurogenic factors such as interleukin-1, -6, and -8; tumor necrosis factor-α, prostaglandin E2, monocyte chemotactic protein-1, and NGF. 121 , 122 , 123 , 124 , 125 These nucleus-derived chemicals can both sensitize existing nerves and promote new nerve growth. 126 ,127,128 Additionally, end plate nerves can be irritated via accumulated by-products of disk cell anaerobic metabolism, such as lactic acid. 129
A predisposing factor for enhanced communication between the nucleus and vertebral marrow is end plate damage. For example, end plate damage has been qualitatively related to BML that contain pain fibers, 105 , 117 and more directly, end plate damage significantly increases diffusion between the vertebra and nucleus. 82 , 130 Not surprisingly, therefore, various forms of end plate defects have been clinically associated with disk degeneration and axial back pain. 131 , 132 , 133 , 134 These include Schmorl nodes, fractures, avulsions/erosions, and calcifications (Fig. 5). Small end plate defects are difficult to observe radiographically, 58 , 101 and they are thought to be a common component of normal aging. 135 Schmorl nodes are large, focal end plate indentations that represent herniations of nucleus into adjacent vertebrae and are significantly associated with disk degeneration severity. 136 Because it can be challenging to distinguish between nodes that arise prior to skeletal maturity (e.g., at sites of cartilage defects that remain after notochord regression and growth plate closure) and nodes that form traumatically secondary to age-related subchondral weakening, 137 , 138 , 139 , 140 the results from clinical studies relating Schmorl nodes to symptoms are mixed. For example, Schmorl nodes are relatively common in asymptomatic individuals. 136 , 141 However, in cases where the nodes associate with CLBP, MRI shows evidence of BML and fibrovascular bone marrow changes, 142 , 143 which suggests a traumatic etiology. A recent cadaveric study relating different types of end plate defects to back pain history demonstrated a clear dose effect: larger lesions were associated with more severe degeneration and more frequent back pain (odds ratio = 17.88). 144
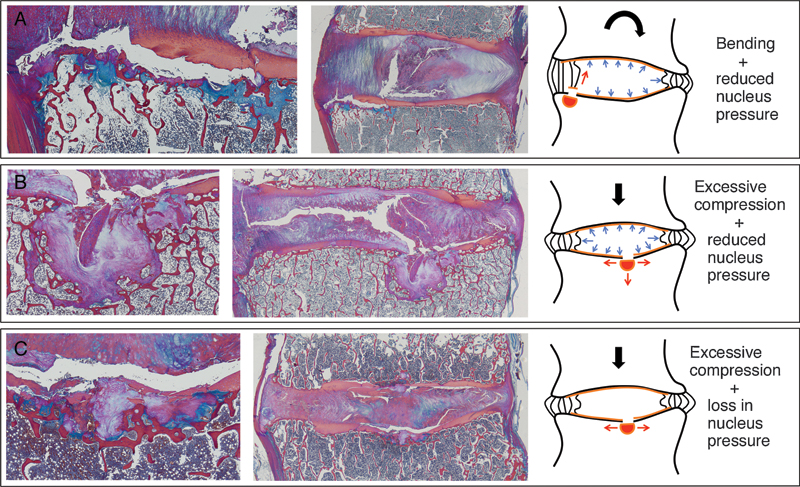
Various end plate defects with hypothesized etiologies. (A) End plate cartilage avulsion resulting from bending motion that causes traction at the interface between the end plate and inner annulus. (B) Traumatic node with end plate fragment resulting from excessive compression with a healthy, gel-like nucleus pulposus. (C) Central end plate fracture with exposed trabeculae resulting from excessive compression with a degenerate, fibrous nucleus pulposus.
The finding that certain types of end plate defects predominate at distinct levels and locations in the spine suggests that end plate defects have unique etiologies (Fig. 6). Nodelike defects are more common in the central end plates of the upper lumbar and thoracolumbar spine, 131 where trabecular bone density is lower, 145 end plates are less strong, 146 and subchondral softening is more severe. 145 , 147 In contrast, avulsions/erosions and calcifications are more common at the vertebral rim in the lower lumbar spine, 148 , 149 , 150 , 151 where the greater range of motion in flexion and extension could lead to increased traction at the junction of the annulus and end plate cartilage.152 Calcification and sclerosis at the vertebral rim of the lower lumbar levels may be a consequence of repeated compressive trauma.
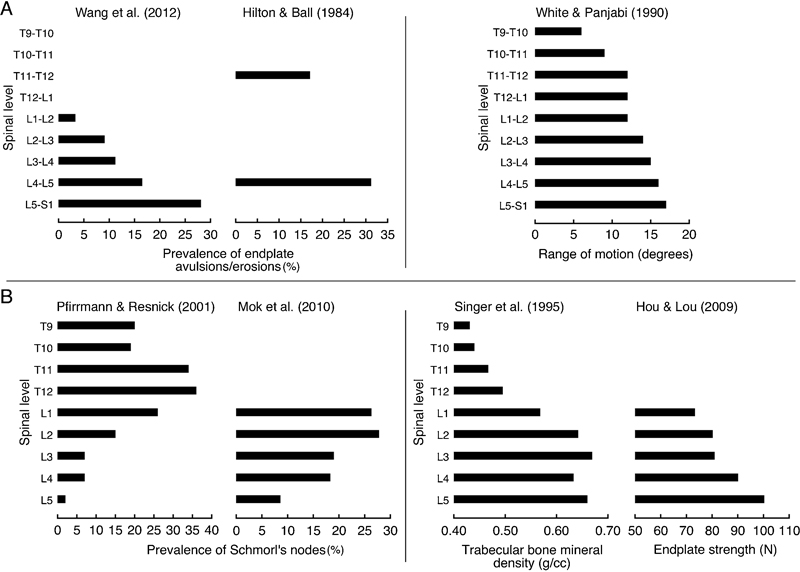
Prevalence of end plate pathologies in different regions may arise from distinct biomechanical conditions. (A) In the lower lumbar spine, the prevalence of end plate cartilage avulsions and erosions increases caudally, 148 , 149 , 150 mirroring the increase in range of motion (combined flexion/extension data are shown).152 (B) In the upper lumbar spine, Schmorl nodes increase cranially, 131 mirroring the decrease in trabecular bone mineral density 145 and end plate strength. 146
End Plates and Disk Regeneration
There is growing interest in developing novel technologies to repair or regenerate the degenerated intervertebral disk. These approaches consist of increasing the signals for cell matrix synthesis (gene or growth factor therapy) in attempts to reestablish nuclear swelling. 153 Because the disk is relatively acellular (typically 4,000 cells/mm3 in the nucleus), 12 it may also be critical to augment these approaches by introducing cells.
It is unclear whether end plate permeability and vascularity in degenerated disks is sufficient to support increases in cell density and metabolism (because poor nutrition may have led to the degeneration in the first place). These uncertainties may ultimately limit or prevent the successful extrapolation of disk repair technologies from small animals to man. Importantly, if it proves true that disk cell density and ultimately disk degeneration are tightly coupled to end plate permeability, then by definition, disk cellularity cannot be enhanced without commensurate increases in end plate permeability and vascularity (by yet undetermined methods). Similarly, efforts to increase synthesis rates of existing cells (by gene or growth factor therapy) may create excessive demands on a tenuous nutrition supply and thereby promote cell death.
Summary and Future Directions
End plates play a central role in maintaining disk and vertebral health. Their structure and composition reflect a balance between competing requirements for porosity and strength. As a result, end plates are particularly vulnerable to damage. End plate regions weakened by damage facilitate communication between the disk nucleus and vertebral marrow, which can cause an adverse combination of end plate nerve proliferation plus chemical sensitization and mechanical stimulation.
Unfortunately, current diagnostic tools do not depict subtle end plate damage that associates with neoinnervation, and consequently, the clinical significance of end plate damage may be underappreciated. Therefore, more research is needed to clarify the role of end plates in accelerated disk degeneration and discogenic pain. Three areas are of particular importance. First, more data are needed to define the dependence of disk cell function on the quality of end plate vascularity and end plate permeability. This information will help establish individual risk factors that associate with disk degeneration severity. Second, disk/vertebra structural models with improved fidelity of end plate architecture and composition are needed to define mechanisms of increased end plate damage risk. This information may guide development of new diagnostic tools that stratify injury risk. Third, the biological basis for BML and end plate neoinnervation is unknown. Studies are needed to identify the chemical factors and cellular participants in the development of innervated, fibrovascular bone marrow. Ultimately, strong links between clinical observations in back pain patients and scientific studies on disk cells and tissues are a necessity given the lack of validated animal models of discogenic pain.
Funding Sources
National Institutes of Health Grant AR052811 Relievant Medsystems.
Disclosures
J. C. Lotz, Consultant, Research Support, Stock/Options: Relievant Medsystems, Nocimed LLC
A. J. Fields, None
E. C. Liebenberg, None